利用高通量測序快速定位目標(biāo)性狀位點的研究
2020-01-07 09:58:56趙青松張孟臣楊春燕趙團(tuán)結(jié)
華北農(nóng)學(xué)報 2019年6期
關(guān)鍵詞:大豆
林 靜,趙青松,張孟臣,楊春燕,趙團(tuán)結(jié)
(1.河北省農(nóng)林科學(xué)院 糧油作物研究所,國家大豆改良中心石家莊分中心,農(nóng)業(yè)部黃淮海大豆生物學(xué)與遺傳育種重點實驗室, 河北省農(nóng)作物遺傳育種重點實驗室,河北 石家莊 050035;2.南京農(nóng)業(yè)大學(xué)國家大豆改良中心,農(nóng)業(yè)部大豆生物學(xué)與遺傳育種 重點實驗室(綜合),國家作物遺傳與種質(zhì)資源重點實驗室,江蘇 南京 210095)
大豆(GlycinemaxL.)是我國主要的糧、油、飼兼用作物。隨著我國經(jīng)濟(jì)水平發(fā)展和人民生活水平的日益提高,我國對大豆的需求總量與日俱增。據(jù)報道,我國近幾年大豆消費量維持在1億t以上,而我國大豆年生產(chǎn)量僅為1 200萬t左右(http://www.stats.gov.cn/),自給率不足20%,大豆嚴(yán)重依賴進(jìn)口的現(xiàn)狀對我國糧食安全造成巨大的隱患[1-3]。目前,我國大豆單產(chǎn)水平在1.8 t/hm2左右,而大豆前三大生產(chǎn)國美國、阿根廷和巴西的單產(chǎn)水平分別為3.51,3.02,2.91 t/hm2[4],因此,我國大豆單產(chǎn)有很大的提升空間,通過提高我國大豆單產(chǎn)水平,可以一定程度上緩解對進(jìn)口大豆的依賴性。
常規(guī)的作物育種需要通過雜交、選育、株行觀察、加代穩(wěn)定、產(chǎn)量鑒定、產(chǎn)量比較、區(qū)試和生產(chǎn)試驗等步驟[5],整個育種周期需要5~10 a。常規(guī)品種選育一般至少需要5代以上,要求主要農(nóng)藝性狀穩(wěn)定。然而,在實際選育過程中,即使到F5以上,選育株行中仍會出現(xiàn)農(nóng)藝性狀分離,其對育種家造成很大的困擾。隨著分子技術(shù)的發(fā)展,分子輔助育種(Marker assistance selection, MAS)被應(yīng)用到現(xiàn)代育種當(dāng)中,利用與性狀關(guān)聯(lián)的分子標(biāo)記可以快速篩選出有利于生產(chǎn)的純合性狀[6],不僅減輕了田間工作量,并且顯著縮短了育種周期。因此,鑒定與目標(biāo)性狀關(guān)聯(lián)的分子標(biāo)記,即定位調(diào)控目標(biāo)性狀的關(guān)鍵位點是分子輔助育種的基礎(chǔ)。
目前,常用基因定位的遺傳群體有F2、RIL、DHL和自然群體等。其中F2群體為臨時群體,一般用于定位質(zhì)量性狀位點(Qualitative trait locus)效果較好[7-9], RIL和DHL則為穩(wěn)定遺傳的永久性群體,適用于需要多重復(fù)的數(shù)量性狀位點(Quantitative trait locus, QTL)的定位[10-11]。自然群體屬于非人工構(gòu)建的,經(jīng)過自然馴化而成的群體,其優(yōu)點是含有豐富的遺傳多樣性,而缺點則是不適用于鑒定微效的QTL位點[12-13]。另外,自然群體往往受群體大小的限制,多種情況下仍然難以獲得調(diào)控目標(biāo)性狀關(guān)鍵基因的精確位置。因此圖位克隆(Map-based cloning)往往需要擴(kuò)大遺傳群體。而數(shù)量性狀由于受遺傳背景的影響,需衍生出次級群體,例如剩余雜合系(Residual hybrid lines, RHL)和近等基因系(Near-isogenic line, NIL)等,進(jìn)而進(jìn)行精細(xì)定位和圖位克隆,目前RHL和NIL已經(jīng)廣泛用于水稻、玉米、小麥和大豆等作物的精細(xì)定位研究[14-17]。
圖位克隆從方法和策略上已日趨成熟,但構(gòu)建基本遺傳群體、鑒定性狀和構(gòu)建次級群體,過程繁瑣且耗時耗力。尤其是次級群體RHL和NIL的構(gòu)建往往需要連續(xù)回交或自交至4~5代以上,才可以將90%以上的遺傳干擾去除。而育種過程中,F(xiàn)5以上的單株行如發(fā)生性狀分離,其本身就包含RHL和NIL 2種特性,但由于檢測技術(shù)手段的限制往往被忽略。基于以上研究現(xiàn)狀,本研究利用高世代的中間材料,以調(diào)控大豆花色的基因位點W1為例,在田間F6株行中篩選花色分離的家系,運用近等基因系原理和重測序技術(shù)快速挖掘調(diào)控目標(biāo)性狀的遺傳位點,進(jìn)而闡釋本策略的有效性,結(jié)果將提供一種可以快速挖掘調(diào)控目標(biāo)性狀遺傳位點的定位策略,以加速育種進(jìn)程。
1 材料和方法
1.1 試驗材料及性狀鑒定
試驗材料選用大豆MS輪回群體中364個F6株行,于2017年在河北石家莊堤上實驗站進(jìn)行種植,行長6 m,每行播種50~100粒,在開花期對株行的花色進(jìn)行統(tǒng)計觀察,對出現(xiàn)分離的株行進(jìn)行單株收獲形成F6∶7。分別取F6∶7每個家系的種子20~30粒,于2018年繼續(xù)進(jìn)行株行觀察花色分離情況。
1.2 DNA樣品采集與重測序方法
2017年對出現(xiàn)分離的株行單株進(jìn)行掛牌,每單株取1~2片嫩葉凍存?zhèn)溆谩?018年對F6∶7每家系進(jìn)行觀察,統(tǒng)計分離情況。在每個不發(fā)生分離的株行中隨機(jī)掛牌1株,取1~2片嫩葉,并將株行表型一致的葉片混合凍存?zhèn)溆谩?/p>
重測序在廣州基迪奧生物科技有限公司進(jìn)行。具體方法如下:利用商用基因組試劑盒提取植株DNA。運用NEB建庫試劑盒(NEBNext? ΜLtraTMDNA Library Prep Kit for Illumina?)構(gòu)建測序文庫。測序均在Illumina測序平臺進(jìn)行。將去除接頭后的測序數(shù)據(jù)和參考基因組Wm82.a2.v1進(jìn)行比對,參數(shù)設(shè)置為“mem 4-k 32-M”[18],SNP變異運用GATK′s Unified Genotyper軟件進(jìn)行分析。
1.3 dCAPS標(biāo)記開發(fā)和檢測方法
在重測序結(jié)果基礎(chǔ)上,利用鑒定到的SNP變異開發(fā)目標(biāo)區(qū)間的dCAPs標(biāo)記。具體流程方法如下:明確在目標(biāo)區(qū)段的SNP變異位點,在Phytozome v.12(https://phytozome.jgi.doe.gov/pz/portal.html)下載SNP變異位點上下游各30 bp的DNA序列,將野生型和突變類型的DNA序列同時輸入至網(wǎng)站http://helix.wustl.edu/dcaps/dcaps.html進(jìn)行引物設(shè)計,并在F5植株的DNA中進(jìn)行多態(tài)性驗證。將F5植株的DNA提取后利用200 μL 1×TE Buffer進(jìn)行稀釋。PCR反應(yīng)液根據(jù)TaKaRa 的PrimerSTAR Max DNA Polymerase試劑盒說明進(jìn)行配制。PCR反應(yīng)條件:95 ℃ 10 min;92 ℃ 30 s,52 ℃ 30 s,72 ℃ 1 min,35個循環(huán);70 ℃ 10 min。SSR引物的PCR產(chǎn)物銀染后在8%聚丙烯酰胺凝膠上進(jìn)行電泳拍照記錄,dCAPS引物的PCR產(chǎn)物利用對應(yīng)的酶進(jìn)行酶切后,在3%瓊脂糖凝膠上進(jìn)行電泳拍照記錄。
1.4 花色基因連鎖定位
分別與紫花或白花DNA池一致的多態(tài)標(biāo)記分別記為“a”或 “b”,不清楚或偏分離的記為“-”。表型為紫花和白花的分別記為“d”和“b”,利用Join Map 4.1 內(nèi)置算法Regression algorithm 和 Kosambi函數(shù)對進(jìn)行遺傳圖譜的構(gòu)建。運用MapChart 2.2軟件進(jìn)行圖譜繪制[19]。
2 結(jié)果與分析
2.1 花色性狀遺傳分析
MS輪回群體的F6于2017年進(jìn)行夏播,行長6 m,在開花期調(diào)查其花色性狀,發(fā)現(xiàn)1個花色分離的株行。75個單株有58個表現(xiàn)為紫花,17個白花,符合3紫花∶1白花的表型分離比例(P>0.05) (表1);對75個單株衍生的F6∶7家系。2018年種植75個單株衍生的F6∶7家系,結(jié)果顯示,17個白花的后代株行均表現(xiàn)為白花;58個紫花的后代株行中,20個株行未出現(xiàn)花色性狀分離,38個株行出現(xiàn)花色性狀分離,符合1紫花∶2花色分離∶1白花的預(yù)期分離比例(P>0.05)。結(jié)果表明,在本研究群體中大豆花色性狀由單基因位點控制,其中紫花對白花為顯性性狀,研究結(jié)果與以往研究結(jié)果一致[20]。

表1 大豆花色的遺傳分析Tab.1 The genetic analysis of soybean flower color
2.2 重測序變異基本分析
2018年在 MS輪回群體F6∶7的20個和17個純紫花或白花株行中隨機(jī)取1株采集葉片,將紫花和白花分別混成樣品池,提取其DNA進(jìn)行高通量測序,測序數(shù)據(jù)量為10 Gb。對樣品池間SNP變異進(jìn)行比對分析,共檢測到329 992個SNP變異位點。由基因功能區(qū)分布可知(圖1-A),其中252 651個(占比76.56%)SNP變異位點分布于基因間隔區(qū)(Intergenic region)。可變剪切區(qū)(Splicing region)中SNP變異位點分布最少,僅為81個(占比0.02%)。此外,3個SNP變異位點分別跨5′UTR和3′UTR區(qū)域,4個SNP變異位點位于外顯子的可變剪切區(qū)。由SNP變異位點在整個基因組和染色體的分布可知(圖1-B、C),SNP變異位點在20條染色體上分布相對均勻,占比在4.5%左右。其中,20號染色體富集最多,有32 341個SNP變異位點(占比9.80%),11號染色體最少,僅含有5 186個(占比1.57%)SNP變異位點,此外,不同染色體區(qū)段存在多個明顯SNP變異位點富集區(qū),表明這些區(qū)段可能為大豆花色調(diào)控基因的候選區(qū)段。但此結(jié)果仍無法判斷調(diào)控大豆花色位點的區(qū)間,因此需要進(jìn)一步篩選。
2.3 花色候選區(qū)段篩選
為進(jìn)一步明確大豆花色調(diào)控基因的候選區(qū)間,針對原始測序結(jié)果進(jìn)行2步篩選:①選出質(zhì)量較高的SNP(Quality>100)變異位點;②去除雜合位點。篩選后統(tǒng)計結(jié)果如下:共獲得3371個高質(zhì)量且純合的SNP變異位點。由基因功能區(qū)分布可知(圖2-A),其中2 424個(占比71.93%)SNP變異位點分布在基因間隔區(qū)(Intergenic region),可變剪切區(qū)(Splicing region)分布最少,僅為2個(占比0.06%)。由SNP變異位點在整個基因組和染色體分布可知(圖2-B、C),SNP變異位點在20條染色體上分布比例為0.98%~20.77%。其中,13號染色體富集最多,有700個SNP變異位點(占比20.77%),表明調(diào)控大豆花色的基因最可能位于13號染色體上(圖2-C)。進(jìn)一步分析其富集的區(qū)段,結(jié)果顯示,在13號染色體20~30 Mb的物理區(qū)間有顯著的SNP富集區(qū)(圖2-B),表明該SNP變異位點富集區(qū)為強(qiáng)候選區(qū)間。

圖1 篩選前SNP變異位點分布Fig.1 The original distribution of SNP variation before screening
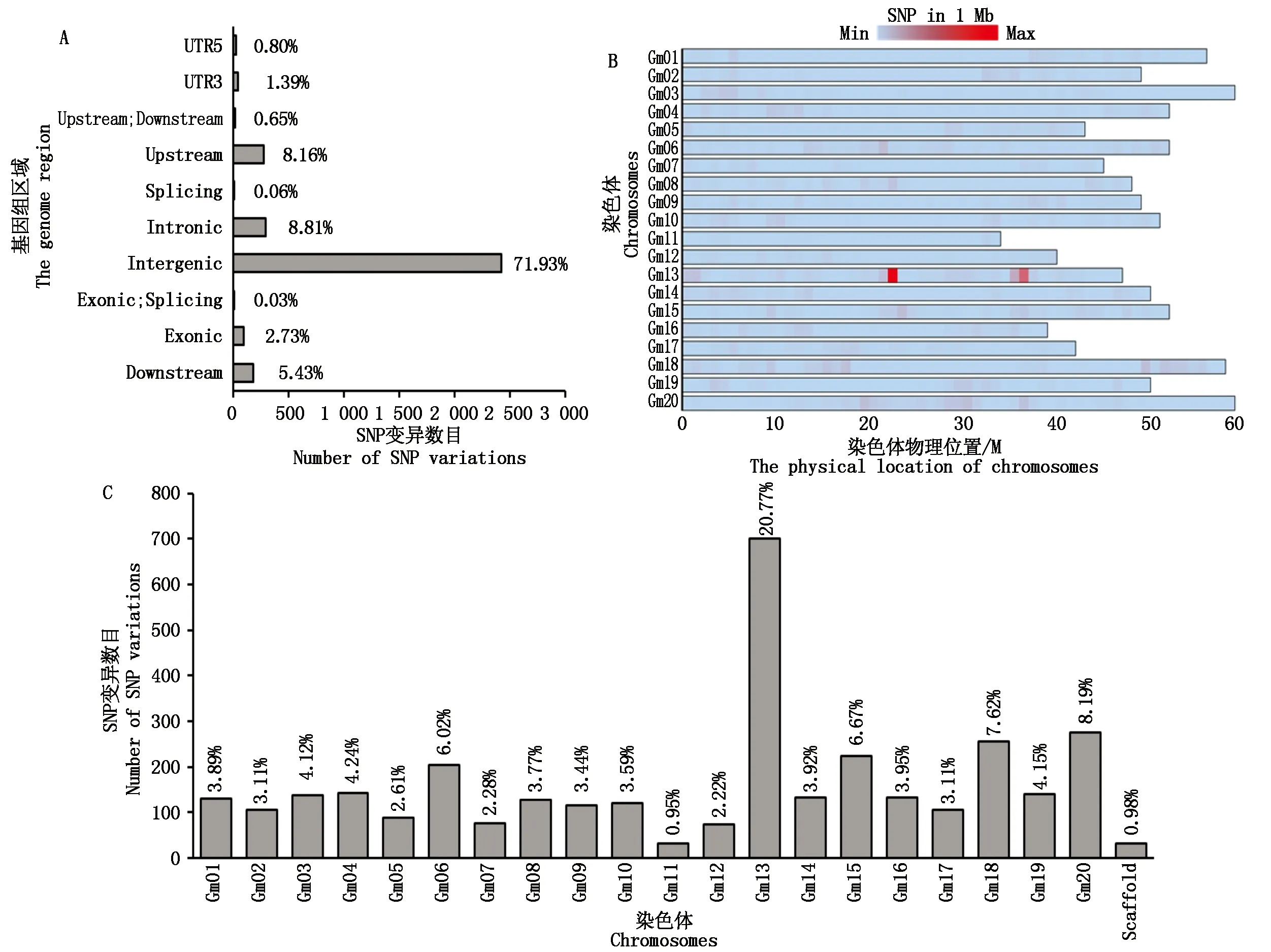
圖2 篩選后SNP變異位點分布Fig.2 The distribution of SNP variation after screening
2.4 dCAPs 標(biāo)記的開發(fā)和調(diào)控大豆花色基因位點的連鎖分析
為進(jìn)一步證實結(jié)果的可信性,在候選區(qū)間物理位置21 931 375和21 948 774處分別開發(fā)了2對dCAPS標(biāo)記(表2),同時為了防止設(shè)計標(biāo)記出現(xiàn)異位擴(kuò)增現(xiàn)象,同時篩選出了2對具有多態(tài)性的SSR標(biāo)記Satt149和Satt516作為參照。進(jìn)一步利用F6株行的75個單株和4個多態(tài)標(biāo)記進(jìn)行連鎖分析。結(jié)果顯示(圖3), dCAPS-1、dCAPS-2、W1、Satt149和Satt516之間均呈現(xiàn)連鎖,遺傳距離為Satt149-(8.8 cM)-W1-(0.4 cM)-dCAPS-1-(2.3 cM)-dCAPS-2-(8.7 cM)-Satt516,整個遺傳圖譜共可覆蓋20.2 cM的遺傳距離,其中2個dCAPS標(biāo)記位于W1的同一側(cè),遺傳距離分別為0.4,2.7 cM。結(jié)果表明,新開發(fā)的dCAPS標(biāo)記未出現(xiàn)異位擴(kuò)增,可以代表目標(biāo)位置的多態(tài)性。此外,新開發(fā)的標(biāo)記均與目標(biāo)性狀緊密連鎖,且定位W1所在的位置與以往報道一致[21],表明新開發(fā)的dCAPS標(biāo)記可以用于后續(xù)的MAS中。

表2 本研究中使用的dCAPS引物Tab.2 The dCAPS primers used in this study
注:引物中下劃線的堿基表示引入的突變堿基位點。
Note:The underlined font indicates the position of introduced SNP variation.
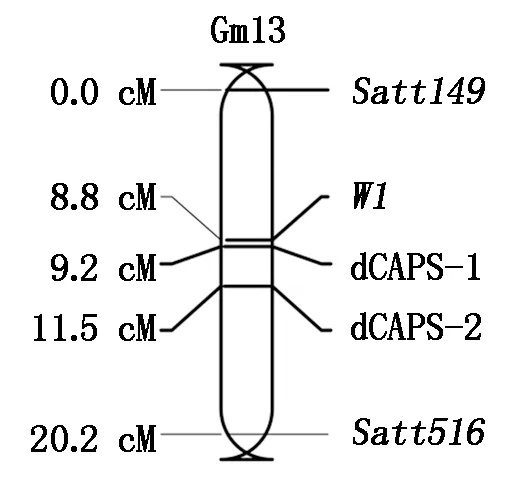
圖3 花色位點W1的連鎖圖譜Fig.3 The genetic map of W1 for flower color
3 討論
群體分離分析(Bulk segregation analysis, BSA)是一種可用于快速定位質(zhì)量性狀或含有主效位點的數(shù)量性狀的研究方法,在水稻、小麥、玉米和大豆等作物的遺傳定位的研究中被廣泛使用[22-24],通常使用F2或RIL群體進(jìn)行混池的構(gòu)建。其中F2群體構(gòu)建比較簡單快速,但對于質(zhì)量性狀而言,子代池中顯性性狀池會混入雜合基因系,對定位效果存在很大干擾;雖然利用RIL群體材料可以很好解決這個問題,但RIL群體構(gòu)建周期較長,并且對數(shù)量性狀定位的效果并不十分理想。對數(shù)量性狀位點的精細(xì)定位需要構(gòu)建更高級的遺傳群體,例如剩余雜合系和近等基因系。花色性狀在育種過程中是非常容易區(qū)分的一個質(zhì)量性狀,盡管沒有證據(jù)表明對大豆產(chǎn)量具有顯著影響,但在品種審定的過程中,花色分離往往會被淘汰。在田間育種的過程中,高世代株行發(fā)生花色分離是一種常見的現(xiàn)象,嚴(yán)重影響了育種的效率。本研究從解決田間高世代株行材料發(fā)生花色分離的現(xiàn)象入手,將高世代F6∶7中不分離的株行構(gòu)建成紫花和白花DNA池,采用重測序的辦法進(jìn)行定位研究。由于使用了高世代材料,材料間遺傳背景相似度高,材料間屬于近等基因系,極大排除了遺傳背景的干擾,易于鑒定。結(jié)果也顯示,篩選出的SNP標(biāo)記形成了明顯的富集區(qū),在富集區(qū)開發(fā)的dCAPS標(biāo)記可以很好地追蹤花色性狀,表明這2個標(biāo)記可以應(yīng)用到分子標(biāo)記輔助育種當(dāng)中。
大豆是古四倍體作物,約在5 900萬a和1 300萬a之前經(jīng)過2次染色體加倍事件[25],隨后經(jīng)歷染色體丟失、重排、分化等事件逐漸形成現(xiàn)代的2倍體大豆。因此,大豆基因間序列具有高度的同源性,因此一個基因可能存在多個拷貝的現(xiàn)象。而不同拷貝間的變異又是相對獨立的,因此不同拷貝間也存在變異。目前主流的重測序技術(shù)是二代重測序,主要以在50~250 bp的DNA片段加接頭的方式進(jìn)行高通量的測序,獲得原始數(shù)據(jù)與參考基因組進(jìn)行比對分析來確定遺傳變異[26]。但受限于參考基因組拼接質(zhì)量和基因組中多拷貝變異之間的影響,往往會鑒定出大量假陽性的SNP變異位點,這種假陽性率在大豆、小麥等古四倍體或多倍體作物中尤為高。而一般測序比對默認(rèn)的篩選參數(shù)較低[27],因此加大了假陽性出現(xiàn)的概率。在研究中發(fā)現(xiàn),根據(jù)測序通用標(biāo)準(zhǔn)在紫花和白花2個混池中僅鑒定到約33萬個SNP變異位點,說明從F6∶7株行的植株間遺傳背景已經(jīng)非常相近。但這33萬個SNP變異位點未呈現(xiàn)明顯的富集區(qū),不能明確調(diào)控花色位點的準(zhǔn)確遺傳位置,推測存在較多的假陽性位點。因此,通過提高測序質(zhì)量(Quality 參數(shù)),進(jìn)一步提高位點真實性。此外,構(gòu)建紫花和白花使用的單株均為花色純合單株,因此與目標(biāo)位點連鎖的區(qū)域也趨于純合,因此進(jìn)一步將呈現(xiàn)雜合的位點排除。最終得到3 371個候選位點,并且在13號染色體處形成了一個明顯的富集區(qū),說明未篩選前數(shù)據(jù)確實存在大量假陽性的SNP變異。
本研究利用田間高世代育種材料和高通量測序技術(shù),對大豆花色調(diào)控基因位點W1進(jìn)行了快速定位。研究結(jié)果顯示,利用高世代分離材料可用于有效、快速排除遺傳背景的干擾,提高了遺傳定位效率,在W1位點候選區(qū)間內(nèi)開發(fā)的分子標(biāo)記可以有效地跟蹤W1的遺傳走向,研究結(jié)果為今后的MAS育種提供了理論參考基礎(chǔ)。
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
