玉米皮纖維發酵培養裂褶菌的轉錄組分析和重組α-L-阿拉伯呋喃糖苷酶的異源表達
2020-01-13 10:45:56劉玉春郭超郭偉群
食品與發酵工業 2019年23期
劉玉春,郭超,郭偉群
(國家糧食和物資儲備局科學研究院,北京,100037)
玉米皮是玉米深加工的主要副產物之一,占玉米籽粒質量的15%左右[1],玉米皮中半纖維素含量可達到68%[2],與其他植物來源的半纖維素相比,其具有支鏈化程度高的特點,其主鏈的每一個木聚糖殘基幾乎都連接有支鏈殘基[3]。玉米皮纖維的生物降解涉及到多種酶的協同作用,包括木聚糖的主鏈降解酶——木聚糖酶(xylanase)、木糖苷酶(xylosidase)以及多種支鏈降解酶[4]。目前,玉米皮多作為廉價的飼料出售,為提高其利用價值,已有應用玉米皮發酵培養黑曲霉[5]、酵母菌發酵玉米皮制備菌體蛋白[6]、玉米皮發酵培養琥珀酸放線桿菌產丁二酸[7]等方面研究。
裂褶菌(Schizophyllumcommune)具有較強的半纖維素和纖維素的分解能力,能夠在含有半纖維素的基質上良好地生長[8]。本課題組在前期研究中,已經測定了玉米皮纖維發酵培養裂褶菌的產酶情況,包括木聚糖酶、乙酰木聚糖酯酶、α-L-阿拉伯呋喃糖苷酶和α-葡萄糖醛酸酶[9];同時完成了1個木聚糖酶基因的克隆和異源表達[9]。但與玉米皮木聚糖連接的阿拉伯糖支鏈抑制了木聚糖水解酶的活性,而α-L-阿拉伯呋喃糖苷酶(α-L-arabinofuranosidase, EC 3.2.1.55)能從含有阿拉伯糖殘基的多聚物的非還原末端水解1個阿拉伯糖分子,該酶同木聚糖酶復合后具有協同作用,能夠促進玉米皮木聚糖降解。因此,本研究設計克隆裂褶菌α-L-阿拉伯呋喃糖苷酶基因,并完成其在畢赤酵母中的異源表達。
真菌轉錄組的測序分析具有速度快、信息全面的優點,能夠全面分析特定培養條件下基因轉錄的情況[10]。例如,對灰樹花的菌絲體進行轉錄組測序分析,獲得了該菌株生長過程中參與多糖合成的基因的表達情況[11]。與真菌的全基因組測序分析相比,轉錄組的測序結果中不含內含子及其他非編碼的核酸序列,可以更高效地分析與培養條件相關的基因轉錄情況。
本研究是以玉米皮纖維為唯一碳源發酵培養裂褶菌為樣本,通過轉錄組測序技術分析與玉米皮纖維降解相關基因的轉錄情況;同時克隆了裂褶菌糖苷水解酶62家族蛋白基因Sabf32,并完成該基因在畢赤酵母中的表達,研究為木聚糖酶與α-L-阿拉伯呋喃糖苷酶協同降解玉米皮纖維提供了基礎,同時為玉米皮的增值利用提供了理論基礎。
1 材料和方法
1.1 材料
1.1.1 菌株
裂褶菌S.communeDB01,本實驗室篩選并保存。
1.1.2 主要試劑
D-木糖、樺木木聚糖,Megazyme公司;PDA培養基,北京陸橋技術股份有限公司;3,5二硝基水楊酸、酒石酸鉀鈉、NaOH,國藥集團化學試劑有限公司;蛋白質濃度測定試劑盒,北京索萊寶科技有限公司;對硝基苯阿拉伯呋喃糖苷(4-nitrophenyl α-L-arabinofuranoside,pNPAF),Sigma公司。
1.1.3 主要儀器
DK-8D電熱恒溫水槽,上海精宏實驗設備有限公司;PL2002IC電子天平,Mettler公司;MQD-SIR單層小容量振蕩培養箱,上海旻泉儀器有限公司;SynergvHT酶標儀,美國BioTek公司;ND OneCNanoDropTM微量紫外-可見光分光光度計,Thermo Fisher公司;電泳儀、凝膠成像儀,美國Bio-Rad公司。
1.2 實驗方法
1.2.1 培養條件
玉米皮纖維誘導培養基成分[12](g/L):蛋白胨1、尿素0.2、CaCl20.3、MgSO4·H2O 0.3、KH2PO42、(NH4)2SO44.2、0.2%(v/v)微量元素(CoCl21.6、MnSO4·4H2O 1.6、ZnSO4·7H2O 1.4、FeSO4·7H2O 5),玉米皮纖維10,作為碳源。
菌株經PDA平板活化后,將裂褶菌接種于300 mL玉米皮纖維培養基中,30 ℃、220 r/min培養120 h。在培養過程中,取發酵液作為檢測樣品,取樣過程中應盡量避免振蕩,將發酵液于12 000×g離心5 min后取上清液待測。
1.2.2 發酵液中蛋白質、還原糖和酶活力測定
發酵液還原糖含量測定:將100 μL上清液加水稀釋至200 μL,加入300 μL DNS試劑,混合后置于沸水浴5 min,迅速冷卻后在540 nm波長下測定吸光值。
木聚糖酶活性測定[13],反應體系為:100 μL適量稀釋的發酵液上清、100 μL底物(10 g/L,樺木木聚糖),在40 ℃保溫10 min,然后加入300 μL試劑終止反應,沸水煮5 min并冷卻,在540 nm波長下測定吸光值。
蛋白質濃度測定:根據蛋白質濃度測定試劑盒說明書測定。
1.2.3 裂褶菌總RNA提取及測序
菌株經PDA平板活化后,接種玉米皮纖維培養基,于30 ℃,220 r/min條件下培養48 h。抽濾液體培養基得到菌絲體,稱取0.1~0.3 g菌絲體于研缽中,加入液氮,并在液氮保護下快速研磨,樣品研磨至無顆粒呈粉狀,將研磨好的粉末移入裝有Trizol的離心管中。用Trizol法提取裂褶菌總RNA,并用瓊脂糖凝膠電泳分析RNA的完整性。應用微量紫外-可見光分光光度計測定樣品濃度及純度(A260/A280 nm及A260/A230 nm)。
1.2.4 數據分析及注釋分類
Illumina HiSeq測序后對原始序列進行過濾(測序工作由安諾優達基因科技有限公司完成),用組裝軟件Trinity(20140717版)[14]組裝出全長轉錄本序列,將每個基因中最長的轉錄本作為Unigene。在NCBI數據庫(https://www.ncbi.nlm.nih.gov/)對Unigene進行NR(non-redundant protein sequence database;non-redundant protein sequences from GenPept, Swissprot, PIR, PDF, PDB, and NCBI RefSeq)、NT(nucleotide sequence database;all traditional divisions of GenBank, EMBL, and DDBJ excluding bulk divisions)數據庫的物種分布分析[16],同時進行BLASTP和BLASTX分析、統計、排序,確定數目前10的物種種類。其他能比對上的物種總稱為Others。
Unigene的GO注釋[17]是以基因NR注釋信息為基礎,獲得每個Unigene的注釋信息。對Unigene進行功能分類統計(WEGO),從生物過程(biological process)、細胞組成(cellular component)、分子功能(molecular function)3方面對裂褶菌Unigene的功能分布進行統計,并分析其分布特征。
1.2.5 α-L-阿拉伯呋喃糖苷酶基因的克隆
根據轉錄組測序結果,獲得α-L-阿拉伯呋喃糖苷酶基因Sabf32的序列信息,應用SignalP 4.0(http://www.cbs.dtu.dk/services/SignalP-4.0/)預測Sabf32的信號肽序列,設計畢赤酵母表達引物:EcoRIsAfF:5′-CCG GAATTC CAGGCGTGCGACCTCCCGT-3′;NotIsAfR:5′-ATAGTTTA GCGGCCGC AGCCTGGGTAAGCAAGCCGGG-3′,引物由生工生物工程(上海)股份有限公司合成。
應用限制性內切酶:EcoR I和NotI分別將質粒pPIC9K和Sabf32進行雙酶切;將酶切產物純化后,應用T4 DNA連接酶構建重組表達質粒pPIC9K-Sabf32。應用PCR擴增驗證陽性轉化子,進一步通過測序驗證核酸序列。
1.2.6 α-L-阿拉伯呋喃糖苷酶的畢赤酵母表達
應用電擊轉化將重組表達質粒pPIC9K-Sabf32和pPIC9K空載體轉化畢赤酵母GS115感受態細胞,篩選獲得陽性重組子。應用BMGY對重組子進行擴大培養,離心收集菌體后用BMMY培養基進行誘導培養;取誘導培養7 d的發酵液,7 000×g離心,取上清液進行SDS-PAGE電泳檢測(轉化、重組子篩選及發酵條件參照Invitrogen公司的畢赤酵母菌實驗操作手冊)。應用陰離子交換樹脂純化重組蛋白,并進行SDS-PAGE檢測和分析重組蛋白的分子量。
1.2.7 重組酶Sabf32的酶活力測定
酶活力測定體系:25 μL 4 mmol/L pNPAF溶液,15 μL 0.1 mol/L pH 6.0 檸檬酸緩沖液混勻,加入10 μL適當稀釋酶液。反應條件:將反應體系在40 ℃反應10 min,迅速加入150 μL 1 mol/L Na2CO3終止反應,于405 nm處測定OD值。將經過10 min沸水浴滅活的酶液做空白對照。用對硝基苯酚制作標準曲線,計算酶解反應體系中產生的對硝基苯酚含量。酶活的定義:每分鐘產生1 μmol/L的對硝基苯酚所需的酶量為一個酶活力單位。
1.2.8 重組Sabf32的酶學性質研究
酶的最適溫度及溫度穩定性:在pH 6.0的條件下,用適當稀釋的酶液分別在30、35、40、45、50、55、60、65、70、75、80、85、90 ℃下測定酶活,反應10 min,以最高酶活力為100%,計算各個溫度下的相對酶活力。溫度穩定性是將適當稀釋的酶液分別置于40、50、60 ℃下,保溫60 min,測定殘余酶活。
酶的最適pH值及pH值穩定性:分別配制pH 2.0~8.0含pNPAF的緩沖液,在最適溫度下測定不同pH值下的相對酶活,確定該酶的最適pH反應條件。pH值穩定性的測定是將濃縮后的酶液在上述不同pH條件下處理30 min后,調節至最適pH下,測定其殘余酶活。
1.2.8 比活力和動力學參數的測定
應用Lowry法測定純化重組酶Sabf32的蛋白濃度;在最適反應條件下(pH 5.0,40 ℃)測定重組酶Sabf32的活性,計算Sabf32的比酶活力。
在最適反應條件下,測定Sabf32催化不同濃度的pNPAF(1~4 mmol/L)釋放的產物量,利用米氏方程(Lineweaver-Burk)繪制曲線,求出Km值和Vmax。
2 結果與分析
2.1 發酵液中蛋白質、還原糖和酶活力測定
將裂褶菌在以玉米皮纖維為唯一碳源的培養基中發酵培養,120 h發酵培養結果顯示其生長良好,該菌株能夠將玉米皮纖維轉化為其菌體生長所必須的碳源。
通過測定發酵液中蛋白質濃度、還原糖含量和木聚糖酶酶活力,確定了裂褶菌轉錄組測序的取樣條件。其中發酵液中還原糖含量在培養48 h后達到最高(圖1),之后隨著培養時間延長,濃度逐漸降低。推測可能是在菌種生長初期開始分泌胞外降解酶,水解玉米皮纖維,培養基中還原糖含量提高,但當菌體生長加快,菌株生長加速利用培養基中還原糖,因此含量開始逐漸降低。裂褶菌表達分泌胞外酶是其降解、利用玉米皮纖維的主要途徑,當培養基上清液中還原糖含量快速上升并達到最高點時,推測其纖維降解酶也同時在進行高效地轉錄,因此選擇培養48 h為RNA提取時間。當培養至72 h時,發酵液蛋白質含量、酶活力趨于穩定,還原糖含量逐步降低,推測裂褶已經開始利用培養基中已經降解的纖維維持生長,胞外酶的轉錄效率可能開始降低。發酵液中木聚糖酶酶活力隨著培養時間延長逐漸提高(圖2),但超過72 h后,酶活力增長趨于穩定。其在48~72 h間酶活力增長較快。發酵液中蛋白質濃度隨著培養時間延長逐漸提高(圖3),但超過60 h后,蛋白質濃度增長趨于穩定。
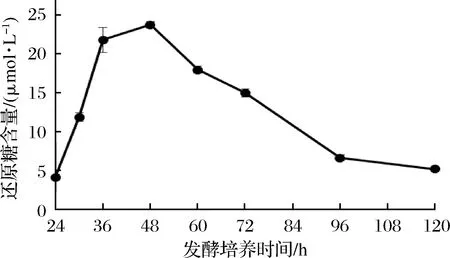
圖1 發酵液中還原糖含量Fig.1 Reducing sugar content in fermentation
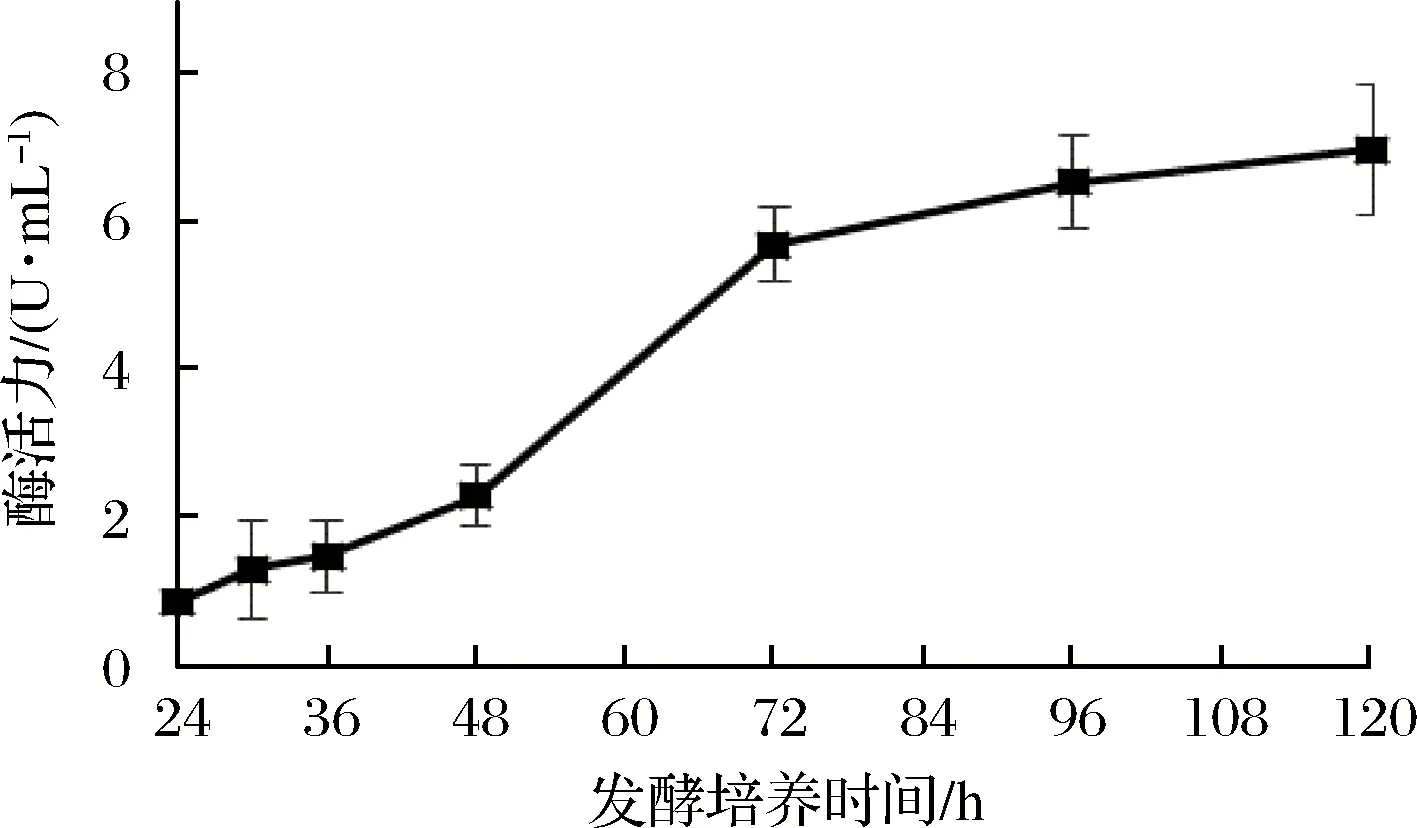
圖2 發酵液中木聚糖酶酶活力Fig.2 Activity of xylanase in fermentation
2.2 總RNA提取、測序及Unigene的組裝
提取了發酵培養48 h時的裂褶菌總RNA,經過NanoDrop定量后,確定提取的總RNA濃度為395 ng/μL,260/280比值為2.07。提取的裂褶菌RNA完整,沒有彌散(圖4),28S∶18S的比值符合構建cDNA文庫和轉錄組測序質量要求。
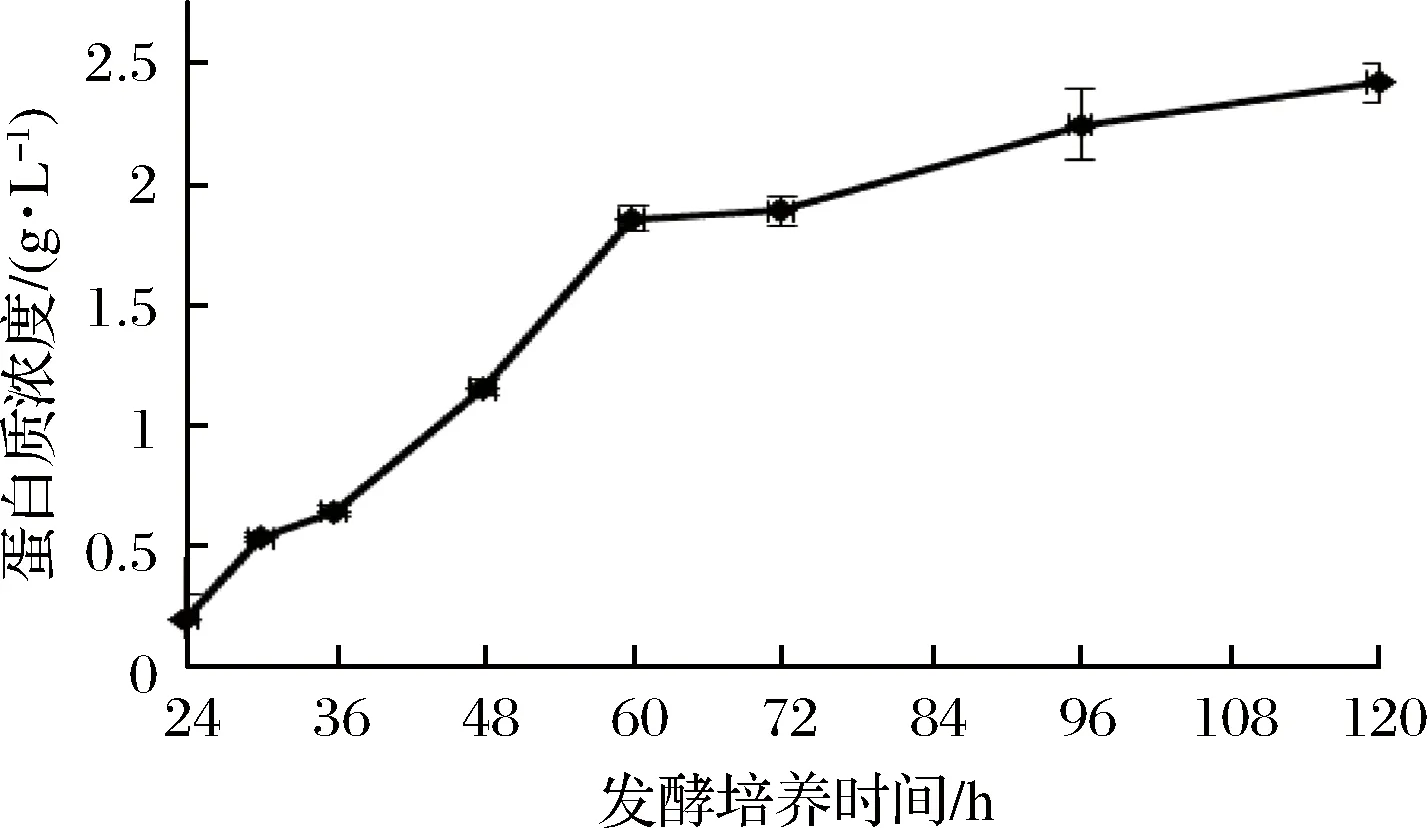
圖3 發酵液中蛋白質濃度變化Fig.3 Change of protein concentration in fermentation
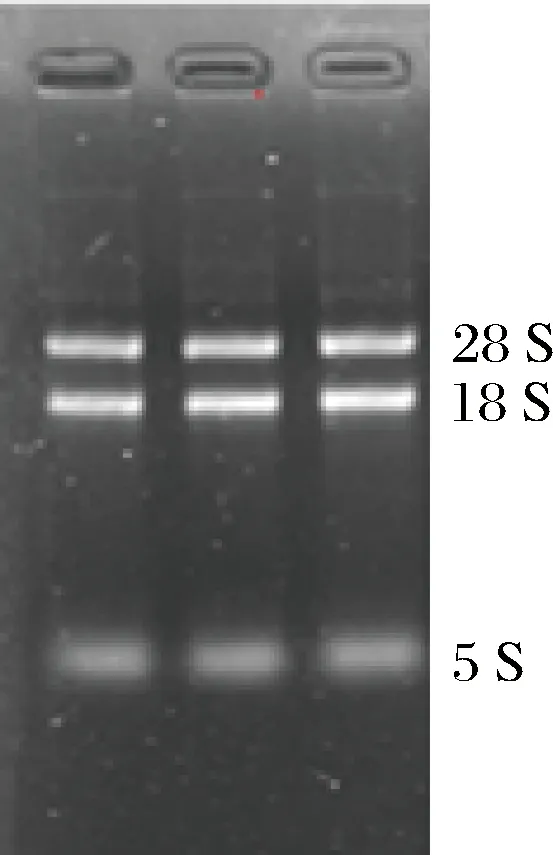
圖4 裂褶菌總RNA電泳檢測Fig.4 The electrophoresis detection of maitake RNA
轉錄組測序數據經過濾統計、拼接,組裝出全長轉錄本Unigene序列,共獲得23 656個Unigene,最長為13 092 nt,最短為201 nt,Unigene序列的平均GC含量為0.589 7。
根據Unigene序列的長度進行分析,核酸序列長度在200~400 nt之間的Unigene占比最多,而長度在200~2 000 nt之間的占85.51%,為20 229個;長度在2 000~3 000 nt之間的為2 210個,占比為9.34%;長度大于3 000 nt的Unigene數為1 217個,占總數的5.14%。
2.3 Unigene的數據庫比對分析
Unigene序列分別與NR、NT數據庫進行BLAST比對分析,分別有7 281和1 559個能夠比對成功,占總數目的30.78%和6.59%。Unigene序列分別進行BLASTP和BLASTX分析后,有7 353和7 084個能夠比對成功,占總數的31.08%和29.95%。分析顯示,共有5個作用于玉米皮半纖維素主鏈的木聚糖酶基因(表1),分別屬于糖苷水解酶第10家族和第11家族,木聚糖酶基因在687~1 113 bp之間。玉米皮半纖維素的徹底降解需要多種酶的協同作用[18],而半纖維素主鏈的降解同樣需要多種木聚糖酶的協同作用。有研究顯示,通過裂褶菌基因組測序分析能夠預測到6個木聚糖酶基因[19],其中5個為糖苷水解酶的第10家族,1個為第11家族。轉錄組測序分析預測木聚糖酶基因數量少于基因組測序分析數量,可能是由于在本研究的培養條件下,某些木聚糖基因沒有轉錄或轉錄水平低造成的。按序列同源性,木聚糖酶分別屬于糖苷水解酶第10家族、11家族和5家族,屬于同一家族的木聚糖酶催化區域具有同源性。第10家族的木聚糖酶分子量高,結構復雜,主要作用于支鏈,通常生成較小的低聚糖;第11家族主要作用于主鏈,對木聚糖有很高的特異性;第5家族只作用于有阿拉伯糖集合的位點[20-21]。

表1 木聚糖酶基因序列分析Table 1 Analysis of xylanase gene sequence
裂褶菌能夠分泌表達多種半纖維素酶,并通過多種酶的協同作用將玉米皮纖維降解利用。轉錄組結果顯示,裂褶菌還表達了多種半纖維素支鏈水解酶基因,包括木糖苷酶(β-xylosidase)、α-阿拉伯呋喃糖苷酶(α-arabinofuranosidase)、甘露糖苷酶(mannosidases)、阿魏酸酯酶(feruloyl esterase)、酯酶(esterase)等。其中與木聚糖主鏈連接的阿拉伯糖基側鏈抑制了木聚糖水解酶的活性,而α-L-阿拉伯呋喃糖苷酶能水解木聚糖的阿拉伯糖殘基,釋放阿拉伯糖分子,促進半纖維素的降解[22]。根據α-L-阿拉伯呋喃糖苷酶氨基酸序列的相似性以及活性蛋白質催化中心的結構特征,將該酶歸為糖苷水解酶的第3、43、51、54、62和127家族[23]。轉錄組測序顯示,裂褶菌能夠表達多種α-L-阿拉伯呋喃糖苷酶(表2),共有6個α-L-阿拉伯呋喃糖苷酶基因,分別屬于糖苷水解酶第43、51和62家族。目前已有研究表明α-L-阿拉伯呋喃糖苷酶可同其他半纖維素酶協同作用,促進半纖維素水解[24],因此本研究設計克隆α-L-阿拉伯呋喃糖苷酶基因Sabf32。
2.4 Unigene的注釋分類
通過GO注釋和GO功能分類分析,Unigene被注釋到生物過程(biological process)、細胞組成(cellular component)、分子功能(molecular function)3個部分,其中生物過程注釋23個條目、細胞組成注釋22個條目、分子功能注釋22個條目。
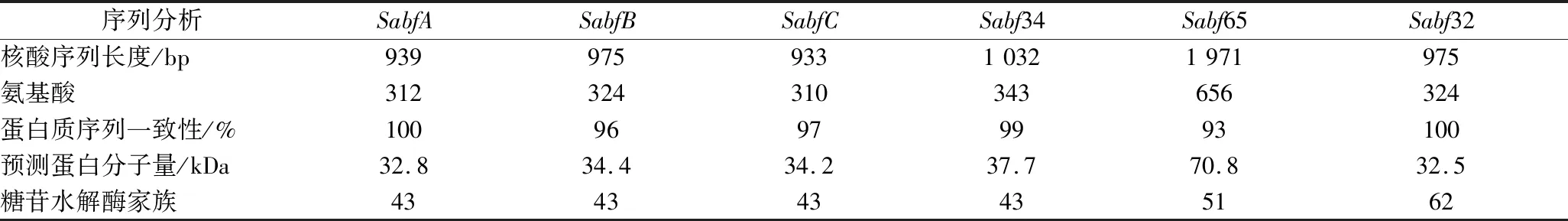
表2 α-L-阿拉伯呋喃糖苷酶基因序列分析Table 2 Analysis of α-L-arabinofuranosidase gene sequence
將Unigene的蛋白質序列提交到NCBI,獲得與Unigene編號相對應的COG編號,對COG每個類別Unigene的數目進行統計。數量最多的是一般功能預測蛋白,其他較多的分類包括與氨基酸轉運和代謝、能量產生和轉化、糖類轉運和代謝、翻譯/核糖體結構和發生、翻譯后修飾/蛋白質轉化和伴侶有關的基因,根據基因數量分布繪制裂褶菌的Unigene COG分類圖(圖5)。
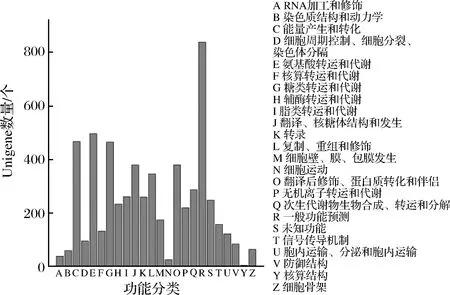
圖5 裂褶菌Unigene的COG分類Fig.5 COG functional classifications of maitake Unigene
2.5 Sabf32基因的克隆及重組表達載體的構建
以裂褶菌cDNA為模版,PCR擴增Sabf32基因的編碼序列,并進行測序分析。測序顯示Sabf32編碼序列全長975 bp,編碼324個氨基酸和1個終止密碼子;信號肽預測分析顯示,Sabf32的N端具有21個氨基酸組成的信號肽序列。通過BLASTX比對,Sabf32與來源于SchizophyllumcommuneH4-8的糖苷水解酶62家族蛋白(XP_003038505.1)具有最高一致性(100%),目前還沒有關于該基因的研究報道。
根據Sabf32序列設計合成表達引物,以cDNA為模板,PCR擴增獲得Sabf32基因片段(圖6)。
將基因片段Sabf32與表達載體pPIC9k分別進行雙酶切;將純化后的片段鏈接,構建重組表達載體pPIC9k-Sabf32。測序確定序列正確后,將重組表達載體pPIC9k-Sabf32與空載體pPIC9k分別電擊轉化畢赤酵母GS115感受態細胞,通過α-L-阿拉伯呋喃糖苷酶酶活力的測定篩選獲得陽性重組子。
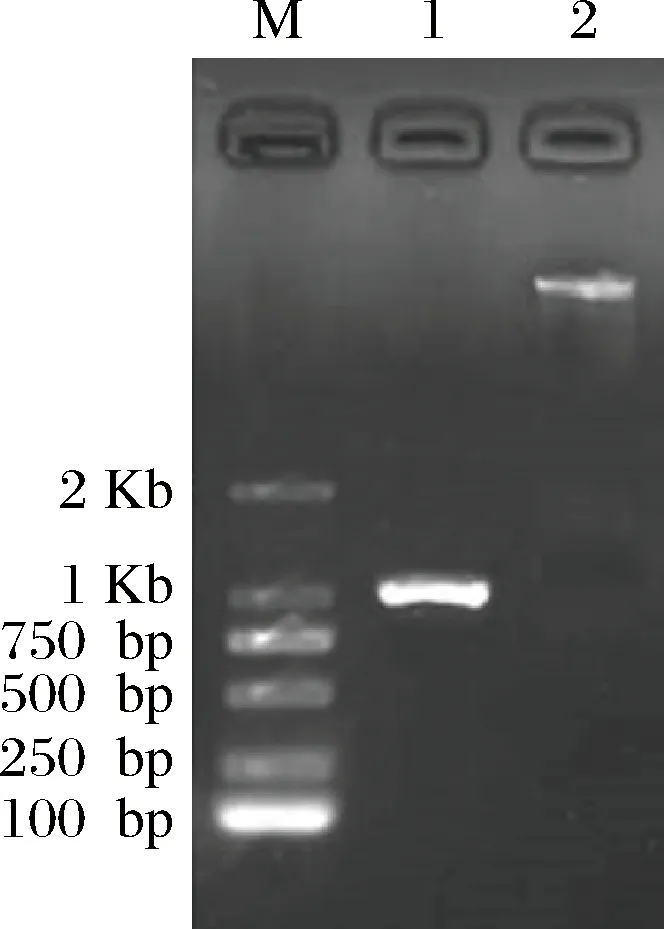
M-核酸分子量標準; 1- Sabf32;2-pPIC9k載體圖6 Sabf32和pPIC9k載體雙酶切瓊脂糖凝膠電泳檢測Fig.6 Agarose gel electrophoresis of Sabf32 and pPIC9k
2.6 重組畢赤酵母的誘導表達及酶活力測定
挑取轉化pPIC9k-Sabf32的畢赤酵母,利用BMGY發酵培養,離心收集菌體后在BMMY培養基中進行甲醇誘導表達。取發酵7 d后的發酵液的上清,進行酶活力測定和SDS-PAGE電泳檢測(圖7)。重組酶Sabf32的相對分子量為32.5 kDa。
2.7 溫度和pH對重組Sabf32活性的影響
重組Sabf32的最適反應溫度為40 ℃(圖8),在40 ℃條件下有較好的穩定性,在50 ℃條件下酶活力迅速降低,30 min后酶活完全喪失(圖9)。重組Sabf32的最適pH值為5.0,在pH 3.5~5.5的緩沖液中處理30 min,能保存80%以上的酶活(圖10)。在pH值低于2.5或高于7.0的緩沖液中處理30 min,酶活基本喪失(圖11)。
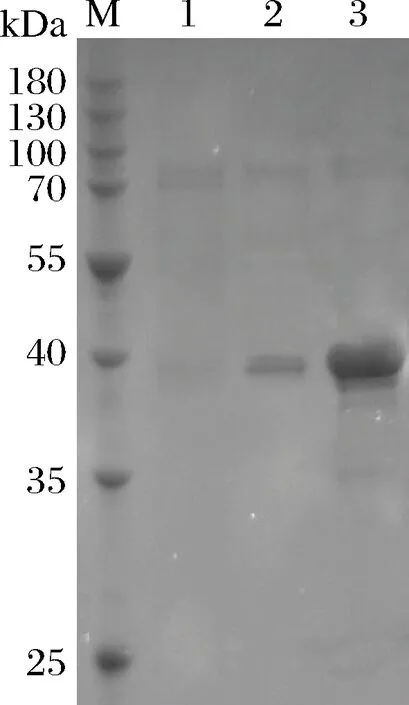
M-低分子量蛋白標準;1-對照組;2-Sabf32誘導表達發酵液;3-純化的酶液圖7 重組酶Sabf32的SDS-PAGE電泳檢測Fig.7 SDS-PAGE analysis of Sabf32
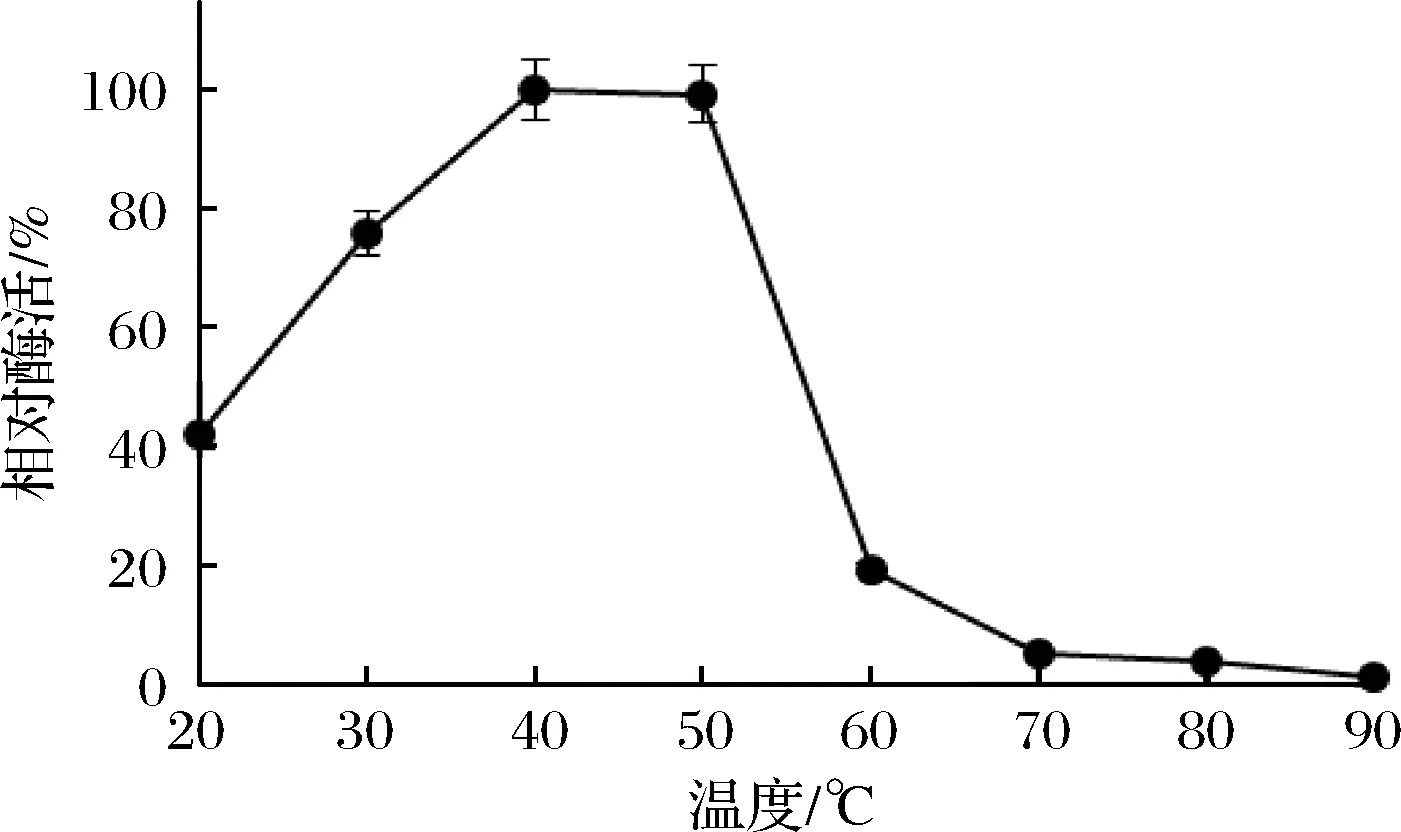
圖8 重組Sabf32的最適溫度Fig.8 The optimum temperature of Sabf32
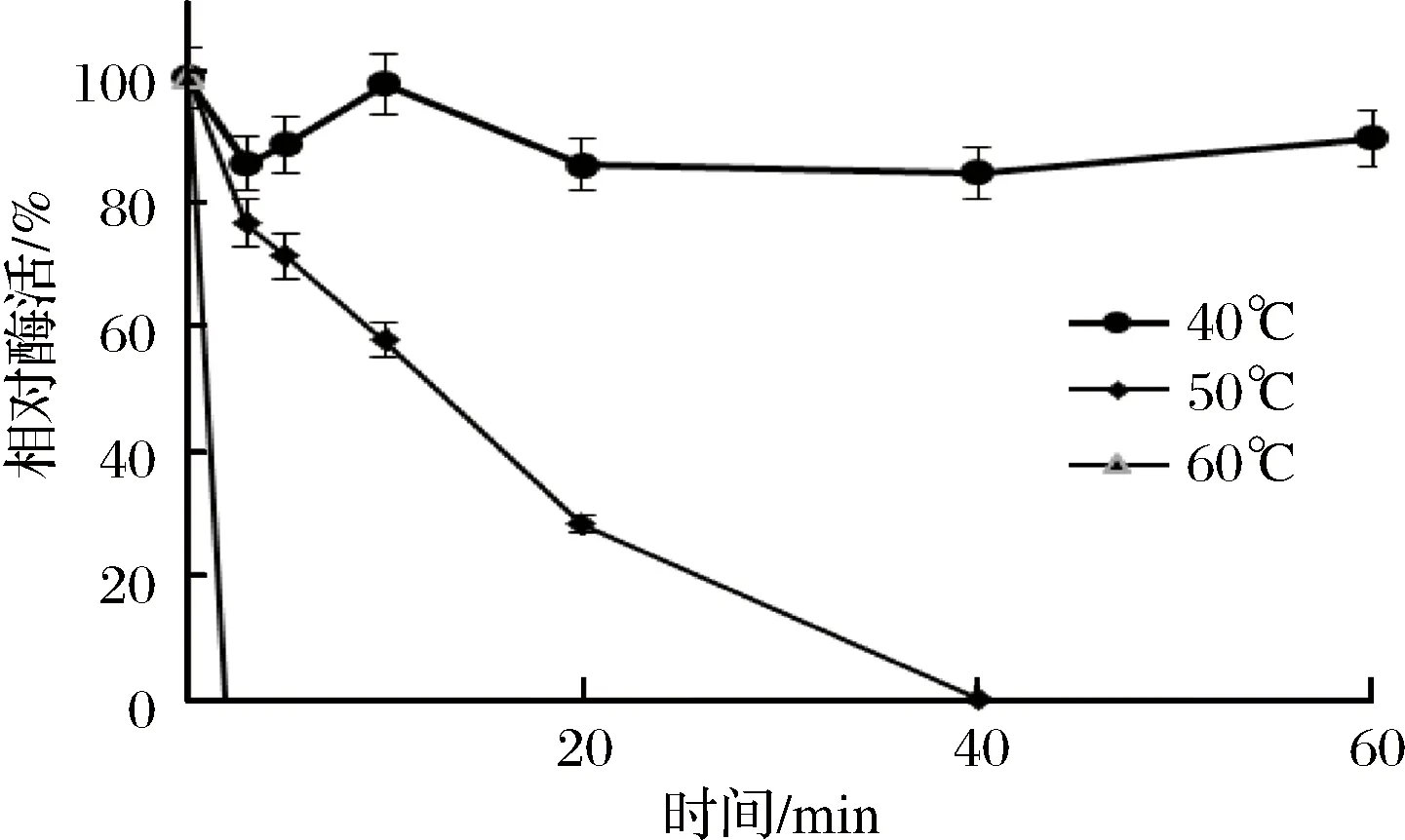
圖9 重組Sabf32的溫度穩定性Fig.9 The temperature stability of Sabf32
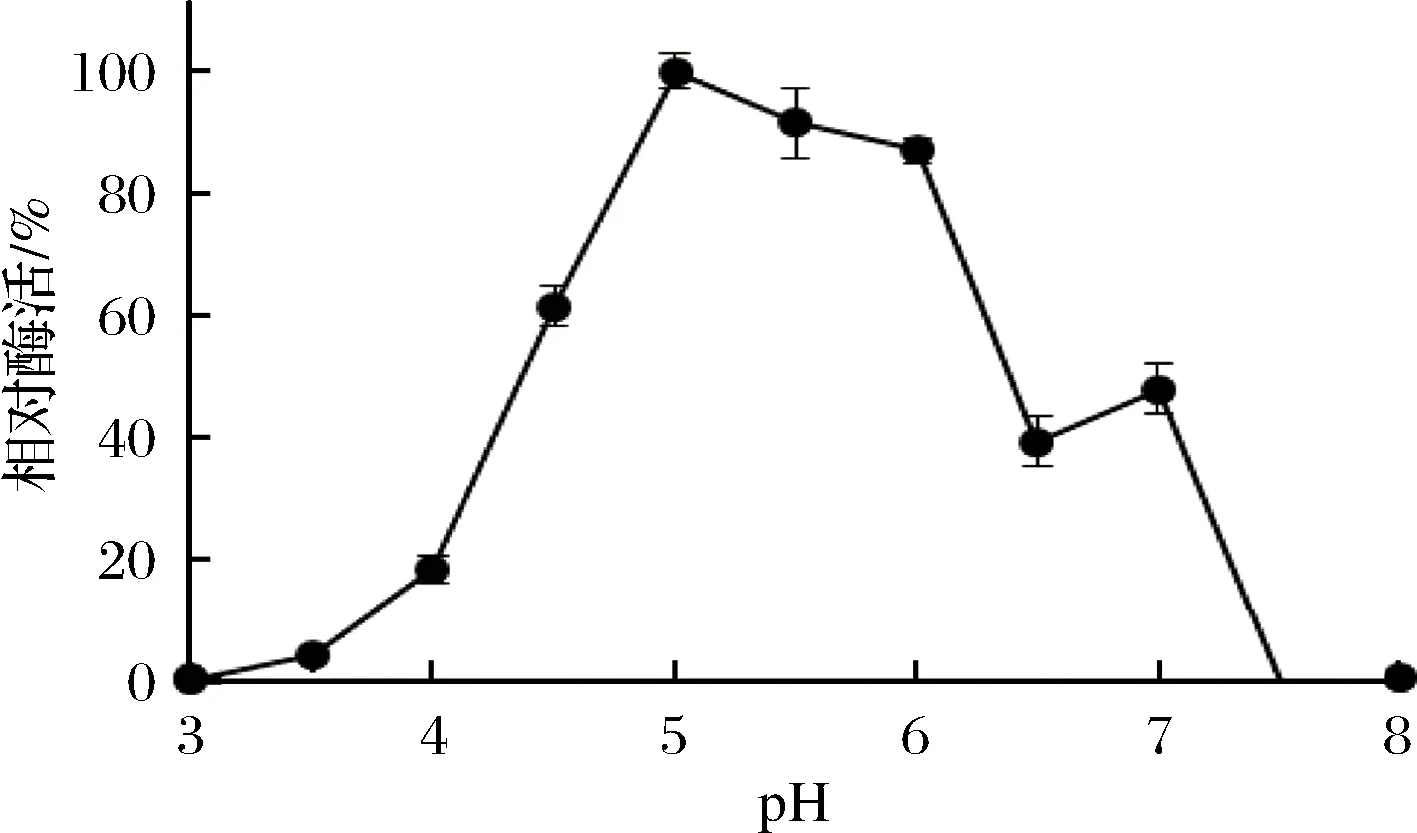
圖10 重組Sabf32的最適pHFig.10 The optimum pH of Sabf32
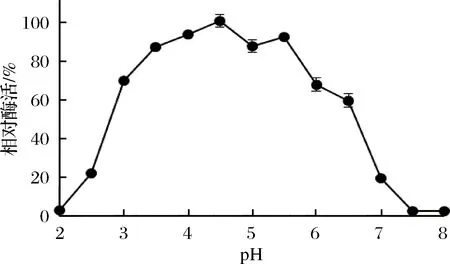
圖11 重組Sabf32的pH穩定性圖Fig.11 The pH stability of Sabf32
2.8 比活力和動力學參數的測定
經測定重組阿拉伯呋喃糖苷酶Sabf32比酶活力為16.18 U/mg,動力學常數Km和Vmax分別為(3.98±0.32) mmol/L和(2.59±0.09) μmol/(min·mg)。Sabf32比酶活力低于TalaromycesleycettanusJCM12802來源的阿拉伯呋喃糖苷酶TlAbf51,其比酶活力達到1 068 U/mg[25]。
3 結論
通過測定發酵液中蛋白質濃度、還原糖含量和木聚糖酶酶活力,確定了裂褶菌轉錄組測序的取樣條件。其中發酵液中還原糖含量在48 h達到最高,之后隨著培養時間延長逐漸降低;木聚糖酶活力隨培養時間延長而提高,在48~72h間酶活力增長較快;蛋白質濃度隨著培養時間延長逐漸提高,但超過60 h增長趨于穩定。
提取了發酵培養48 h后的裂褶菌總RNA,進行轉錄組測序,原始數據經分析后共獲得23 656個Unigene,平均GC含量為0.589 7。序列分析顯示,共有5個木聚糖酶基因,分別屬于糖苷水解酶第10家族和11家族;有6個α-L-阿拉伯呋喃糖苷酶基因,分別屬于糖苷水解酶第43、51和62家族。此外,裂褶菌還表達了多種半纖維素支鏈水解酶基因,包括木糖苷酶、甘露糖苷酶、阿魏酸酯酶、酯酶等。
克隆了α-L-阿拉伯呋喃糖苷酶基因Sabf32,編碼序列全長975 bp。Sabf32的N端具有21個氨基酸的信號肽序列,與來源于SchizophyllumcommuneH4-8的糖苷水解酶62家族蛋白(XP_003038505.1),具有最高一致性100%。完成了該酶在畢赤酵母表達,其最適溫度為40 ℃,最適pH值為4.0;在pH 3.5~5.5和40℃下較穩定。Sabf32比酶活力為16.18 U/mg,動力學常數Km和Vmax分別為(3.98±0.32) mmol/L和(2.59±0.09) μmol/(min·mg)。本研究為進一步開發玉米皮纖維降解酶以及多酶協同降解提供了基礎。
