甘藍(lán)型油菜開花時間的遺傳分析及相關(guān)分子標(biāo)記
2020-01-07 09:58:56張冰冰秦夢凡梁峰豪左凱峰徐愛遐
華北農(nóng)學(xué)報 2019年6期
關(guān)鍵詞:模型
張冰冰,劉 霞,秦夢凡,梁峰豪,張 燕,左凱峰, 郭 娜,馬 寧,黃 鎮(zhèn),徐愛遐
(1.西北農(nóng)林科技大學(xué) 農(nóng)學(xué)院,陜西 楊凌 712100;2.吳忠市鹽池縣市場監(jiān)督管理局,寧夏 吳忠 751100)
甘藍(lán)型油菜開花時間與其成熟早晚密切相關(guān),同時也是影響油菜產(chǎn)量與品質(zhì)的重要因素。開花時間既是數(shù)量性狀,又是復(fù)合性狀,受遺傳、栽培條件與環(huán)境的共同影響,導(dǎo)致開花時間的遺傳研究相對滯后[1]。深入研究甘藍(lán)型油菜開花時間遺傳規(guī)律,對選育甘藍(lán)型油菜早熟品種具有重要理論和實踐意義。目前,有關(guān)油菜開花期遺傳模型的研究,一直受到國內(nèi)外學(xué)者的普遍關(guān)注。郭志剛[2]發(fā)現(xiàn),油菜開花主要受多基因調(diào)控,但存在主基因加顯性效應(yīng),蔡長春[3]認(rèn)為,開花期主要由2對或2對以上主基因及多基因控制,羅玉秀等[4]認(rèn)為,開花期受2對加性基因控制。不同研究者的研究結(jié)果不盡一致,試驗材料不同可能是導(dǎo)致結(jié)果不一致的主要原因。甘藍(lán)型油菜72-27-1-2(P2)具有早花、早熟等優(yōu)良特性,為明確其早花性狀的遺傳,以晚開花甘藍(lán)型油菜YG-1(P1)與其雜交構(gòu)建6世代遺傳群體(P1、P2、F1、B1、B2和F2),應(yīng)用數(shù)量性狀主基因+多基因混合遺傳模型聯(lián)合分析法對開花時間進(jìn)行遺傳分析,研究結(jié)果可進(jìn)一步明確甘藍(lán)型油菜開花時間的遺傳規(guī)律,指導(dǎo)早熟品種選育。
分子標(biāo)記輔助育種是利用分子標(biāo)記與決定目標(biāo)性狀基因緊密連鎖的特點,通過檢測分子標(biāo)記,即可檢測到目的基因的存在,具有快速、準(zhǔn)確、不受環(huán)境條件干擾的優(yōu)點。SSR分子標(biāo)記技術(shù)由于其操作簡單方便、要求不高等優(yōu)點,是目前最常用的微衛(wèi)星標(biāo)記之一。近年來,已有學(xué)者在甘藍(lán)型油菜中利用分子標(biāo)記技術(shù)進(jìn)行了大量關(guān)于開花相關(guān)基因QTL定位的研究。石鵬[5]檢測到28個開花期 QTLs,解釋的表型貢獻(xiàn)率介于0.55%~16.33%。Raman等[6]檢測到20個控制開花時間的 QTL位點,可解釋表型變異的2.4%~28.6%。傅鷹[7]檢測到19個開花期QTLs,揭示了2.35%~47.68%的表型變異。柳海東[8]檢測到 48個QTLs,解釋表型變異的3.2%~46.5%。本研究利用SSR分子標(biāo)記技術(shù),開發(fā)與開花時間相關(guān)的分子標(biāo)記,對油菜開花時間基因的定位及標(biāo)記輔助育種具有重要意義。
1 材料和方法
1.1 試驗材料及性狀考查
以晚開花甘藍(lán)型油菜YG-1(P1)與早開花甘藍(lán)型油菜72-27-1-2(P2)及其配制的F1、B1、B2、F2為遺傳分析材料,其中YG-1平均開花時間(播種到開花的天數(shù),同d表示)197 d左右,72-27-1-2平均開花時間180 d左右。以F2群體(127個單株)為分子標(biāo)記篩選群體,上述材料均由西北農(nóng)林科技大學(xué)農(nóng)學(xué)院油菜遺傳及分子育種實驗室提供。分別于2017年9月18日和9月20日種植于西北農(nóng)林科技大學(xué)三原斗口試驗站和楊凌曹新莊試驗田,常規(guī)栽培管理。2018年4月避開行兩端的邊株逐株掛牌并記錄6世代各單株開花日期。油菜開花標(biāo)準(zhǔn)參考伍曉明等[9]以油菜主莖第一朵花開為標(biāo)準(zhǔn)。
1.2 性狀遺傳分析
利用Microsoft Excel 2007繪制開花時間次數(shù)分布圖,SPSS 19.0進(jìn)行基本統(tǒng)計量、正態(tài)性檢驗及親本顯著性分析。利用SEA 1.0軟件進(jìn)行開花時間遺傳分析(軟件由華中農(nóng)業(yè)大學(xué)植物科技學(xué)院章元明教授提供網(wǎng)址:https://cran.r-project.org/web/packages/SEA/index.html)。依據(jù)AIC值最小和適合性檢驗(均勻性、Smirnov和Kolmogorov檢驗)確定最優(yōu)遺傳模型;最小二乘法進(jìn)行最優(yōu)遺傳模型遺傳參數(shù)估算。
1.3 DNA提取及SSR分子標(biāo)記
在幼苗四-五葉期,取雙親及F2群體單株幼嫩葉片0.5 g,采用改良的CTAB法[10]提取DNA,利用Lab Tech紫外可見分光光度計測定DNA濃度,最終將樣品DNA濃度稀釋至50 ng/μL使用。
SSR分子標(biāo)記引物來源于http://ukcrop.net/perl/ace/search/BrassicaDB與http://www.brassica.info/ssr/SSRinfo.htm,均由上海生工試劑公司合成。SSR反應(yīng)體系:總體積10 μL,包括:DNA模板3.0 μL、引物1.5 μL、10×Buffer 1.0 μL、25 mmol/L MgCl20.8 μL、10 mmol/L dNTPs 0.2 μL、Taq酶0.1 μL、ddH2O 3.4 μL。PCR反應(yīng)程序設(shè)定為:94 ℃預(yù)變性5 min;94 ℃ 變性30 s,Tm退火30 s,72 ℃延伸75 s,35個循環(huán);72 ℃再延伸5 min,4 ℃保存。擴(kuò)增產(chǎn)物利用6%聚丙烯酰胺凝膠電泳,銀染程序參考陸光遠(yuǎn)等[11]的方法。
帶型記錄依據(jù)群體在親本間具有的差異性條帶,清晰的帶記為“1”,未出現(xiàn)條帶記為“0”,符號“-”表示整體帶型缺失或模糊難以判讀。選取群體極端早開花20株與極端晚開花20株,參照張發(fā)[12]的方法,利用SPASS 19.0對開花時間與分子標(biāo)記進(jìn)行相關(guān)性分析。
2 結(jié)果與分析
2.1 各世代開花時間數(shù)據(jù)分析
楊凌和三原兩地各世代開花時間見表1。結(jié)果表明,親本間開花時間具有明顯差異,YG-1(P1)的開花時間明顯晚于72-27-1-29(P2),同一環(huán)境下雙親間開花時間存在極顯著差異,不同環(huán)境下同一親本間無顯著性差異;兩地F1開花時間平均值均介于雙親之間,且略偏向于早花親本P2,兩地B1、B2和F2世代開花時間的平均值也介于雙親之間;B2群體均有超早親分離,楊凌B1、F2群體發(fā)現(xiàn)超晚親分離;由表2知:兩環(huán)境中楊凌點B2、F2群體及三原B1群體的K-S正態(tài)檢驗P值大于0.05,符合正態(tài)分布,而其他分離世代P值均小于0.05,但偏度或峰度基本都介于-1與1,呈偏離正態(tài)分布,各分離世代中開花時間性狀均表現(xiàn)單峰分布(圖1,2),表明甘藍(lán)型油菜開花時間遺傳可能存在主基因作用并有多基因修飾效應(yīng)。
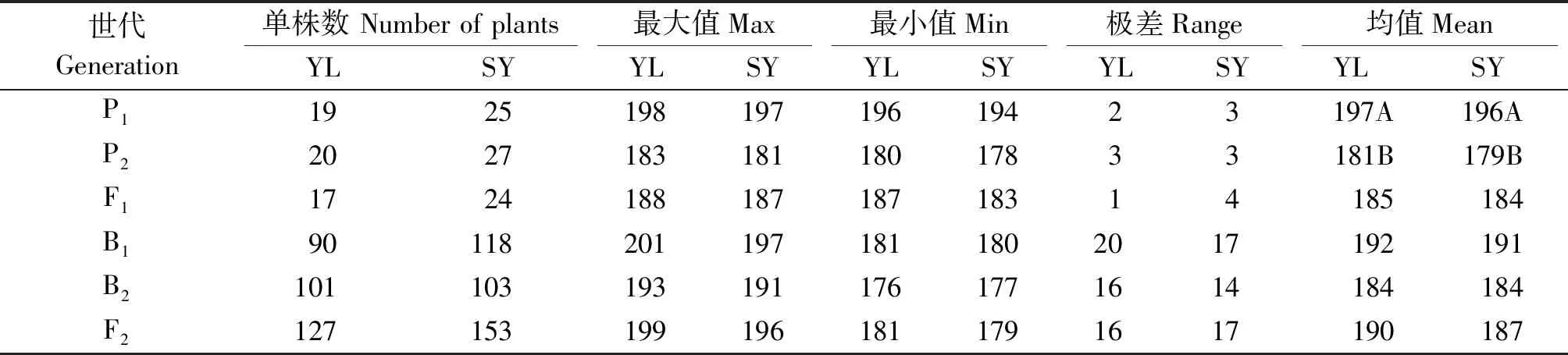
表1 各世代開花時間統(tǒng)計及親本差異顯著性檢驗Tab.1 Performance of flowering time in six generations and significance test of difference between the parents
注:YL.楊凌;SY.三原;表2-4、圖1-2同。標(biāo)以不同字母的數(shù)值在0.01水平下差異顯著,相同字母的數(shù)值在0.01水平下無顯著性差異。
Note:YL.Yangling;SY.Sanyuan;The same as Tab.2-4,Fig.1-2.Values followed by different letters are significantly different at the 0.01 probability level, values followed by same letters are not significantly different at the 0.01 probability level.
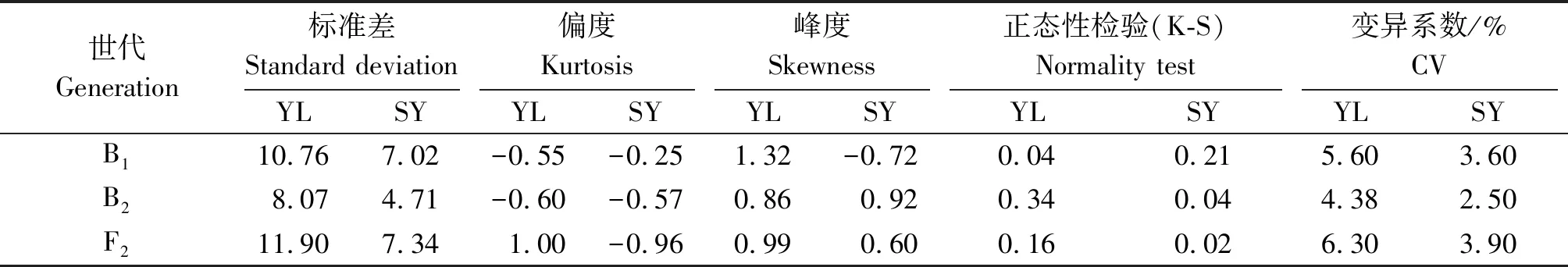
表2 B1、B2和F2開花時間的統(tǒng)計分布和正態(tài)性檢驗Tab.2 Statistic analysis and normal distribution test for flowering time in B1,B2 and F2 generations
2.2 開花時間主基因+多基因遺傳分析
2.2.1 遺傳模型分析 利用主基因+多基因混合遺傳模型的多世代聯(lián)合分析方法,以SEA 1.0軟件對兩地6世代開花時間進(jìn)行遺傳分析,獲得5類24種模型。依據(jù)AIC數(shù)值最小選取準(zhǔn)則,選取最小AIC值及與之比較接近的一組共2個模型作為開花時間性狀的備選模型(表3)。通過適合性檢驗(0.05水平)進(jìn)一步確定比較適合的遺傳模型,結(jié)果表明:共30個統(tǒng)計量中,楊凌點C-0和E-0模型均有5個統(tǒng)計量有顯著性差異(P<0.05),但E-0模型AIC值較小,依據(jù)AIC值最小和顯著差異個數(shù)最少的選取準(zhǔn)則,確定楊凌開花時間性狀最適遺傳模型為E-0(2對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型)。三原點E-1模型有4個達(dá)到顯著差異而E-3模型有8個達(dá)到顯著差異,所以確定三原開花時間性狀最適遺傳模型為E-1(2對加性-顯性-上位性主基因+加性-顯性多基因模型)。
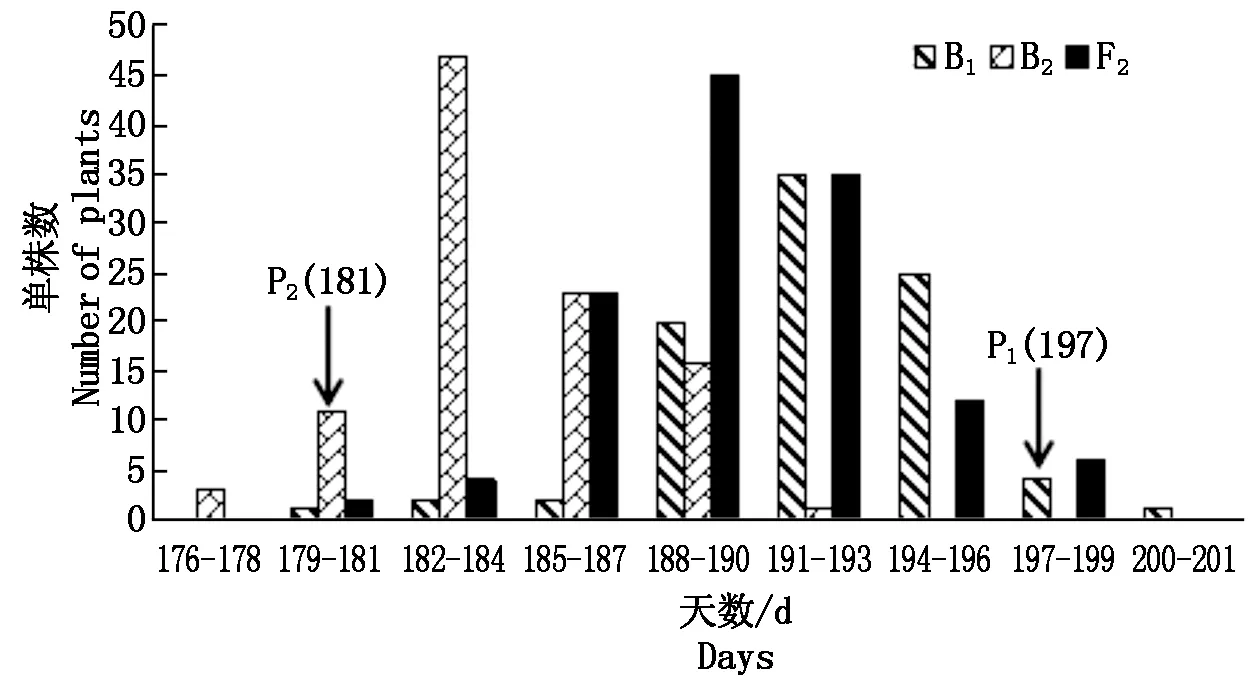
圖1 楊凌點B1、B2和F2群體開花時間次數(shù)分布Fig.1 Frequency distribution of flowering time traits in B1,B2 and F2 populations of Yangling
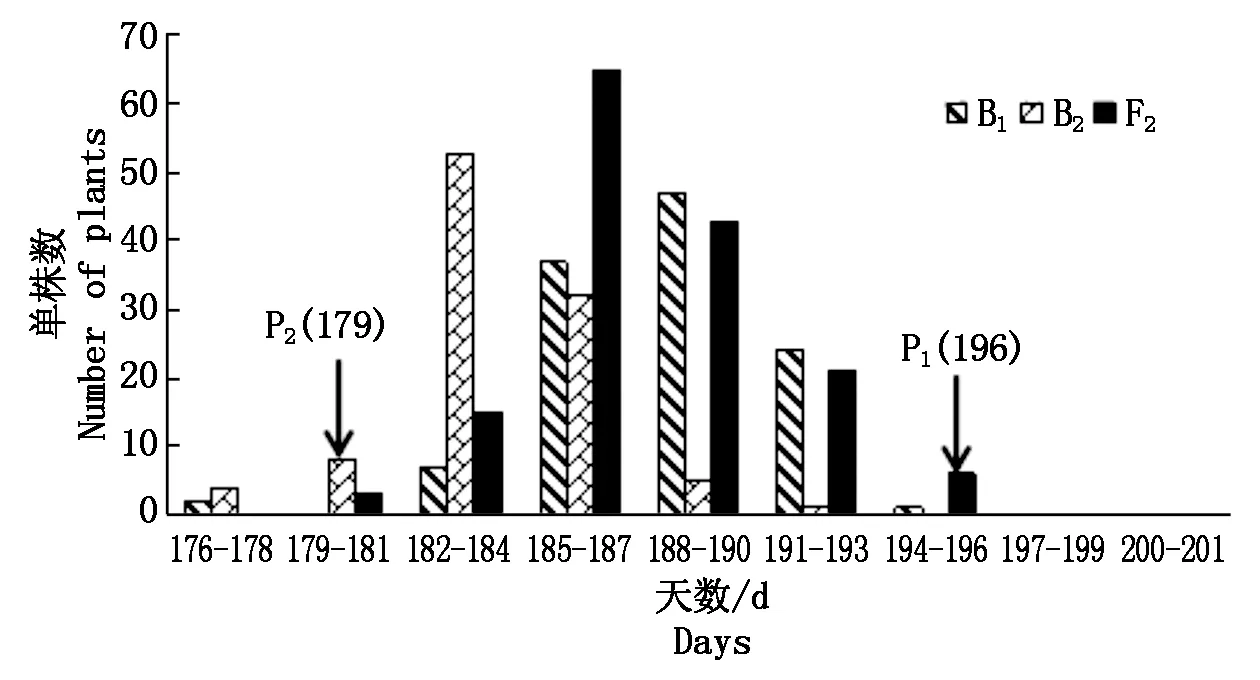
圖2 三原點B1、B2和F2群體開花時間次數(shù)分布Fig.2 Frequency distribution of flowering time traits in B1,B2 and F2 populations of Sanyuan
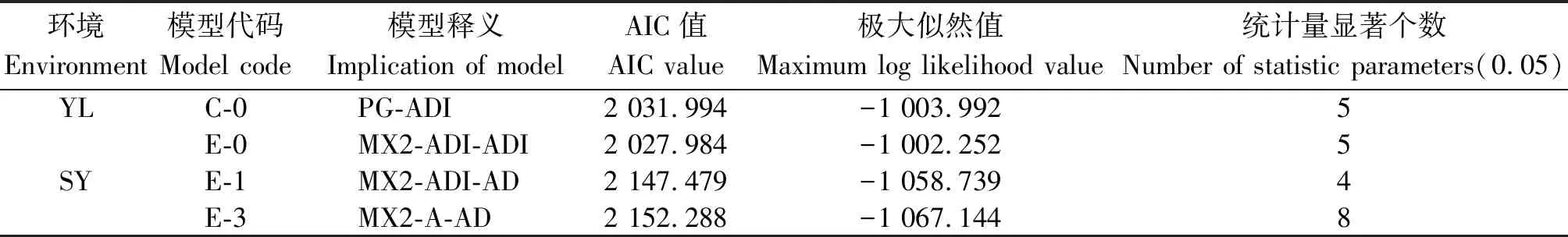
表3 備選模型相關(guān)參數(shù)Tab.3 Estimates on candidate models
注:MG、PG和MX分別表示主基因、多基因及主基因+多基因模型;A、D和I分別代表加性、顯性和上位性效應(yīng)。
Note:MG,PG and MX represent major gene, polygene and mixed major gene and polygene model;A,D and I represent additive,dominant and epistatic.
2.2.2 最適遺傳模型遺傳參數(shù)估算 按最小二乘法計算出成分分布參數(shù),由成分分布參數(shù)估計遺傳參數(shù)(表4)。由表4知,同一環(huán)境下2對主基因加性效應(yīng)值(|da|=|db|)相等,說明雙親中控制開花的2對主基因加性效應(yīng)相同,但地區(qū)間差異較大,分別為0.68和1.64。兩地2對主基因顯性效應(yīng)值均表現(xiàn)|hb|>|ha|,即第2個主基因的顯性效應(yīng)大于第一個主基因的顯性效應(yīng),顯性與加性的比值(|ha/da|和|hb/db|)多大于1,說明控制開花的2對主基因以顯性效應(yīng)為主,結(jié)合表1雙親雜交F1開花時間偏向于早花親本,說明早花性狀對晚花性狀為部分顯性。同時從表4還可以看出,控制開花時間的2對基因存在一定程度的基因互作效應(yīng)(i、jab、jba和l)。
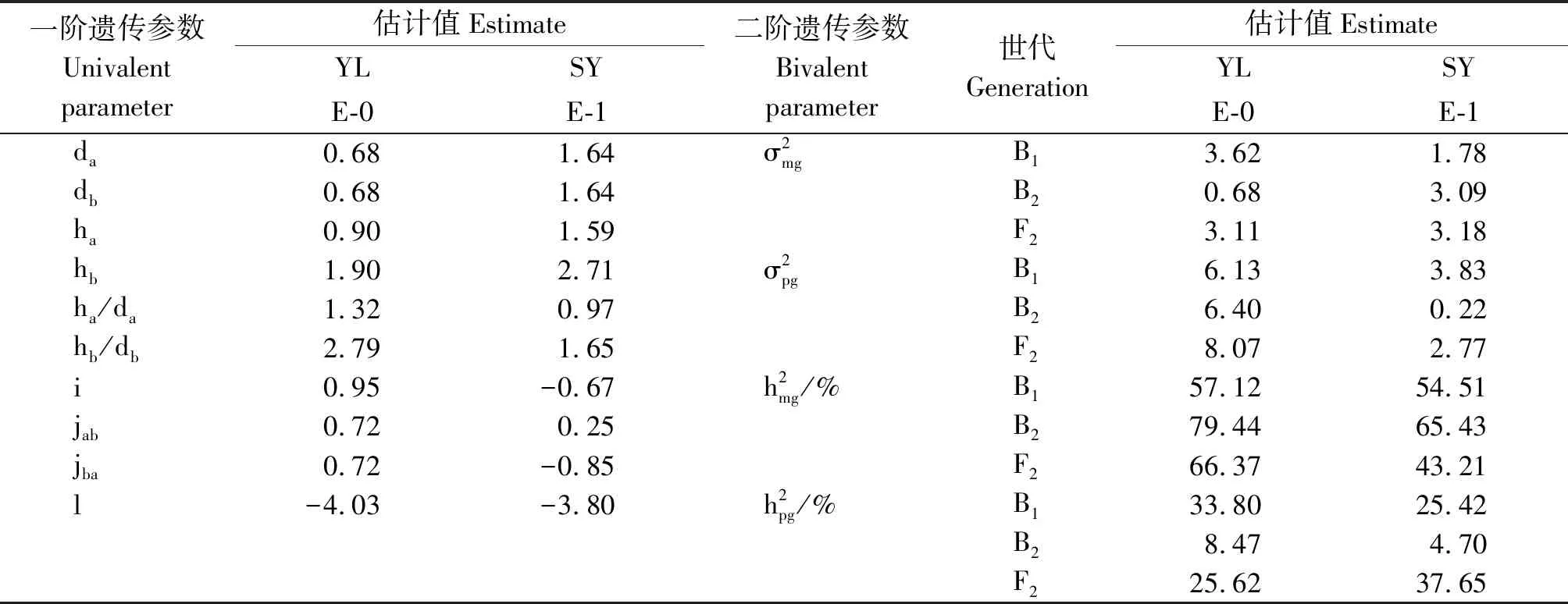
表4 E-0與E-1模型最適模型遺傳參數(shù)估計Tab.4 Estimates of genetic parameters under the E-0 and E-1 models
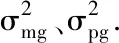
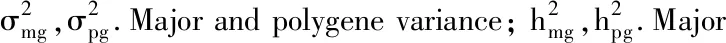
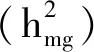
2.3 開花時間分子標(biāo)記
2.3.1 分子標(biāo)記篩選 利用507對SSR引物對雙親YG-1和72-27-1-2進(jìn)行多態(tài)性篩選,結(jié)果105對引物在雙親間存在多態(tài)性,占引物總數(shù)的20.7%,部分引物的篩選結(jié)果見圖3。利用在親本間存在差異的引物進(jìn)行F2群體(127個單株)的多態(tài)性篩選,在50對引物中重復(fù)檢測出多態(tài)性,多態(tài)率為47.6%,獲得帶型清晰的標(biāo)記位點54個。圖4顯示了SSR引物BnGMS148在前60個F2單株中的擴(kuò)增結(jié)果。

圖3 部分SSR引物在親本間擴(kuò)增的結(jié)果Fig.3 The amplification results of SSR primers between two parents

圖4 引物BnGMS148在前60個F2單株中的擴(kuò)增結(jié)果Fig.4 The amplification results of the primer BnGMS148 in F2 population with first 60 lines
2.3.2 開花時間與分子標(biāo)記的相關(guān)性分析 對挑選的F2極端類型單株(極端早20株、極端晚20株)的開花時間與54個標(biāo)記位點進(jìn)行相關(guān)性分析,并將各標(biāo)記引物序列在甘藍(lán)型油菜數(shù)據(jù)庫進(jìn)行Blast比對,相關(guān)分析結(jié)果及引物所在染色體位置見表5,在54個SSR標(biāo)記位點中,有7個SSR標(biāo)記位點與開花時間呈顯著(P<0.05)或極顯著(P<0.01)相關(guān),分別為cnu_m157a、BrgMS351、BnGMS148、BnGMS256-1、BnGMS256-2、BnGMS327及BnGMS370-2,其中,BnGMS256-2、BnGMS327、BnGMS370-23個位點與開花時間呈極顯著(P<0.01)相關(guān)。通過序列比對,BrgMS351和BnGMS148位于A7上,cnu_m157a、BnGMS256-1、BnGMS256-2、BnGMS327和BnGMS370-2位于A9上。說明本研究中控制開花相關(guān)的位點可能位于A7和A9染色體上,且其開花時間可能是由2個QTLs共同控制,與上述數(shù)量模型遺傳分析開花時間由2對主基因控制結(jié)果一致。
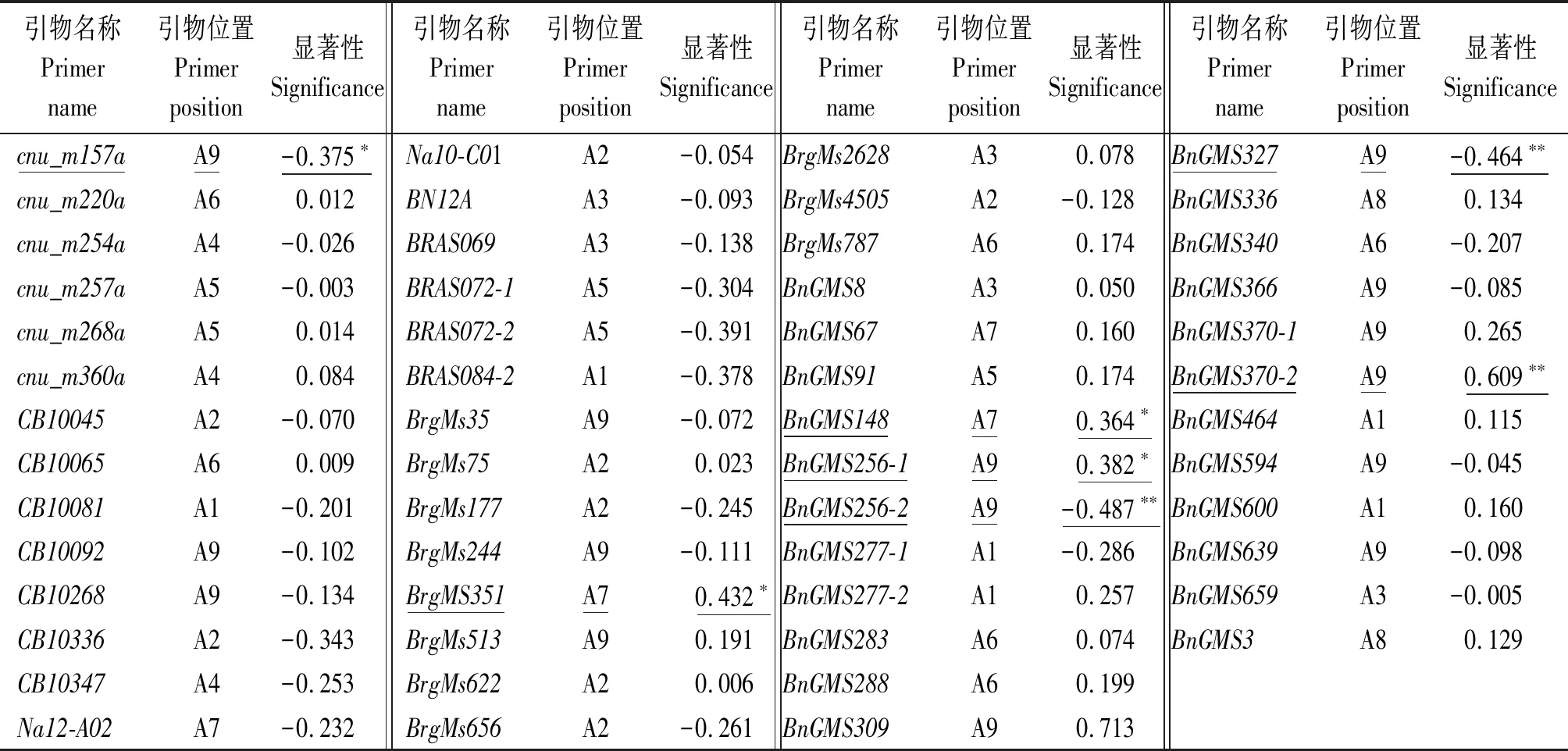
表5 F2群體開花時間與SSR 分子標(biāo)記相關(guān)性Tab.5 The correlation between flower time of F2population and SSR molecular markers
3 結(jié)論與討論
前人對植物數(shù)量性狀的遺傳研究,主要采用雙親本雜種或雙列雜交的世代雜種優(yōu)勢,運用遺傳方差、均值、配合力等分析其性狀的整體基因效應(yīng),但這些方法其結(jié)果僅能分析一組基因的綜合效應(yīng),并不能分析其單個基因的效應(yīng)[13]。蓋鈞鎰[14]將混合分布理論與數(shù)量遺傳相結(jié)合,建立了主基因+多基因混合遺傳模型,并提出了一套鑒定主基因與多基因存在且量化其效應(yīng)值的方法,克服傳統(tǒng)經(jīng)典數(shù)量遺傳方法的缺點。近年已應(yīng)用到各類作物的不同性狀上, 如甘藍(lán)型油菜莖稈強(qiáng)度[15]、株高[16]、分枝角度[17]、角果長度[18]、氮素營養(yǎng)效率[19]、黃瓜節(jié)間長[20]、胡麻粗脂肪[21]、高粱株型[22]、小菊分枝[23]等都獲得了很好的研究進(jìn)展。
郭志剛[2]發(fā)現(xiàn),油菜開花主要受多基因調(diào)控,但存在主基因加顯性效應(yīng),蔡長春[3]利用DH群體在和政、武漢和肇慶三點研究認(rèn)為油菜開花遺傳模型為G-1(3對加性-上位性主基因+多基因)和E-1(2對加性-上位性主基因+多基因),即開花時間受2對以上主基因控制,并受多對基因修飾效應(yīng),三點主基因遺傳率分別為91.13%,63.05%和62.02%,多基因遺傳率分別為4.43%,1.58%和22.71%。羅玉秀等[4]應(yīng)用傳統(tǒng)遺傳分析方法研究DH群體認(rèn)為開花期受2對加性基因控制,狹義遺傳率(h2G)為0.68。至此,甘藍(lán)型油菜開花時間未見更多報道。隨著章元明教授對數(shù)量遺傳模型軟件的不斷優(yōu)化,對分析過程中遺傳參數(shù)的估算更加精確。本研究應(yīng)用最新分析軟件(更新于2018年5月22日),對6世代群體開花時間進(jìn)行遺傳分析,兩地試驗結(jié)果表明,甘藍(lán)型油菜開花時間適應(yīng)E-0與E-1模型,即主要受2對主基因控制,主基因遺傳率大于相應(yīng)世代多基因遺傳率,表明大田育種中開花時間性狀適宜進(jìn)行早代選擇。本研究楊凌點開花時間最適遺傳模型E-1與蔡長春[3]研究結(jié)果保持一致,兩點開花時間受2對主基因控制與羅玉秀等[4]2對主基因研究結(jié)果一致,只是加顯性效應(yīng)不同,究其原因可能與試驗材料不同有關(guān)。這表明不同試驗材料,控制油菜開花的基因數(shù)目與效應(yīng)可能不同。
本試驗構(gòu)建了可用于QTL與相關(guān)性分析的F2群體,并利用127個單株進(jìn)行研究,排除了遺傳背景不同和群體大小對相關(guān)性分析的影響,進(jìn)行標(biāo)記與性狀之間的相關(guān)性分析。而標(biāo)記與性狀的相關(guān)性分析是以多態(tài)性標(biāo)記位點與目標(biāo)數(shù)量性狀的表型進(jìn)行分析,若相關(guān)性顯著或極顯著則說明標(biāo)記與性狀存在相關(guān)性。所以,若一個群體的性狀差異明顯,就可以通過標(biāo)記與性狀的相關(guān)性找出性狀與1個或多個標(biāo)記的遺傳相關(guān)性,若存在顯著相關(guān),就可以認(rèn)為存在1個數(shù)量性狀位點,從而實現(xiàn)表型到基因型的轉(zhuǎn)變[24]。本試驗中共關(guān)聯(lián)到7個SSR標(biāo)記位點與開花時間具有顯著或極顯著相關(guān)。標(biāo)記引物序列在甘藍(lán)型油菜數(shù)據(jù)庫進(jìn)行比對,分別位于A7和A9上,這表明本研究材料控制開花相關(guān)位點可能位于A7和A9染色體,也說明開花時間可能是由2個QTL共同控制。Butruille等[25]利用冬油菜與春油菜雜交和回交群體,在A2、A3、A7、A8、A9、C2和C5染色體上定位到開花期QTL,Lou等[26]利用IBM211與R500雜交創(chuàng)制的RILs群體,在A2、A3、A7、A9和A10定位到6個QTLs,柳海東[8]利用DH系在5個環(huán)境中共檢測到48個QTLs與開花時間有關(guān),分別位于染色體A2、A3、A5、A6、A7、A10、C2、C8上,本試驗所得的與開花時間相關(guān)的遺傳標(biāo)記分別位于A7和A9上,與前人研究結(jié)果比較找到的標(biāo)記較少,其原因可能主要是所用的SSR標(biāo)記數(shù)量較少,進(jìn)一步深入研究還需擴(kuò)大群體,采用更多的標(biāo)記引物,獲得更多距離目標(biāo)基因更近的分子標(biāo)記或主效QTL。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
