黃瓜離體雌核發育早期相關基因的轉錄組初步分析
2020-01-08 01:20:08魏愛民車黎明魏盼盼韓毅科陳正武杜勝利
華北農學報 2019年6期
魏愛民,姚 堯,劉 楠,車黎明,魏盼盼,韓毅科,陳正武,杜勝利
(1.天津科潤農業科技股份有限公司黃瓜研究所 蔬菜種質創新國家重點實驗室 天津市蔬菜研究中心,天津 300192; 2.南開大學 生命科學學院,天津 300071)
單倍體技術是現代商業化育種的重要手段之一,對作物遺傳育種具有重要意義[1]。黃瓜未受精子房培養是獲得單(雙單)倍體植株的重要途徑,已在育種實踐中規模化應用,但在應用中存在基因型差異大、胚胎誘導率及植株再生率較低的問題。研究黃瓜未受精子房培養(離體雌核發育)胚胎發育的分子機制,獲得黃瓜未受精子房培養胚胎發育的關鍵基因,可以為進一步優化完善黃瓜未受精子房培養技術提供理論依據。有關黃瓜未受精子房培養的研究報道,較多圍繞基因型、激素種類及濃度、培養方法等影響因素對黃瓜離體雌核胚胎發育的影響,旨在建立相應的培養技術體系。波蘭學者Gémes-Juhász等[2]對6種不同基因型的黃瓜進行了取材時期和預處理方式的研究,發現熱處理可以提高最適發育階段雌核發育的頻率,最高單倍體植株再生頻率為7.1%。杜勝利等[3]通過對子房大小、激素種類及濃度等影響因素進行系統研究,成功研制華北刺瘤型黃瓜未受精子房培養技術,已規模化應用。王燁等[4]通過黃瓜未受精胚珠離體培養獲得單倍體植株。有關黃瓜未受精子房培養胚胎發育分子機制研究較少:杜勝利等[5]在雌核發育與非雌核發育過程中,未受精子房培養早期POD酶活性急劇上升,IAA和ABA含量明顯下降[5];Wei等[6]對不同基因型多胺含量變化進行了比較研究,發現離體雌核發育高頻材料游離態spd+spm、結合態spd+spm含量均較高,總多胺含量較高,多胺組分及含量變化是離體雌核發育啟動過程的重要特征;韓毅科等[7]應用IEF-SDS-PAGE技術對雌核發育早期蛋白表達進行研究,發現11個特異多肽點,其可能在胚胎啟動階段對細胞功能或組織的形態建成起重要作用。應用基因表達譜芯片,從轉錄組水平研究黃瓜未受精子房培養胚胎發育早期基因表達,揭示黃瓜離體雌核發育的分子機制,相關研究內容未見報道。本研究應用黃瓜未受精子房培養胚胎發育高頻基因型材料與低頻基因型材料,分別提取胚珠部位總RNA,采用基因芯片方法,從基因表達方面研究與黃瓜未受精子房培養胚胎發育早期緊密相關的基因,揭示黃瓜離體雌核胚胎發育機制,為進一步優化提高黃瓜未受精子房培養技術提供理論依據。
1 材料和方法
1.1 試驗材料
以未受精子房培養過程中胚胎發生率差異篩選極端材料:高頻基因型材料W7(未受精子房離體培養胚胎發生率為50%,華北刺瘤類型材料)和低頻基因型材料D03-3(未受精子房離體培養胚胎發生率為0,荷蘭光滑型材料)為供體材料,進行未受精子房培養。
1.2 黃瓜未受精子房的離體培養與胚珠剝離
采集開花前1~2 d的黃瓜雌花,去掉果柄和花冠,75%乙醇和次氯酸鈉溶液對子房表面進行滅菌,子房橫/縱切后,接種于誘導培養基中,置于25 ℃培養室、光照/黑暗16 h/8 h條件下培養。在培養的0,2,4,6 d,在體式顯微鏡下分別剝取不同基因型材料的胚珠,放入-80 ℃冰箱中保存備用。
1.3 RNA提取
采用TRIzol法提取總RNA。取混合材料約100 mg,在裝有液氮的研缽中并于冰上研磨,加入1 mL TRIzol繼續研磨,待TRIzol融化后振蕩并離心;取上清液加入氯仿,振蕩、靜置、離心;取上清液加入等體積的高鹽和異丙醇,混勻離心;吸出殘液,75%乙醇洗滌沉淀,再用RNase-free水溶解沉淀;加入DNaseⅠ、RRI等置于37 ℃水浴鍋中30 min;加入氯仿、水飽和酚反復抽提2次;取上清液加入醋酸鈉、無水乙醇沉淀置于-20 ℃冰箱沉淀過夜;次日,用75%乙醇洗滌沉淀,再用RNase-free水溶解沉淀備用。
1.4 基因芯片雜交及數據分析
1.4.1 基因芯片雜交 以高頻基因型W7為材料,分別剝取未受精子房培養0,2,4,6 d的胚珠,提取總RNA,每一時期隨機選擇20個子房,基因表達譜所用芯片為定制的規格為12×135 K的 ArrayRoche NimbleGen芯片,芯片探針的標準長度為60 mer,依據美國能源部在Phytozome網站上公布的基因序列設計。芯片雜交試驗及數據處理由北京博奧生物技術有限公司完成主要雜交過程如下:
①用 RNA clean-up試劑對各樣品總RNA進行過柱純化,用分光光度計定量,甲醛變性膠電泳質檢;②用SuperScriptⅡ 反轉錄酶將總RNA反轉錄為相應的單鏈cDNA,并在DNase Polymerase Ⅰ的作用下合成雙鏈DNA;③以cDNA為模板,利用T7 Enzyme Mix體外轉錄合成cRNA,然后純化定量;④取5 μg cRNA,用CbcScript酶,Random Primer 進行反轉錄,反轉錄產物用PCR NucleoSpin Extract Ⅱ Kit(MN) 純化;⑤取上述反轉錄產物,以Random Primer 為引物進行Klenow 酶標記,標記產物用PCR NucleoSpin Extract Ⅱ Kit(MN) 純化;⑥將標記好的樣品與雜交Buffer(3×SSC,0.2%SDS,50×Denhart′s,25%甲酰胺)混勻后,注到已經與相應mixer黏合的芯片上,采用Roche NimbleGen Hybridization System 12 雜交儀42 ℃雜交14~16 h。雜交結束后,去除mixer,分別用不同洗液清洗芯片,甩干,掃描;⑦使用Roche-NimbleGen MS200掃描儀掃描芯片;采用NimbleScan進行圖像分析,將圖像信號轉換為數字信號。
1.4.2 數據分析 差異基因表達分析采用SAM軟件,對相鄰離體培養時期差異基因數量進行兩兩對比分析,① 離體培養2 d(試驗組UP2)和離體培養 0 d(對照組UP0),以下表示方法相同;② UP4和 UP2;③ UP6和 UP4。
差異基因篩選采用倍數變化比較,倍數計算以離體培養0 d為起點,將3個不同培養時期基因表達量作兩兩對比(即UP2和 UP0, UP4和 UP2, UP6和 UP4)分析,分別得出倍數(Ratio)變化,數據經log2處理,分析3組之間的倍數變化及變化模式。
差異基因的富集分析應用在線數據庫進行,差異基因功能注釋參照擬南芥蛋白質數據庫、GenBank中非冗余蛋白數據庫以及Phytozome網站。
1.5 實時熒光定量分析
分別提取高頻基因型和低頻基因型材料4個離體培養時期未受精子房胚珠總RNA,在MLV-RTase和引物Oligo(dT)的作用下合成單鏈cDNA,反轉錄程序采用二步法,反應程序為:70 ℃ 5 min,25 ℃ 5 min,42 ℃ 60 min,80 ℃ 20 min,所用PCR儀為Bio-Rad S1000。qPCR采用20 μL體系,反應程序:95 ℃ 3 min;95 ℃ 10 s,58~60 ℃ 30 s,40個循環;溶解曲線溫度為:55~95 ℃ 10 s,81個循環。實時熒光定量試驗選擇TUA作為內參基因。qPCR引物設計使用軟件Primer primer 5.0,引物合成由上海生工生物工程有限公司完成。qPCR儀為Bio-Rad CFX connect。每個反應體系進行3次生物學重復和3次技術重復。qPCR數據采用2-ΔΔCt法進行分析,對得到的2-ΔΔCt數據進行log2處理。
2 結果與分析
2.1 黃瓜離體雌核發育轉錄組差異表達基因分析
應用基因芯片對26 147個基因的表達量進行測定,應用SAM軟件對相鄰離體培養時期基因表達量進行差異分析,基因表達量的對比分析采用倍數(Ratio)關系表示,上調基因為Ratio≥2,表達量差異大于2倍,試驗組的表達量高于對照組;下調基因為Ratio≤0.5,即表達量差異大于2倍,試驗組的表達量小于對照組;無差異表達基因為0.5≤Ratio≤2。共有8 858個基因發生了差異變化,其中上調表達基因4 366個,下調表達基因4 492個。
UP2 vs UP0 差異基因數目為5 760個,無差異基因數目為20 387個;UP4 vs UP2 差異基因數目為2 405個,無差異基因數目為23 472個;UP6 vs UP4 差異基因數目為693個,無差異基因數目為25 454個。對每一組差異表達的基因數量根據上調表達與下調表達進行分類統計,如圖1所示。
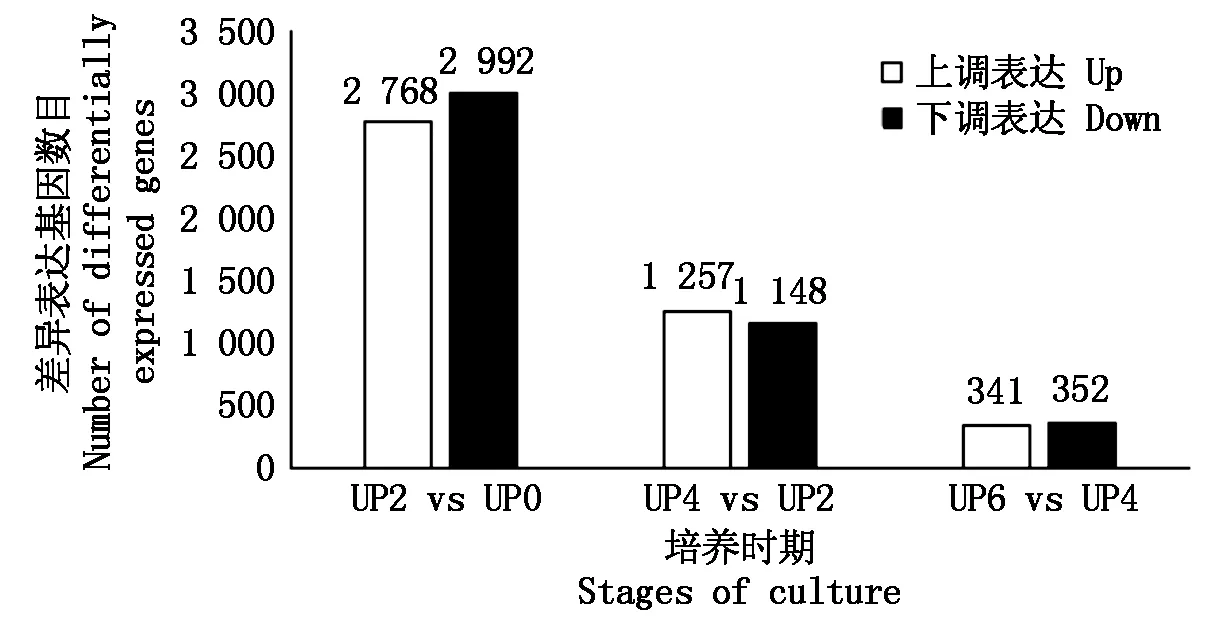
圖1 不同離體培養時期兩兩對比 的差異表達基因數目比較Fig.1 Number comparison of differentially expressed genes among different stages of culture
對3組差異分析數據中所有的上調表達基因和下調表達基因進行維恩圖解,由圖2可以看出,在所有上調基因中,有2 644,1 135,205個基因分別在某一個離體培養時期特異表達;有51,65,63個基因分別在某2個離體培養時期特異表達;有8個基因在連續3個離體培養時期特異表達。在所有的下調基因中,有2 805,1 062,171個基因分別在某一個離體培養時期特異表達;有45,39,140個分別在某2個離體培養時期特異表達;有2個基因在連續3個離體培養時期特異表達。
8個連續上調的基因,其功能主要包括白細胞介素-1受體激酶、大亞基核糖體蛋白、木葡聚糖轉移酶、水通道蛋白、細胞色素P450、三磷酸酶、過氧化物酶等,涉及的主要代謝途徑有嘌呤代謝,苯丙氨酸代謝,物質跨膜運輸、細胞凋亡、Toll-like信號代謝通路、苯并惡嗪類物質合成等。
2.2 差異表達基因的GO功能注釋及分類
2.2.1 所有差異表達基因的GO功能注釋及分類 對所有的差異表達基因做了GO功能注釋和分類統計,生物過程一類共有2 263個差異表達基因被注釋上了GO功能,進一步對所有被注釋上GO生物過程功能的差異基因進行了分類(圖3)。涉及物質運輸、磷酸化與脫磷酸化代謝過程差異表達基因最多;其次為響應刺激類的差異表達基因,占13%,主要包括響應信號、物質刺激的差異表達基因,約占6%,響應脅迫刺激的差異表達基因,約占4%,響應激素刺激的差異表達基因,約占3%。這幾類刺激的響應有可能是關系到黃瓜離體雌核發育過程中的重要途徑,是影響離體雌核胚胎發育的關鍵因素。
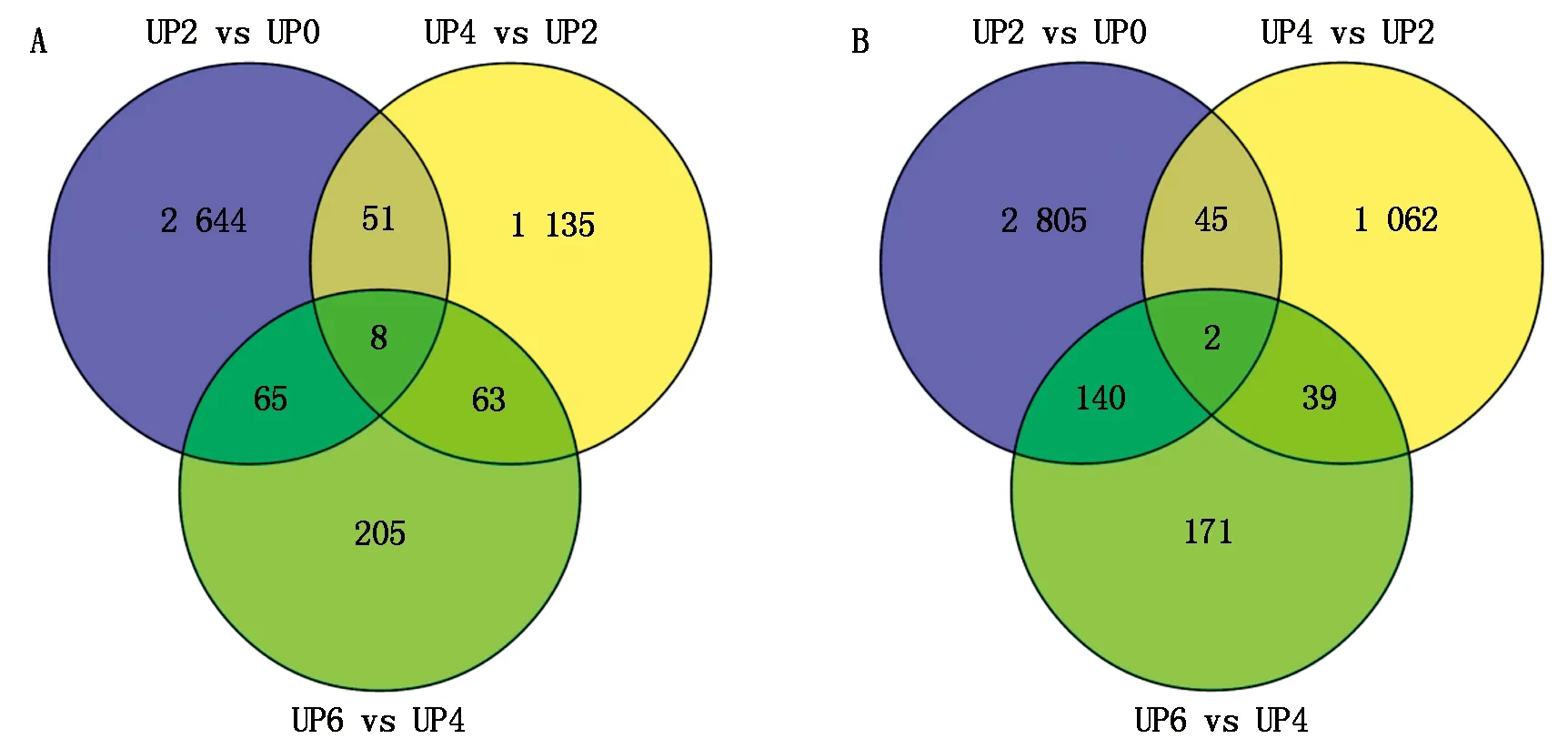
A表示3組中所有上調基因在不同離體培養時期的數目;B表示所有下調基因在不同離體培養時期的數目。 A indicates the number of up regulated genes among groups of different stages of culture; B indicates the number of down regulated genes among groups of different stages of culture.
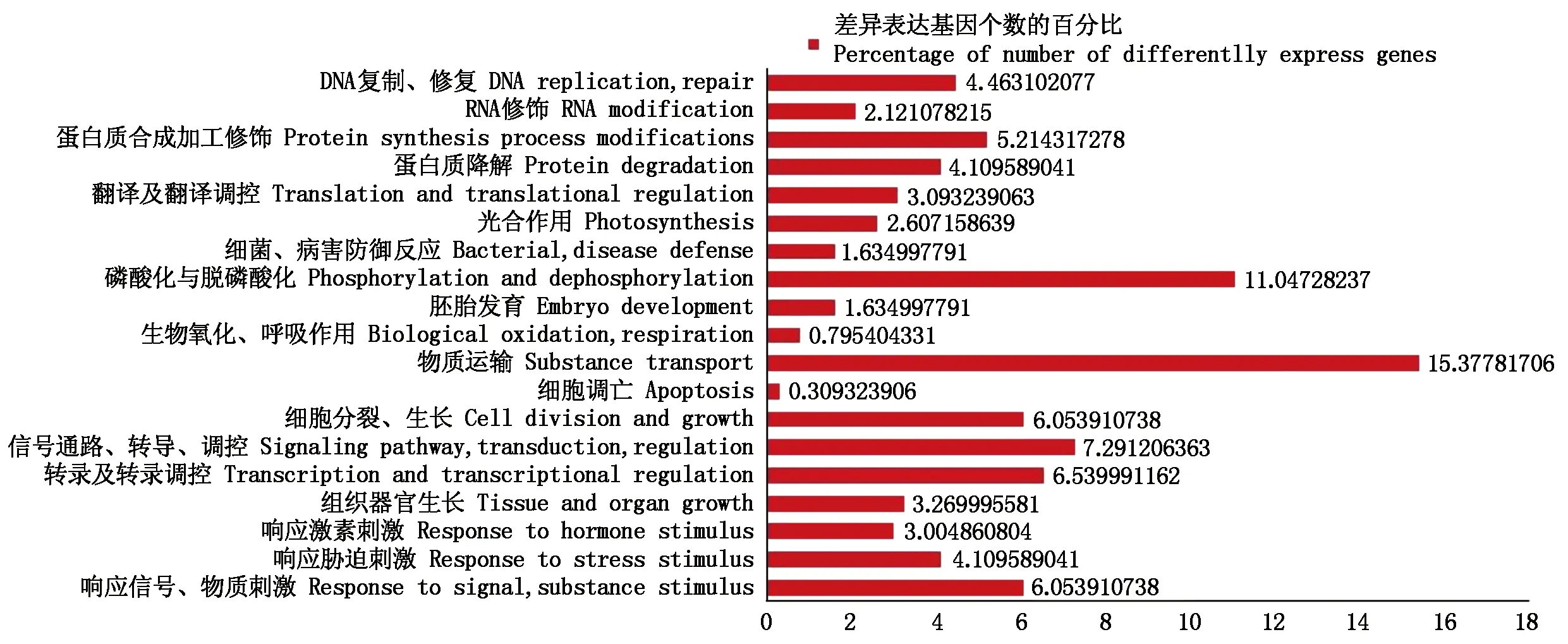
圖3 差異表達基因生物過程GO功能分類Fig.3 GO functional analysis of differentially expressed genes
2.2.2 激素相關的差異表達基因的GO功能注釋及分類 進一步對與激素相關的116個差異表達基因進行分類分析(圖4),發現響應脫落酸刺激的基因最多,占24.1%,其次是參與生長素信號轉導途徑的差異表達基因占20.7%,而響應生長素刺激、水楊酸刺激以及其他激素刺激的差異表達基因也比較多。表明這些差異表達基因在黃瓜離體雌核發育的不同時期發揮了關鍵作用。
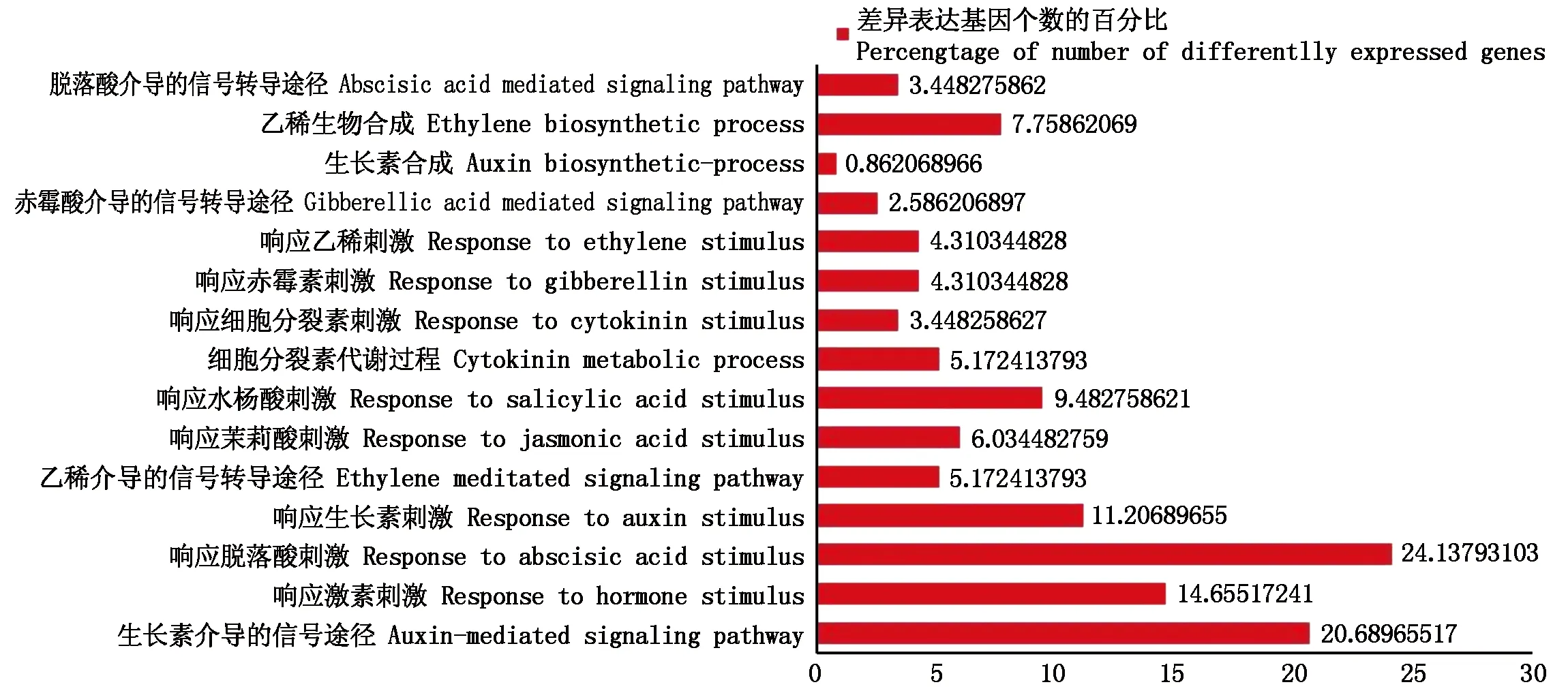
圖4 激素相關的差異表達基因的GO功能分類Fig.4 GO functional analysis of hormone related differentially expressed genes
2.2.3 激素代謝途徑相關的差異基因代謝通路富集分析 針對激素相關的差異表達基因進行了代謝通路富集分析(圖5),找出了2條與激素相關的代謝路徑,即植物激素信號轉導途徑和玉米素合成途徑。其中植物激素信號轉導途徑包括生長素信號轉導途徑及其合成、細胞分裂素信號轉導途徑、赤霉素信號轉導途徑、脫落酸信號轉導途徑、乙烯合成、水楊酸及茉莉酸代謝和植物激素油菜類內酯的代謝等途徑,而玉米素合成途徑涉及萜類化合物的生物合成,參與其代謝途徑的差異表達基因大多涉及相關糖代謝中反應酶,如脫氫酶、轉移酶等。
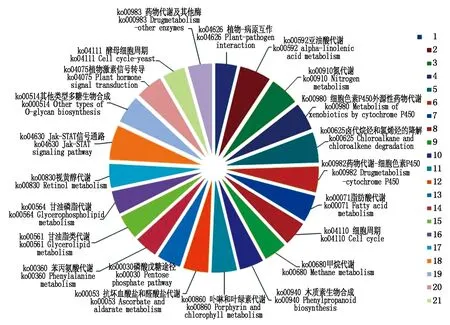
圖5 激素代謝途徑差異表達基因KEGG Pathway分析圖Fig.5 KEGG Pathway analysis of differentially expressed hormone metabolism genes
從3個方面對黃瓜離體雌核發育早期差異表達基因進行了分析和篩選:①所有上調基因的維恩圖解中,篩選出連續在3個不同離體培養時期都上調表達的8個差異基因;②從響應刺激類相關的差異表達基因中,分析并篩選了與響應激素刺激相關的差異表達基因116個,與響應脅迫刺激相關的差異表達基因194個,與響應信號及物質刺激的差異表達基因88個;③還有針對性的研究了GO生物過程中與胚胎發育有特異性關系的50個差異表達基因。共篩選出456個差異表達基因。
2.3 倍數變化模式角度進一步分析差異表達基因
黃瓜離體雌核發育在培養的前6 d已經啟動,在此階段重點關注兩類基因:基因表達倍數變化總體成上調趨勢;無顯著差異表達基因中有重要功能的基因。UP6 和 UP0這2組對比后的倍數經log2處理后,篩選出其倍數(Ratio)大于0的所有差異表達基因,此次篩選從456個差異表達基因中共得到192個差異表達基因,且在第6天時基因的表達與0 d相比呈上調趨勢。對這192個差異表達基因的倍數變化模式進行分類統計,共得出8種變化模式,如圖6差異表達基因的倍數變化可劃分為以下3種:
存在明顯拐點:模式圖type A、type B、 type E、 type F,其中type A、type E差異表達基因倍數變化為先上升再下降,type B、type F差異表達基因倍數變化呈現先下降再上升。無明顯拐點:模式圖type C、type D、 type G,其中type C差異表達基因倍數變化呈上升趨勢,type D、type G差異表達基因倍數變化呈下降趨勢。倍數變化不大:模式圖type H,即在不同培養時期基因表達量變化無顯著差異。
與離體培養0 d的基因表達相比,模式圖type A、type B、type C在某單個離體培養時期上調表達的基因;模式圖type D、type E、type F在某2個培養時期上調表達的基因;模式圖type G在連續3個培養時期都上調表達的基因;模式圖type H無顯著差異表達的基因。
根據基因表達倍數變化分類圖,以及重點挑選上調表達顯著的基因以及無顯著差異表達基因中有重要功能的基因,做進一步表達分析,最終確定了47個基因,見表1。

圖6 差異表達基因的倍數變化模式圖Fig.6 Variation pattern of magnification ratio of differential expressed genes
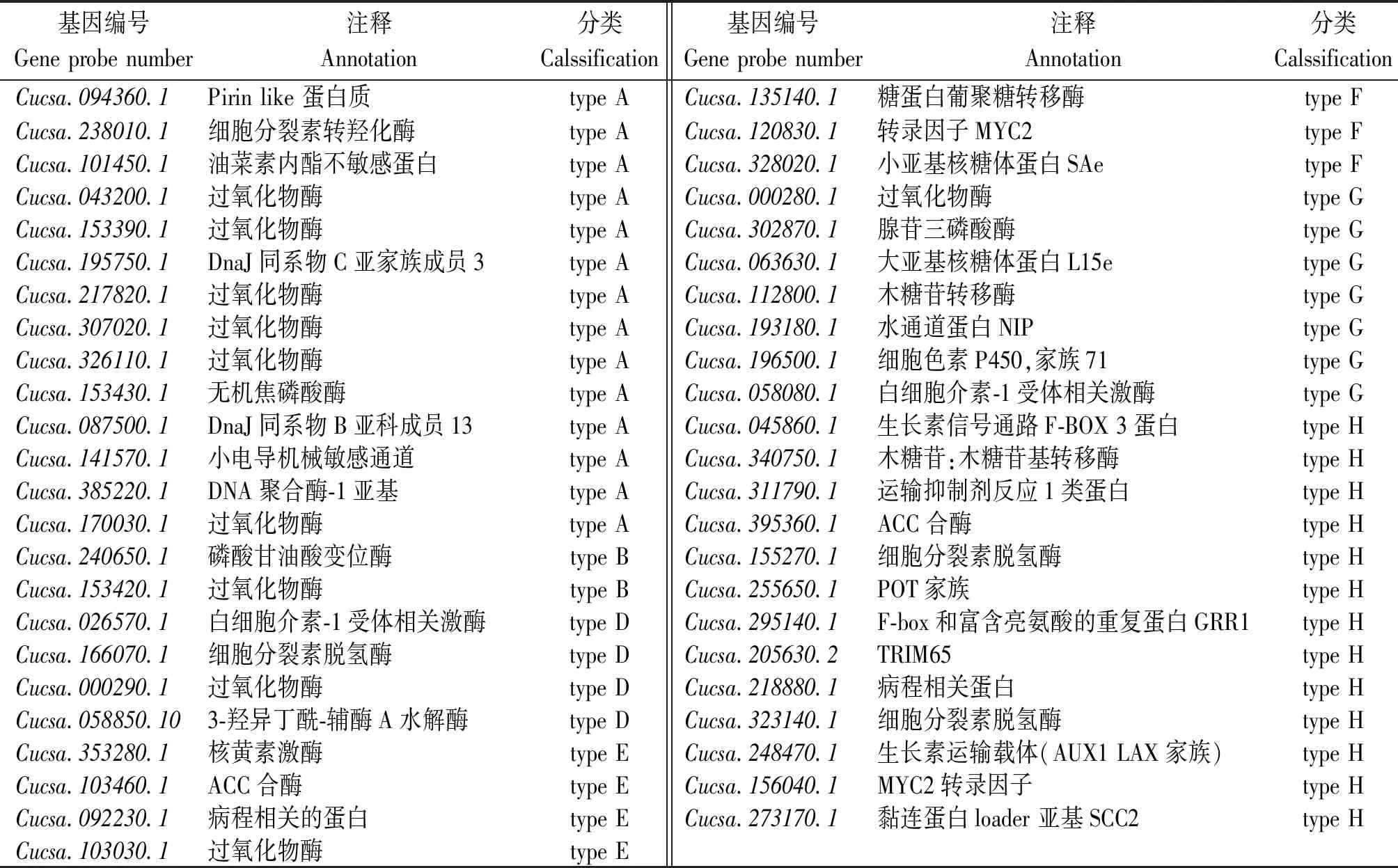
表1 篩選出的各類差異表達基因Tab.1 Selected deferentially expressed genes of different types
表中所列基因主要為:激素相關基因,主要包括玉米素合成途徑中的細胞分裂素脫氫酶、細胞分裂素羥化酶相關基因,與脫落酸信號轉導途徑相關的脫落酸受體蛋白相關基因,乙烯合成途徑中的關鍵酶和限速酶-ACC合酶相關基因以及與生長素信號轉導途徑相關的生長素信號蛋白及生長素運輸蛋白有關的基因等;編碼轉錄因子,主要包括2個MYC2家族成員;其他酶類,包括與生長素信號轉導途徑相關的絲氨酸蘇氨酸激酶,與氧化脅迫相關的過氧化物酶以及相關代謝中的轉移酶、聚合酶類;離子通道蛋白、水通道蛋白等。
2.4 實時熒光定量分析
2.4.1 總RNA提取 從上述47個基因中挑選了與激素、響應脅迫、響應信號刺激、胚胎發育等相關的22個基因進行實時熒光定量驗證。以離體雌核發育高頻、低頻基因型材料為供體材料,分別在培養0,2,4,6 d時剝取胚珠進行總RNA提取。RNA提取的電泳結果如圖7,從電泳圖中可以看出,8個樣品的RNA中28S的亮度約為18S的2倍,且條帶清晰無明顯拖尾和彌散,其質量和完整性較好,可以進行后續試驗。
2.4.2 數據驗證 本次試驗總共對22個相關基因進行了實時熒光定量驗證,從中篩選出不同離體培養時期高頻型和低頻型在倍率變化上有較大差異的8個基因。如圖8所示,8個基因在離體培養2,4,6 d與0 d對比后的芯片數據與qPCR數據的對比結果。
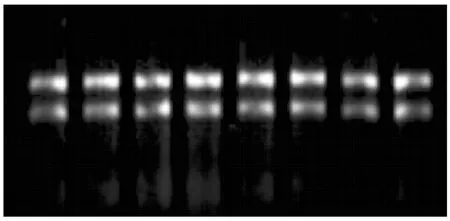
從左至右分別為高頻型0,2,4,6 d,低頻型0,2,4,6 d。 From left to right indicates RNA from ovules of 0, 2,4 and 6 d of culture both high and low efficiency genotypes respectively.
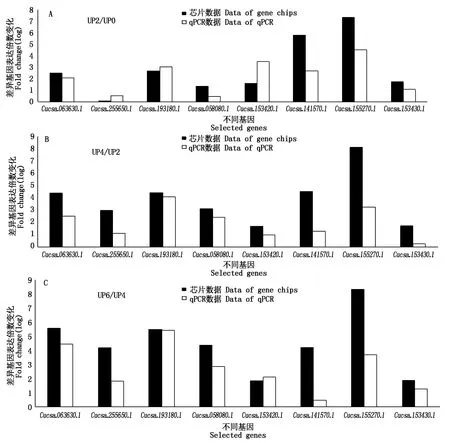
圖8 芯片數據與qPCR數據對比驗證結果Fig.8 Contrast test on the qPCR results with chip data
所選出的8個基因整體都呈上調表達趨勢,與相應天數的芯片數據對比顯示,qPCR數據與芯片數據的基因倍數變化趨勢基本一致,說明芯片數據結果真實有效。
2.4.3 差異表達基因的實時熒光定量分析 對篩選出的8個關鍵基因,分別以高、低頻基因型為材料,進行未受精子房胚珠部位不同培養時期的差異表達分析。采用離體培養2,4,6 d與0 d對比后的qPCR倍數變化進行數據分析,數據經log2處理,見圖9。
從圖中可以看出:與激素相關的3個基因Cucsa.255650.1、Cucsa.063630.1、Cucsa.155270.1在高頻型和低頻型之間有較大差異。基因Cucsa.255650.1與響應茉莉酸、水楊酸和脫落酸刺激有關,其在高頻型胚珠中的表達量總體上升,而在低頻型中除在4 d時有所上升外,在2,6 d均為下降,并且總體呈下降趨勢。Cucsa.063630.1表達產物是一種與植物生長素誘導相關的蛋白,其在高頻型中也是呈總體上升的趨勢,與低頻型的主要差異是在第6天。Cucsa.155270.1與玉米素合成途徑相關,其主要表達產物是細胞分裂素脫氫酶,其在高頻型中一直處于高量表達,而在低頻型中呈逐漸下降的趨勢。與POD酶相關的基因有2個,分別是Cucsa.153420.1和Cucsa.153430.1。結果顯示,這2種基因在高頻型離體培養的前6 d表達量都比較高,尤其在前2 d的表達量急劇上升,而在低頻型中呈下調表達或表達量低。另外KEGG通路分析顯示,這2種酶還參與了苯丙氨酸代謝途徑、甲烷代謝途徑、苯丙素合成途徑,GO分子功能還表明其參與亞鐵血紅素結合以及金屬離子的結合。另外3個相關基因:Cucsa.193180.1基因在高頻型中前6 d連續上調表達,而在低頻型中第6天下調表達,此基因為水通道蛋白NIP家族的NIP Ⅱ亞家族基因NIP 5-1,GO生物過程功能顯示其主要響應含硼酸鹽、亞砷酸鹽等含硼、砷物質。Cucsa.058080.1基因在高頻型前6 d中連續上調表達,而在低頻型中第6天呈現下調表達,此基因與蘇氨酸-絲氨酸蛋白激酶(STK)相關。Cucsa.141570.1基因在高頻型的前6 d上調表達,而低頻型中除第4天上調表達外,第2天和第6天均為下調表達,總體上為下調表達,此基因主要參與機械敏感性離子通道過程,與感受和傳遞細胞內外信號有關。

縱坐標大于0代表基因上調表達,縱坐標小于0代表基因下調表達。 Longitudinal axis value greater than 0 indicates gene up regulation whereas longitudinal axis value less than 0 indicates gene down regulation.
3 結論與討論
本研究以黃瓜未受精子房培養高頻基因型與低頻基因型胚珠部位為材料,進行離體雌核發育早期基因的表達差異研究,獲得早期差異表達基因8 858個,其中上調表達基因4 366個,被注釋上GO生物過程功能的差異基因2 263個,主要涉及物質運輸、磷酸化與脫磷酸化等代謝過程,說明離體雌核胚胎發育過程與體細胞胚胎發育過程類似,需要糖、蛋白、核酸等物質參與,胚胎建成過程中與細胞功能、組織形態建成及生理生化密切相關[8]。
體細胞胚胎發生被認為是研究植物早期發育調控和形態發生的模式系統,已通過胡蘿卜、紫花苜蓿、菊苣、松柏等體外培養體系對體細胞胚發生途徑的分子發生機制進行了深入研究,鑒定了在體細胞胚發生不同階段表達的基因,主要包括響應脅迫應答基因(熱激蛋白基因[9-10]、抗氧化酶類[11-12])、響應激素刺激基因(受生長素調控的基因[13]、ABA誘導系統相關基因[14-17])、信號轉導相關基因(鈣調素蛋白[18]、蛋白激酶[19-20]、應答激酶[21-23])等。植物游離小孢子培養在農作物單倍體育種中已成為研究的熱點之一,研究者從細胞形態學、代謝組分及分子水平對小孢子胚胎發生的機制進行了研究,分離鑒定了多個與胚胎發育相關的基因,主要包括在胚性小孢子時期表達的基因,EcMt[24]、NtEPc[25]、熱激蛋白基因[26]等;多細胞結構過程中表達的基因,BBM[27]、AGL15[28]、SERK[29]等。
本研究重點針對響應激素刺激、脅迫應答、信號轉導等方面的差異表達基因進行篩選。篩選出的重要基因包含:生長素和細胞分裂素誘導合成相關的2個基因Cucsa.063630.1和Cucsa.155270.1,在離體培養過程中在高頻型胚珠中表達量高,呈總體上升的趨勢,而在低頻型胚珠中表達量低,基本呈下降趨勢,推測其可能為黃瓜離體雌核早期胚胎發育中與激素代謝相關的重要基因。
參與過氧化物酶表達的2個基因,其在前6 d處于上調表達,而且在離體培養的第2天表達量急劇升高,代謝通路分析顯示這2個基因參與苯丙氨酸代謝途徑、甲烷途徑以及苯丙素合成途徑。這幾類代謝過程是植物代謝中的次生代謝,其產物是細胞生命活動或植物生長發育正常運行的非必需的小分子化合物,苯丙氨酸代謝途徑中的限速酶苯丙氨酸轉氨酶(PAL)研究較早、較多,在植物響應機械損傷過程中誘導表達,切口部位PAL活性增加、切口褐化,激素刺激也可誘導其表達[26]。本研究中參與苯丙氨酸代謝途徑的基因在胚胎發育早期高表達,推測其與機械損傷及激素誘導有關。
本研究發現一個與水通道蛋白相關的基因,屬于NIP家族成員。研究表明,NIPⅡ家族中的AtNIP 5;1和AtNIP6;1具有運輸硼的功能[30-31]。推測本試驗發現的NIP基因可能與含硼物質的運輸有關。在植物代謝方面,硼作為生長素調節的輔助因子,對生長素代謝途徑有一定影響[32]。此基因在黃瓜離體雌核發育的早期過程中表達活躍,可能與胚胎發育過程中含硼物質參與細胞組分構建、相關激素代謝以及抵抗脅迫環境等因素有關。
本試驗還發現一個植物體細胞胚胎發育受體蛋白激酶類基因,為受體蘇氨酸-絲氨酸蛋白酶(STK)類基因。此類蛋白激酶的一個關鍵作用就是參與抵抗外界環境脅迫,如低溫、干燥、鹽脅迫等[33]。除此之外,在擬南芥的研究中已經發現并證實了20多個與STK相關的基因,不僅參與脅迫環境應答,還響應脫落酸、水楊酸等激素刺激相關的代謝[34]。此基因在高頻型前6 d中一直呈現上調表達,而低頻型在第6天則呈現下調表達,因此,推測此基因在胚胎發育早期過程中,參與STK對外界脅迫的應答方面有一定聯系。
本研究篩選到的重要基因還包含與蛋白質的合成、磷酸化等相關基因:Cucsa.058850.10為線粒體3-羥基異丁酰基-CoA水解酶、Cucsa.063630.1為大亞基核糖體蛋白L15e、Cucsa.302870.1為三磷酸酶基因;與淀粉合成相關基因:Cucsa.112800.1為木葡萄糖基轉移酶、與細胞分裂相關基因Cucsa.273170.1為黏連蛋白SCC2、與病原體相關蛋白Cucsa.218880.1。在對不同作物游離小孢子培養胚胎發生的分子機理研究中,均有同類基因被發現,這些基因與小孢子胚胎發生中蛋白質降解、淀粉合成、啟動孢子體途徑的細胞分裂等過程相關。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國衛生(2014年12期)2014-11-12 13:12:52
