普通煙草Aux/IAA轉(zhuǎn)錄因子家族全基因組鑒定分析
2020-01-15 00:59:46王國平周東潔牛永志鄭昀曄
中國煙草學(xué)報 2019年6期
關(guān)鍵詞:煙草
王國平,周東潔,牛永志,鄭昀曄,2*
1 玉溪中煙種子有限責(zé)任公司,玉溪市南祥路14號 653100;
2 云南省煙草農(nóng)業(yè)科學(xué)研究院,玉溪市南祥路14號 653100
生長素(auxin)是一類重要的植物激素,在細(xì)胞分裂與伸長、器官的發(fā)生與形態(tài)建成、頂端優(yōu)勢、植物向性、逆境脅迫等諸多方面都起著非常重要的調(diào)控作用[1-3],還可以與油菜素內(nèi)酯、赤霉素、乙烯等激素信號通路相互作用[4-6],影響植物種子萌發(fā)、植株生長、花果形成、植株衰老的整個生長發(fā)育過程。
生長素水平的時空動態(tài)變化可以精確而快速地激活下游一系列信號通路。其中生長素早期響應(yīng)因子在信號傳導(dǎo)中起重要作用,如生長素/吲哚-3-乙酸(Aux/IAA)家族、生長素響應(yīng)因子(ARF)家族、生長素上調(diào)小RNA(SAUR)、氨基環(huán)丙基-1-羧酸合成酶(ACS)、谷胱甘肽-S-轉(zhuǎn)移酶(GST)、生長素響應(yīng)基因GH3[7-8]。Aux/IAA是一種生長素響應(yīng)基因的轉(zhuǎn)錄抑制因子,在生長素濃度較低時,ARF-Aux/IAA異質(zhì)二聚體的形成抑制了ARF的轉(zhuǎn)錄活性;當(dāng)生長素濃度較高時,Aux/IAA發(fā)生降解,ARF從Aux/IAA中釋放,調(diào)控下游靶基因的表達(dá)[9]。典型的Aux/IAA包含四個保守的結(jié)構(gòu)域(結(jié)構(gòu)域I、II、III和IV),在許多預(yù)測的蛋白質(zhì)中可能會缺少一個或幾個結(jié)構(gòu)域[10]。結(jié)構(gòu)域I包含保守的亮氨酸重復(fù)基序LxLxLx,抑制生長素下游調(diào)控基因。結(jié)構(gòu)域II保守的GWPPV基序與F-box蛋白TIR1相互作用,導(dǎo)致Aux/IAA的不穩(wěn)定性和快速降解[11]。位于C端的結(jié)構(gòu)域III和IV能與其他Aux/IAA或ARF相互作用,形成同源二聚體和異源二聚體,從而調(diào)控下游基因表達(dá)[12]。
第一個被分離到的Aux/IAA基因是豌豆的PSIAA4/5和PS-IAA6基因[13]。隨著眾多植物基因組測序的完成,Aux/IAA基因家族已在30多種植物中得到鑒定分析[14],包括擬南芥29個[15]、油菜119個[16]、黃瓜27個[17]、大豆63個[18]、棉花44個[19]、番茄25個[20]、馬鈴薯26個[21]、玉米34個[22]、小麥84個[23]等。煙草作為模式植物,從中鑒定Aux/IAA家族基因?qū)τ谘芯可L素調(diào)控植物生長發(fā)育的機(jī)制具有重要作用。本研究利用生物信息學(xué)方法從煙草參考基因組中鑒定Aux/IAA家族基因,并對其序列特性、進(jìn)化模式、表達(dá)模式和功能注釋等進(jìn)行系統(tǒng)分析,為進(jìn)一步解析生長素調(diào)控機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1 Aux/IAA基因家族成員鑒定及序列特征分析
最新的煙草基因組數(shù)據(jù)是由Edwards K D等人于2017年發(fā)布,測序品種為K326[24]。從茄科數(shù)據(jù)庫(https://solgenomics.net/)下載該基因組數(shù)據(jù),下載的文件包括GFF文件、蛋白序列文件和核酸序列文件。從Pfam數(shù)據(jù)庫(http://pfam.xfam.org/)下載Aux/IAA基因家族的HMM模型文件(PF02309),利用HMMER3.0軟件進(jìn)行Aux/IAA蛋白序列的初步檢索,E值設(shè)為1e-5。將初步檢索得到的序列上傳至pfam網(wǎng)站進(jìn)行結(jié)構(gòu)域預(yù)測,E值設(shè)為1e-5,剔除不含Aux/IAA蛋白典型結(jié)構(gòu)域的序列,剩余的即為煙草Aux/IAA基因家族成員,同時根據(jù)Pfam結(jié)果統(tǒng)計各蛋白序列所包含的結(jié)構(gòu)域類型和個數(shù)。利用ExPaSy網(wǎng)站的ProtParam工具(http://web.expasy.org/protparam/)分析蛋白的氨基酸含量、分子量、等電點、脂肪族氨基酸指數(shù)和疏水性指數(shù)等理化性質(zhì)。
1.2 連鎖群定位和亞細(xì)胞定位
從GFF文件提取各蛋白序列的連鎖群位置信息。利用在線網(wǎng)站Softberry(http://linux1.softberry.com/)的ProtComp工具進(jìn)行亞細(xì)胞定位預(yù)測。
1.3 系統(tǒng)發(fā)育樹構(gòu)建
從Uniprot數(shù)據(jù)庫分別下載擬南芥的29個Aux/IAA家族成員[14]和番茄的25個Aux/IAA家族成員[19]。利用MEGA軟件進(jìn)行多序列比對和進(jìn)化樹構(gòu)建,采用ClustalW比對方式,構(gòu)建NJ樹,參數(shù)設(shè)置為:檢驗方法為Bootstrap法,檢驗次數(shù)設(shè)為1000次,gap處理選擇Pairwise Deletion方式。利用在線工具iTOL(https://itol.embl.de/)對進(jìn)化樹進(jìn)行可視化。
1.4 序列結(jié)構(gòu)及蛋白結(jié)構(gòu)域分析
將Aux/IAA家族成員的GFF文件提交到在線工具GSDS2.0(http://gsds.cbi.pku.edu.cn/)進(jìn)行基因結(jié)構(gòu)分析,并進(jìn)行可視化。將Aux/IAA家族成員的蛋白序列文件提交到在線工具M(jìn)EME(http://meme-suite.org/tools/meme)進(jìn)行保守結(jié)構(gòu)域的預(yù)測,參數(shù)設(shè)置為:Motif長度范圍為6~100,檢索的最大Motif數(shù)為10,其他選擇默認(rèn)參數(shù)。利用在線工具EvolView(http://www.evolgenius.info/evolview/)對蛋白結(jié)構(gòu)域進(jìn)行可視化。利用NCBI的CDD工具(https://www.ncbi.nlm.nih.gov/cdd/?term=)鑒定結(jié)構(gòu)域的類型。
1.5 表達(dá)模式分析
基因表達(dá)數(shù)據(jù)從EMBL-EBI網(wǎng)站下載,登錄號為 E-MTAB-176(http://www.ebi.ac.uk/arrayexpress/experiments/E-MTAB-176/)。煙草表達(dá)圖譜(TobEA)是由Edwards K D等人于2010年發(fā)布,該圖譜包含了早期衰老葉、中早期衰老葉、中后期衰老葉、后期衰老葉、成熟葉、莖生葉、幼葉、上部莖、下部莖、成熟根、幼根、種子、未開花蕾、開放花蕾、花朵、幼芽、莖頂端、子葉和花頂端19個樣品組織,貫穿從種子到衰老整個生命周期[25]。在SGN Unigenes數(shù)據(jù)庫,通過blastn比對檢索Aux/IAA家族成員匹配的Unigene ID,通過檢索得到的ID獲取對應(yīng)的基因表達(dá)譜數(shù)據(jù)。將數(shù)據(jù)整理后利用Cluster3.0軟件進(jìn)行聚類,利用Java TreeView工具可視化熱圖。
1.6 GO注釋分析
利用Blast2GO軟件對Aux/IAA蛋白序列進(jìn)行功能注釋,利用在線工具WEGO2.0(http://wego.genomics.org.cn/)對結(jié)果進(jìn)行可視化。
2 結(jié)果與分析
2.1 Aux/IAA家族成員鑒定及序列特征分析
基于隱馬爾科夫模型序列譜,從煙草基因組數(shù)據(jù)庫中共檢索得到84條候選基因序列,通過Pfam數(shù)據(jù)庫對候選序列進(jìn)行結(jié)構(gòu)域分析,剔除不含Aux/IAA同源異型結(jié)構(gòu)域的序列,最終得到77個煙草Aux/IAA家族基因,按照NtIAA1~NtIAA77的順序?qū)γ總€基因依次進(jìn)行編號(見表1)。通過Pfam數(shù)據(jù)庫進(jìn)行結(jié)構(gòu)域分析結(jié)果顯示,在77個基因中,只包含Aux/IAA結(jié)構(gòu)域的有56個基因;同時包含Aux/IAA和ARF結(jié)構(gòu)域的有21個基因,這21個基因同樣也屬于ARF家族;同時包含Aux/IAA、ARF和B3結(jié)構(gòu)域的有15個基因,這15個基因同樣也屬于B3超家族;分析Aux/IAA家族成員的理化性質(zhì)發(fā)現(xiàn),平均氨基酸長度為379,范圍在72~1104之間;平均分子量為42191.17 kDa,范圍在 8119.66~121828.27 之間;平均等電點為6.74,范圍在4.31~9.65之間,其中酸性蛋白(等電點小于7)有51個,堿性蛋白(等電點大于7)有26個;平均脂肪族氨基酸指數(shù)為71.95,范圍在59.5~92.08之間;平均疏水性指數(shù)為-0.559,范圍在-0.106~-1.244之間,該家族蛋白全部表現(xiàn)為親水性。
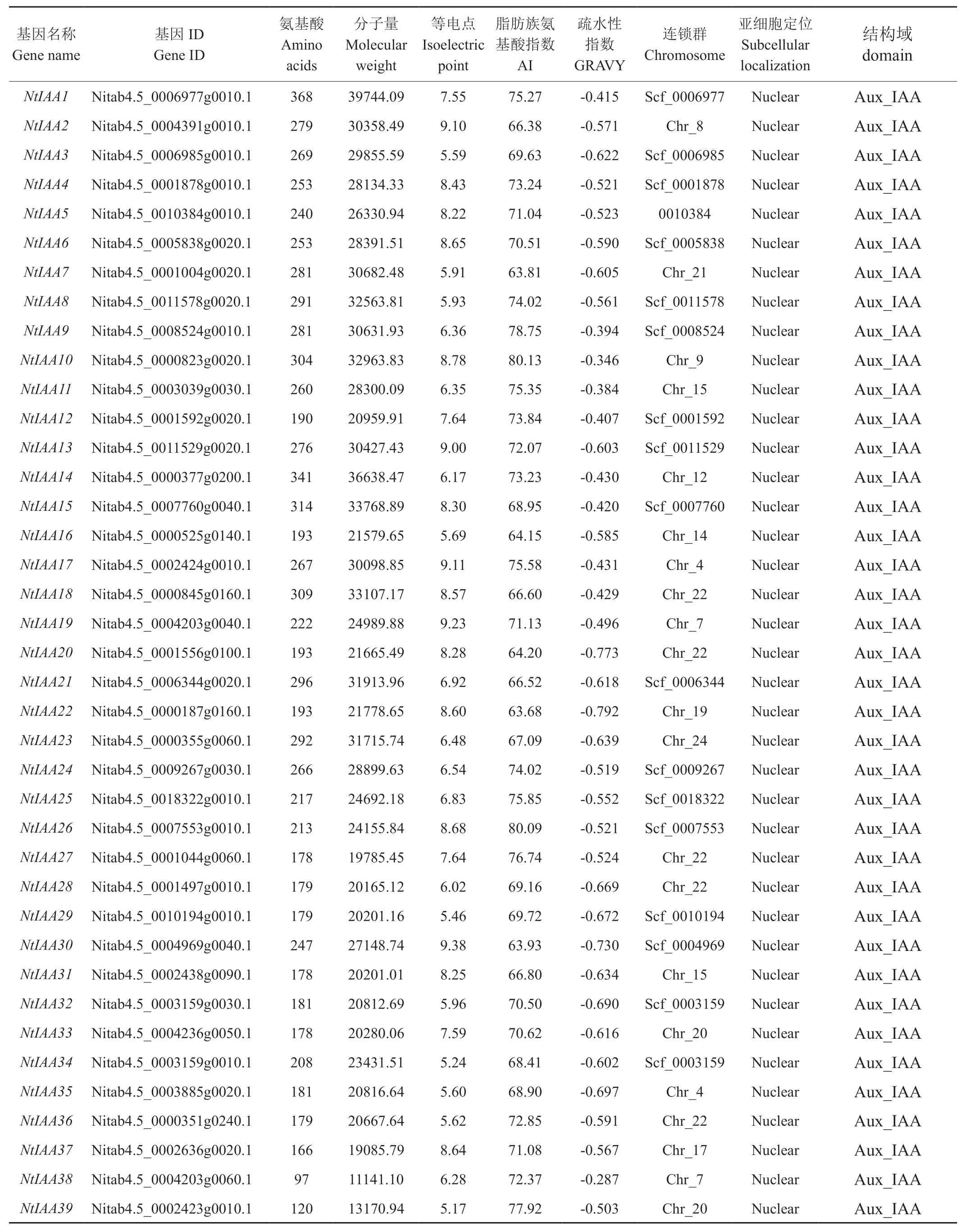
表1 Aux/IAA 家族蛋白特征Tab.1 Protein characteristics of Aux/IAA gene family

續(xù)表1
2.2 Aux/IAA家族連鎖群定位和亞細(xì)胞定位分析
連鎖群定位結(jié)果顯示(見表1),其中有46個基因定位到連鎖群上,有31個基因位于Scafford上。在定位到連鎖群上的46個Aux/IAA家族基因中,除了連鎖群2、3、6、11和16號外,其他連鎖群上均有分布,其中22號連鎖群上分布最多,達(dá)6個基因,9、17、19和20號連鎖群上各分布有4個基因,4和15號連鎖群各分布有3個基因,7、8、12、14、18、23號連鎖群上各分布有2個基因,1、5、10、21和24號連鎖群各分布有1個基因。在定位到Scafford上的31個Aux/IAA家族基因中,其中Scf_0007553和Scf_0003159上各分布有2個基因,其余Scafford上各分布有1個基因。亞細(xì)胞定位分析結(jié)果顯示(見表1),Aux/IAA家族所有基因都定位于細(xì)胞核內(nèi),這與以往研究結(jié)果一致[26]。
2.3 Aux/IAA家族進(jìn)化分析
選取模式植物擬南芥的Aux/IAA基因29個,選取茄科作物番茄的Aux/IAA基因25個,同本研究獲得的77個煙草Aux/IAA基因一同構(gòu)建系統(tǒng)進(jìn)化樹。參照以往研究結(jié)果[27-28],將Aux/IAA家族分為A和B兩個類群(圖1),A類群又分為4個亞群(A1、A2、A3和A4),B類群又分為6個亞群(B1、B2、B3、B4、B5和B6)。A類群包括65個基因(煙草35個,擬南芥14個,番茄15個),B類群包括66個基因(煙草42個,擬南芥15個,番茄10個)。除了B6外,每個亞群中都分布有擬南芥、番茄和煙草的Aux/IAA基因,說明Aux/IAA分化時間早于物種分化時間。整個系統(tǒng)發(fā)育樹中共包括46對姐妹對,其中大部分為旁系同源蛋白,有40對(煙草30對,擬南芥9對,番茄1對),直系同源蛋白有6對(5對煙草-番茄,1對擬南芥-番茄),這可能是因為普通煙草是由絨毛煙草和林煙草經(jīng)過染色體加倍進(jìn)化而來,因此存在大量相似序列。另外,通過分枝的聚集也可以看出,煙草Aux/IAA基因與番茄更為接近,這是因為煙草和番茄同屬于茄科作物,親緣關(guān)系更近。令人驚奇的是,在B6亞群中,21個煙草Aux/IAA基因聚到了一起,而且與其他基因距離較遠(yuǎn),進(jìn)一步對序列結(jié)構(gòu)分析發(fā)現(xiàn),這是因為這21個基因中,除了包含Aux/IAA結(jié)構(gòu)域,還包含了ARF結(jié)構(gòu)域,其中15個基因還包含B3結(jié)構(gòu)域,這一點與其他作物差異較大。

圖1 Aux/IAA 家族系統(tǒng)進(jìn)化樹Fig.1 Phylogenetic tree of Aux/IAA gene family
2.4 Aux/IAA家族序列結(jié)構(gòu)及蛋白結(jié)構(gòu)域分析
典型的Aux/IAA基因包含四個結(jié)構(gòu)域(Ⅰ、Ⅱ、Ⅲ和Ⅳ)。本研究結(jié)構(gòu)域分析結(jié)果顯示(圖2),所有煙草Aux/IAA基因都缺失了結(jié)構(gòu)域Ⅰ,有40個基因同時包含了結(jié)構(gòu)域Ⅱ(Motif3)、Ⅲ(Motif2)和Ⅳ(Motif1和Motif6),但其中有10個基因的結(jié)構(gòu)域Ⅳ缺失了一部分(Motif6);有27個基因只包含結(jié)構(gòu)域Ⅲ和Ⅳ,但其中有18個基因的結(jié)構(gòu)域Ⅳ缺失了一部分(Motif6),這27個基因中的大部分屬于B6亞群成員(21個);基因NtIAA24、NtIAA30和NtIAA40只包含結(jié)構(gòu)域Ⅱ和Ⅳ,但NtIAA40的結(jié)構(gòu)域Ⅳ缺失了一部分(Motif6);基因NtIAA37和NtIAA41只包含結(jié)構(gòu)域Ⅱ和Ⅲ;基因NtIAA53、NtIAA52、NtIAA45和NtIAA48分別只包含結(jié)構(gòu)域Ⅱ、Ⅲ、Ⅳ;基因NtIAA74沒有檢測到任何結(jié)構(gòu)域。綜合來看,大部分煙草Aux/IAA家族基因(有68個)都包含了結(jié)構(gòu)域Ⅲ和Ⅳ,結(jié)構(gòu)域Ⅲ和Ⅳ可與ARF相互作用,形成同源或異源二聚體,從而抑制生長素響應(yīng)基因的表達(dá),在功能行使中發(fā)揮重要作用。很明顯,B6亞群基因(21個)同時也包含了B3結(jié)構(gòu)域(Motif4和Motif5)、ARF結(jié)構(gòu)域(Motif7)以及類型未知的Motif8、Motif9和Motif10,B3基因家族是一個超基因家族,ARF基因家族也是一個生長素早期響應(yīng)因子,Aux/IAA、B3和ARF家族成員之間通過互作,將眾多的信號通路聯(lián)系在一起。Aux/IAA內(nèi)含子-外顯子結(jié)構(gòu)分析顯示,除了B6亞群基因序列較長、外顯子數(shù)目較多外,其余基因序列相對較短,外顯子數(shù)目較少,結(jié)構(gòu)較為簡單。
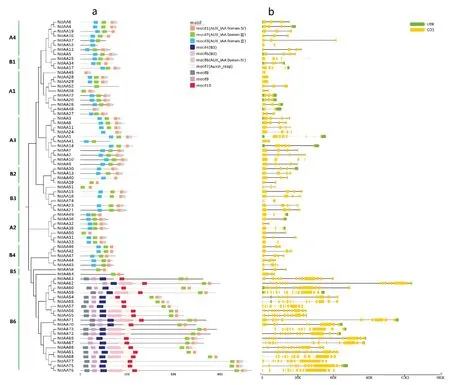
圖2 Aux/IAA家族蛋白結(jié)構(gòu)域與序列結(jié)構(gòu)Fig.2 Protein domain and sequence structure of Aux/IAA gene family
2.5 Aux/IAA家族表達(dá)模式分析
煙草Aux/IAA家族的77個成員中有50個基因,在煙草表達(dá)圖譜(TobEA)檢索到表達(dá)數(shù)據(jù)。基因聚類結(jié)果將這50個基因分為X和Y兩個類群,其中X類群又分為X1和X2兩個亞群,Y類群又分為Y1和Y2兩個亞群,X類群基因與系統(tǒng)發(fā)育樹中的A類群基因基本相同,Y類群基因與系統(tǒng)發(fā)育樹中的B類群基因基本相同,進(jìn)一步說明同源性較高的基因之間有著相似的表達(dá)模式和生物學(xué)功能(圖3)。組織聚類結(jié)果將這19個組織分為兩個類群,其中一個類群包含根、莖、葉和種子等成熟的組織,另一個類群包含花蕾、子葉、幼葉、莖端等幼嫩的組織。
整體上看,不同亞群基因在不同的組織中的表達(dá)水平有所差異,同一亞群基因在不同的組織中的表達(dá)水平類似。X1亞群基因在不同發(fā)育時期的葉片中的表達(dá)量較高,X2亞群基因在花尖、幼芽、幼葉、未開放花蕾和開放花蕾等幼嫩組織中的表達(dá)量較高,Y1亞群基因在花朵和種子中的表達(dá)量較高,Y2亞群基因在根、莖和種子中的表達(dá)量較高。雖然不同Aux/IAA家族基因表達(dá)存在一定的組織特異性,但在植株生長發(fā)育的每個階段,都有Aux/IAA家族基因的參與。
2.6 Aux/IAA家族GO注釋分析
GO注釋結(jié)果顯示(圖4),Aux/IAA家族基因在細(xì)胞組成、分子功能和生物進(jìn)程中都發(fā)揮了重要作用。在細(xì)胞組成方面,主要參與細(xì)胞、胞內(nèi)和細(xì)胞器的組成,這關(guān)系到生長素可以直接作用于細(xì)胞膜或胞內(nèi)組分,協(xié)同調(diào)節(jié)分裂、伸長和分化等胞內(nèi)過程,而且作為非細(xì)胞自主信號,與其他信號轉(zhuǎn)導(dǎo)途徑相互作用,從而影響一些細(xì)胞反應(yīng);在分子功能方面,主要參與蛋白結(jié)合,這間接調(diào)控下游基因的表達(dá),從而影響生長素對植物生長發(fā)育的調(diào)節(jié);在生物進(jìn)程方面,主要參與了胞內(nèi)、代謝等生物進(jìn)程調(diào)節(jié),信號轉(zhuǎn)導(dǎo),內(nèi)源、胞內(nèi)、化學(xué)等刺激反應(yīng),氮、細(xì)胞代謝過程等功能,這關(guān)系到生長素對植物向性生長、組織分化、器官發(fā)生、頂端優(yōu)勢等的調(diào)節(jié)作用。
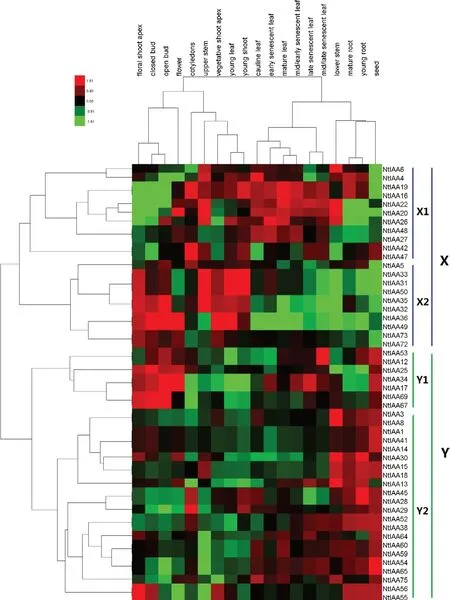
圖3 Aux/IAA家族基因表達(dá)熱圖Fig.3 Heatmap of gene expression of Aux/IAA gene family
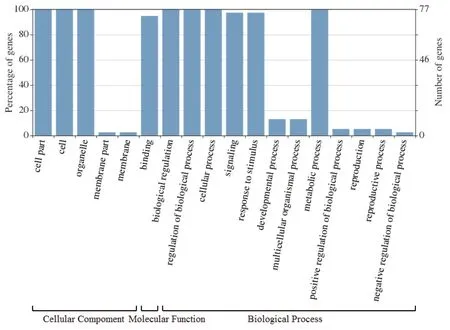
圖4 Aux/IAA家族GO注釋結(jié)果Fig.4 GO annotation results of Aux/IAA gene family
3 討論
隨著越來越多的植物基因組測序的完成,全基因組基因鑒定為基因功能研究提供了一種更高效的方法。Aux/IAA家族基因已在許多植物中得到鑒定[13],以往研究表明,很大一部分Aux/IAA家族基因是由基因復(fù)制事件(片段復(fù)制、串聯(lián)復(fù)制)進(jìn)化產(chǎn)生的,而且Aux/IAA家族基因的復(fù)制事件似乎與全基因組復(fù)制(WGD)相一致。例如:大豆發(fā)生了兩次全基因組復(fù)制事件和一次全基因組三倍化事件,其中90%的Aux/IAA基因以片段復(fù)制的形式產(chǎn)生[18]。本研究共得77個普通煙草Aux/IAA家族基因,在已鑒定的30多種植物中,數(shù)量僅次于甘藍(lán)型油菜的119個[16]和普通小麥的84個[23]。而普通煙草是異源四倍體,普通小麥?zhǔn)钱愒戳扼w,甘藍(lán)型春油菜是由自然種間雜交后雙二倍化進(jìn)化而來的一種復(fù)合種,這三種作物都發(fā)生了基因擴(kuò)張,可能是在進(jìn)化過程中發(fā)生了全基因組復(fù)制使連鎖群加倍導(dǎo)致的,Aux/IAA基因擴(kuò)張產(chǎn)生的功能冗余可能是為了適應(yīng)復(fù)雜的自然環(huán)境。
Aux/IAA基因一般包括四個典型的結(jié)構(gòu)域(I、II、III和IV),但在大部分植物中都存在一部分Aux/IAA基因缺失了其中某個或數(shù)個結(jié)構(gòu)域[10]。例如:擬南芥中有11個(占比38%)[15],木瓜中有8個(占比44%)[29],馬鈴薯中有5個(占比15%)[21]等。在普通煙草中檢測到的77個Aux/IAA基因中,全部缺失了結(jié)構(gòu)域I,有一半多(40個)包含了其余所有結(jié)構(gòu)域,大部分(68個)都包含了結(jié)構(gòu)域Ⅲ和Ⅳ。這些結(jié)構(gòu)域的多樣性可能在生長素信號通路中行使不同的功能以適應(yīng)復(fù)雜多變的自然環(huán)境。有趣的是,在B6亞群中的21個基因不僅含有Aux/IAA的結(jié)構(gòu)域III和IV,也包含了ARF結(jié)構(gòu)域,其中15個基因還包含了B3結(jié)構(gòu)域。這些基因也屬于ARF基因家族或B3超基因家族。B3是一個廣泛存在于高等植物基因組中,并能與DNA結(jié)合的高度保守結(jié)構(gòu)域,ARF基因家族也是一個生長素早期響應(yīng)因子。以往研究表明,ARF基因的C端包含一個CTD結(jié)構(gòu)域,該結(jié)構(gòu)域與Aux/IAA的結(jié)構(gòu)域III和IV高度相似。因此,Aux/IAA的結(jié)構(gòu)域III和IV可以與ARF的CTD結(jié)構(gòu)域相互作用,從而調(diào)控ARF蛋白的活性[30-31]。
一般認(rèn)為普通煙草是由S基因組供體林煙草和T基因組供體絨毛煙草兩個祖先種,大約在20萬年前經(jīng)過一次種間雜交產(chǎn)生[32],其存在兩套異源基因組。因此,本研究進(jìn)化分析顯示,在鑒定的77個普通煙草Aux/IAA家族中存在大量的旁系同源基因。通過系統(tǒng)發(fā)育樹可以看出,煙草與番茄親緣關(guān)系更近,這是因為他們同屬于雙子葉綱茄科植物。以往研究表明,SiIAA3連接生長素和乙烯代謝途徑,RNA干擾該基因會導(dǎo)致幼苗葉片下垂和葉尖卷曲[33];RNA干擾SiIAA9該基因會導(dǎo)致葉片不規(guī)則和單性果實[34];抑制SiIAA15的活性會導(dǎo)致側(cè)根增多、腋芽異常發(fā)育、絨毛數(shù)量減少、坐果率降低、葉片厚度增大[35]。SiIAA17控制果實的重量[36];SiIAA27與葉綠素合成和獨腳金內(nèi)酯合成有關(guān)[37-38]。SiIAA3與煙草的NtIAA20、NtIAA22、NtIAA27和NtIAA48聚到一起;SiIAA9與煙草的NtIAA1、NtIAA14和NtIAA41聚到一起;SiIAA15與煙草的NtIAA17和NtIAA34聚到一起;SiIAA17與煙草的NtIAA37和NtIAA53聚到一起;SiIAA27與煙草的NtIAA3和NtIAA8聚到一起。這些同源基因可能也具有類似的生物學(xué)功能。
Aux/IAA蛋白可以與ARF蛋白相互作用形成二聚體,從而影響生長素的調(diào)控作用。Aux/IAA蛋白是一種核蛋白,這與本研究的結(jié)果一致。理論上講ARF蛋白也屬于核蛋白,但是最新的研究發(fā)現(xiàn)大部分ARF蛋白被定位到細(xì)胞核,個別ARF蛋白卻被定位到葉綠體或線粒體[39]。這可能是由于ARF除了可以與Aux/IAA結(jié)合發(fā)揮作用,還有其他結(jié)合方式,例如:可以與含有TGTCTC AuxREs元件的生長素響應(yīng)基因結(jié)合,可以通過它們的CTDs形成二聚體,再與靶基因結(jié)合等[40]。
Aux/IAA參與了胚胎發(fā)育、下胚軸生長、向性、花器官發(fā)育、側(cè)根伸長等生物過程,大部分?jǐn)M南芥基因突變都會表現(xiàn)出相似的功能缺陷[41-42]。本研究中雖然不同Aux/IAA家族基因表達(dá)存在一定的組織特異性,但在植株生長發(fā)育的每個階段,都有Aux/IAA家族基因的參與。另外,Aux/IAA在植物抗逆性方面也具有很大作用。高粱在鹽和干旱脅迫下,SbIAA1受到誘導(dǎo),其表達(dá)量在葉和根部組織中不同,該基因可能對高粱抗鹽抗干旱有重要作用[43]。本研究所分析的77種Aux/IAA蛋白均為親水性蛋白,蛋白質(zhì)的高親水性使蛋白質(zhì)能夠代替細(xì)胞中的水分子,從而很好的抵御干旱脅迫對植物造成的損傷[44]。
在過去十年里,在生長素感知和信號轉(zhuǎn)導(dǎo)機(jī)制方面取得了顯著成就。但在生長素信號通路中,Aux/IAA的介導(dǎo)途徑仍存在很多空白,Aux/IAA在植物抗逆方面的相關(guān)研究也相對較少。相信隨著基因組的不斷解析以及計算模型的不斷發(fā)展,越來越多的Aux/IAA信號網(wǎng)絡(luò)將被發(fā)現(xiàn)。煙草作為一種模式植物,通過研究煙草Aux/IAA基因,對于解析Aux/IAA功能及信號通路具有重要意義。
4 結(jié)論
本研究從普通煙草基因組中共鑒定了77個Aux/IAA家族基因,均定位于細(xì)胞核內(nèi),全部為親水性蛋白,大部分為酸性蛋白。Aux/IAA家族劃分為2個類群,10個亞群,Aux/IAA基因分化時間早于物種分化時間;煙草是異源四倍體,Aux/IAA家族存在大量旁系同源姐妹對(40對)。所有的煙草Aux/IAA基因均缺失了結(jié)構(gòu)域Ⅰ,大部分基因都同時包含了結(jié)構(gòu)域Ⅲ和Ⅳ;B6亞組基因同時包含了B3、ARF和Aux/IAA結(jié)構(gòu)域,與其他基因差別較大。不同亞群的基因組織表達(dá)特異性不同,同一亞群的基因組織表達(dá)特異性一致。Aux/IAA家族基因主要參與細(xì)胞組成、蛋白結(jié)合、生物進(jìn)程調(diào)節(jié)、信號轉(zhuǎn)導(dǎo)、刺激反應(yīng)、代謝過程等功能。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學(xué)報(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學(xué)林(2013年2期)2013-11-12 12:58:54
中國煙草學(xué)報(2012年5期)2012-04-12 06:21:18
