土壤氮素異質性分布和馬先蒿寄生對長芒棒頭草生長發育及根系分布的影響
2020-01-18 02:24:00田玉清隋曉琳張婷李艷梅李愛榮
廣西植物 2020年12期
田玉清 隋曉琳 張婷 李艷梅 李愛榮


摘?要:無論在農田還是自然生態系統中,土壤養分異質性普遍存在。植物具有感知土壤養分異質性的能力,并通過調節根系生物量分配及空間分布以獲取更多資源。了解寄生脅迫在不同養分條件下對寄主生長發育及根系空間分布的影響,對解析寄主應對寄生脅迫和養分脅迫的適應策略,進而指導寄生性雜草防控具有重要的指導意義。該文采用分根試驗,通過對寄主分根,并控制根室兩側氮供應水平及寄生脅迫程度,考察了氮脅迫及兩種寄主依賴程度不同的馬先蒿的寄生對寄主長芒棒頭草生長發育及根系空間分布的影響。結果表明:(1)土壤氮水平與馬先蒿寄生均可顯著影響長芒棒頭草生物量及根冠比,并且兩者之間存在顯著交互作用,其中土壤氮水平為主要影響因子。(2)兩種馬先蒿對長芒棒頭草的危害程度不同。在NPK和2NPK 處理時,三色馬先蒿的寄生顯著降低長芒棒頭草生物量(莖葉:37.1%、51.5%;根系:35.6%、63.6%);在NPK處理時,大王馬先蒿的寄生顯著增加長芒棒頭草生物量(莖葉:29.9%,根系:61.2%)。(3)長芒棒頭草的根系生長和空間分布受氮營養的異質分布和寄生的影響,具有明顯的感知養分空間分布及調節根系生長能力。
關鍵詞:土壤氮水平, 馬先蒿, 根部半寄生植物, 根系空間分布, 分根盆栽試驗
中圖分類號:Q945
文獻標識碼:A
文章編號:1000-3142(2020)12-1838-11
Abstract:Soil nutrient heterogeneity is ubiquitous in both farmlands and natural ecosystems.Plants have the ability to perceive soil nutrient heterogeneity and adjust biomass allocation to optimize root spatial distribution so as to obtain more resources.As an important nutrient stress, the effects of a root hemiparasite on spatial distribution of its host roots remain untested.Understanding parasitism effects on host growth and root distribution under different nutrient conditions is of great significance for a better knowledge of host adaptation strategies, which will contribute to parasitic weed management.A split-root pot cultivation experiment was conducted to investigate the effects of nitrogen supply levels and parasitism by two root hemiparasitic Pedicularis species on host (Polypogon monspeliensis) growth and biomass allocation to each side of the split roots.The results were as follows:(1) Nitrogen levels and parasitism by Pedicularis species had significant effects on host biomass and root allocation patterns.Significant interaction effects were detected between nitrogen levels and parasitism, with nitrogen levels contributing more to the observed variances.(2) P.tricolor and P.rex showed different effects on host biomass.P.tricolor reduced host shoot biomass by 37.1% and 51.5%, and root biomass by 35.6% and 63.6%, respectively, when supplied with NPK and 2NPK solutions.However, P.rex significantly increased host shoot biomass by 29.9% and root biomass by 61.2% in NPK treatment and had little effect in 2NPK treatments.(3) Polypogon monspeliensis roots had obvious perception of nitrogen heterogeneity and parasitism by the root hemiparasites, as shown by foraging behavior for nitrogen and a tendency to avoid Pedicularis roots in treatments with equally low N levels.
Key words:soil nitrogen levels, Pedicularis, root hemiparasitic plants, root spatial distribution, split-root pot cultivation
無論在農田還是自然生態系統中,由于受多種生物和非生物過程的影響,養分和水分等在土壤中常呈不均衡分布,導致土壤環境具有明顯的異質性(Jackson & Caldwell, 1993)。植物具有感知土壤環境異質性,并相應調節植株養分和生物量分配以獲取更多資源的能力 (Benning & Seastedt, 1997; Zhang & Forde 1998; Li et al., 2012b)。如植物可以將更多的根系分布到養分富集區域,以提高養分吸收效率(Wijesinghe et al., 2001; Day et al., 2003; 李洪波,2014)。此外,植物之間的種內競爭、種間競爭以及食草動物的取食等均會影響植物根系空間分布與生長狀態( 王煒等,2000;邵玉琴等,2006; Schiffers et al., 2011)。了解植株生物量分配和根系空間分布變化對生物和非生物脅迫的響應,對認識植物應對不同脅迫因子的響應策略,進而探討物種間的相互作用和生物多樣性的維持機制至關重要。
根部半寄生植物(root hemiparasitic plants)是寄生植物中的一大類群,它們具有一定的光合能力,但通常根系發育不健全,從土壤中獲取養分的能力較差,需要通過吸器(haustorium)從寄主根部獲取養分和水分等供自身生長與繁殖(Press, 1989; Stewart & Press, 1990; Shamoun, 2009)。根部半寄生植物大量掠奪寄主養分和水分,常顯著降低寄主生物量和品質(Press, 1989; Parker & Riches, 1993)。因此,很多根部半寄生植物已成為農田或草地生態系統中的惡性雜草,給農業和畜牧業生產造成嚴重損失(Press, 1989; Parker & Riches, 1993)。如獨腳金屬(Striga Lour.)植物可以寄生于高粱、玉米等農作物,導致寄主產量下降6%~21%(Frost et al., 1997),甚至絕收(Hearne, 2009 ; Smaling et al., 1991),給非洲農業生產造成巨大損失。在我國,馬先蒿屬(Pedicularis L.)根部半寄生植物在亞高山草甸廣泛分布,對禾草類寄主生長的抑制作用尤為明顯,大面積蔓延的種類會嚴重危害當地畜牧業發展(王文香等,2009;隋曉琳,2013)。
由于根部半寄生植物分布廣泛且寄主范圍較廣,是多數生態系統中常見的一種生物脅迫因素(Press 1989; Parker & Riches, 1993)。之前對根部半寄生植物與寄主植物互作的研究多關注地上部分的寄生危害,對寄主植物如何調節根系空間分布來應對根部半寄生植物的寄生脅迫研究較少。有報道表明,在遭遇根部半寄生植物的寄生脅迫時,寄主對根系的投資比例有明顯變化(Li et al., 2012a),但因先前研究中寄主根系被侵染和未被侵染的區域對馬先蒿的反應還未知,寄主被寄生后對根系投資比例的變化是一種主動適應過程還是被動脅迫結果仍是未解之謎。
對于大部分草地生態系統來說,氮素是限制其生產力的重要營養元素之一,同時也是調節草地生態系統結構和功能的一種關鍵性元素(Liu et al., 2017)。土壤氮素水平可顯著影響根部半寄生植物與禾草類寄主間的互作關系。如Aflakpui et al.(2002)發現獨腳金(Striga hermonthica)在土壤氮素水平較低時對玉米的危害更嚴重。Gibson & Watkinson(1991)發現在土壤氮素水平較低時,小佛甲草(Rhinanthus minor)對寄主的危害更大;而在土壤氮素水平較高時,其對寄主生物量的抑制程度相對降低。近期的研究顯示,提高草地生態系統氮素水平,不僅可以促進禾本科植物的生長,而且可以抑制根部半寄生植物甘肅馬先蒿的生長(Liu et al., 2017)。然而,氮素水平的變化如何通過影響根部半寄生植物和寄主植物地下部分的互作而使寄生雜草的危害得以緩解還尚未可知。
鑒于寄生植物對寄主植物養分資源的掠奪性以及植物根系對養分的趨向性,我們推測,寄主植物在遭遇寄生危害時,會主動調節根系生長,增加對未被寄生部分的根系投資,以提高根系整體的養分吸收效率。在養分水平較高的條件下,增加的根系投資可獲取更多養分,這在一定程度上補償了寄生造成的養分損失;而在養分貧瘠條件下,這種補償效應不明顯。由于土壤養分的異質性分布特征,在有根部半寄生植物存在時,寄主根系常同時遭遇局部養分脅迫和寄生脅迫的雙重壓力。明確寄主植物根系在寄生脅迫下且養分異質分布的土壤環境中如何調節根系空間分布以及土壤養分變化對該行為的影響,對于了解寄主響應養分空間異質分布和寄生脅迫的適應策略,進而解析施肥對根部半寄生植物寄生危害的緩解效應十分必要。
本研究選擇滇西北亞高山草甸中兩種對禾草類寄主危害程度不同的馬先蒿,及其生境中常見、受馬先蒿寄生危害明顯的一種禾草類寄主,采用分室分根盆栽試驗,將寄主植物被寄生部分的根系與未被寄生部分的根系區分開來,一半寄主根系在養分匱乏的基質(不添加營養液)中生長,另一半根系在有養分供應(添加營養液)的基質中生長,探討土壤養分不均衡分配對其根系空間分配的影響。同時,在有養分供應的一側種植根部半寄生植物,探討兩種對寄主脅迫程度不同的根部半寄生植物分別寄生于寄主時,在不同土壤養分水平下對寄主根冠比和根系空間分布的影響。在馬先蒿廣泛分布的亞高山草甸中放牧活動比較頻繁,畜牧擾動和排放糞便導致土壤養分空間異質分布更為明顯。因此,本研究探討養分異質分布環境下馬先蒿寄生對寄主生長發育及根系空間分布的影響,將有助于從寄主根系空間分布變化了解養分對馬先蒿與禾草類寄主互作過程的調控作用,進而為有針對性地利用養分調節防控馬先蒿屬根部半寄生雜草提供理論指導。
1?材料與方法
1.1 材料
選用兩種根部半寄生植物,大王馬先蒿(Pedicularis rex)和三色馬先蒿(P.tricolor)開展試驗。其中,大王馬先蒿對寄主依賴程度較低,對寄主生長的抑制程度較輕,而三色馬先蒿對寄主依賴程度較高,對寄主生長的危害較重(Li et al., 2012a)。此外,兩種馬先蒿對養分需求也有所不同,前者對氮需求較高,后者對磷需求較高(Li et al., 2013b)。寄主植物則選擇兩種馬先蒿生境中的常見伴生植物長芒棒頭草(Polypogon monspeliensis)。
兩種馬先蒿的種子于2016年9月6日采自云南省迪慶州香格里拉高山植物園(99°38′E, 27°54′ N , 海拔3 370 m);寄主長芒棒頭草種子于2008年4月17日采自昆明植物園(102°44′ E, 25°08′ N, 海拔1 990 m);所有種子自然晾干后包于紙質信封中于4 ℃冰箱保存。
栽培基質使用沙土混合物,河沙和紅土體積比按9∶1充分混勻裝袋,于高壓滅菌鍋(溫度:121 ℃)中滅菌處理4 h,滅菌處理第二天再高溫(121 ℃)干燥處理4 h,自然冷卻后待用。栽培基質同Sui et al.(2019),有效氮、磷、鉀分別為14.3、2.7和62.4 mg·kg-1 干土重。
1.2 試驗設計
分室分根裝置參考Li et al.(2013a)的設計,有所改進。用透明膠帶將兩個方形塑料盆(上底寬10 cm, 下底寬7 cm, 高8.5 cm)粘在一起,然后將分根管套置于兩盆之間;在分根管內種植寄主植物(長芒棒頭草),在套盆一側不種馬先蒿的作為對照組處理(H),或者分別種植大王馬先蒿(PR+H)和三色馬先蒿(PT+H),共計三種組合。為設置異質的土壤養分環境,只在套盆一側添加營養液。寄生處理中,馬先蒿被種在添加營養液的一側,以考察土壤養分變化下長芒棒頭草對馬先蒿寄生脅迫的響應(圖1)。
基于前期研究結果,氮素供應水平對長芒棒頭草生長發育影響明顯,因此本試驗在添加營養液的一側設置3個氮素施用水平,即未施加外源氮(缺氮,-NPK)和分別澆施一倍氮(NPK)或兩倍氮(2NPK)的Long Ashton營養液,以考察氮供應變化對長芒棒頭草根系空間分布調節的影響。養分配比參考Li et al.(2013a),“NPK” 為Long Ashton標準營養液[2 mmol·L-1 K2SO4, 1.5 mmol·L-1 MgSO4·7H2O, 4 mmol·L-1 CaCl2, 0.1 mmol·L-1 FeEDTA, 4 mmol·L-1 (NH4)2SO4, 8 mmol·L-1 NaNO3, 1.33 mmol·L-1 Na2HPO4, 1.81 mg·L-1 MnCl2·4H2O, 0.5 mg·L-1 ZnSO4·7H2O, 0.08 mg·L-1 CuSO4·5H2O, 0.025 mg·L-1 NaMoO4·2H2O, 2.86 mg·L-1 H3BO3],“-NPK”表示去氮處理,“2NPK”表示氮加倍處理。共設9個處理,每處理5個重復。
1.3 馬先蒿和寄主種子萌發
先將兩種馬先蒿種子在75%的酒精中浸泡并搖床震蕩處理(120 r·min-1, 25 ℃)5 min,RO(Reverse Osmosis)水反復沖洗至無味,再用5%次氯酸鈉溶液消毒并搖床震蕩處理(120 r·min-1, 25 ℃)10 min,最后用RO水反復沖洗至無味后平鋪于三層濕潤濾紙上并置于4 ℃冰箱冷層積1周后放入恒溫培養箱(18 ℃/25 ℃, 高溫時12 h光照)進行萌發。長芒棒頭草種子經相同消毒處理后直接放入恒溫培養箱(18 ℃/25 ℃, 高溫時12 h光照)進行萌發。
1.4 寄主分根幼苗培育
待長芒棒頭草胚根長約2 cm時移栽至以石英砂為基質的一次性育苗盆中,每盆20株幼苗。寄主幼苗育苗期每周添加20 mL Long Ashton標準營養液(NPK)以滿足幼苗生長所需。每日澆水3次保證基質水分充足。每隔兩天調換育苗盆位置,排除位置效應對幼苗長勢的影響。在室溫條件下培養30 d后進行分根移栽。
1.5 幼苗移栽及栽培條件
待寄主幼苗根系長出8~10條側根時(育苗時間約30 d),小心取出并移栽到分根裝置中(側根被隨機均勻分至兩側),在套盆一側距分根管約2 cm處移栽3株馬先蒿。馬先蒿移栽時胚根長約2 cm,移栽后進行為期1周的遮蔭處理以提高存活率;1周后間苗,每盆保留一棵長勢一致的馬先蒿。每周在套盆種有馬先蒿的一側添加兩次營養液(-NPK、NPK、2NPK),每次15 mL,同時在套盆的另一側添加等量的自來水,分根管中添加2.5 mL自來水。試驗期間栽培基質水分保持在基質干重的10%左右。避免在添加營養液的當天澆水,以防止養分快速流失。每次添加營養液后隨機打亂套盆位置,排除位置效應。栽培試驗于2017年10月27日至2018年2月1日在昆明植物研究所玻璃精控溫室進行。試驗期間最低溫為8.9 ℃,最高溫為38.3 ℃,空氣相對濕度在21.3%~99.3%之間。
1.6 材料處理與數據分析
栽培過程中兩種馬先蒿前期長勢較好,后期長勢差且逐漸死亡。其中,三色馬先蒿在移栽第4周左右與寄主建立寄生關系,至第7周全部枯死(寄生時間約3周,死亡時生物量<0.001 8 g);大王馬先蒿在移栽第4周左右與寄主建立寄生關系,第10周全部枯死(寄生時間約6周,死亡時生物量<0.002 3 g)。移栽14周后收獲寄主植物。收獲時將地上部分和根系分開,其中套盆兩側寄主根系分別收獲并做記錄。將地上和地下部分于85 ℃干燥48 h烘干稱重。總根重為套盆兩側根系生物量之和。
使用SPSS 20.0對數據進行處理分析,用單因素方差分析法(One-way ANOVA)考察養分梯度對各部分生物量及根冠比的影響,用雙因素方差分析法(Two-way ANOVA)考察養分梯度與馬先蒿寄生對寄主生物量及根冠比的影響,利用t檢驗分析各栽培處理中分室兩側根系生物量的差異顯著性(P<0.05)。在進行單因素方差分析時對寄主兩側根系生物量分別進行了對數或平方根轉化,以滿足數據方差齊性和服從正態分布的檢驗假設。利用SPSS 20.0 和畫圖軟件(Adobe Illustrator)繪制和調整圖像。
2?結果與分析
2.1 土壤氮水平及馬先蒿寄生對長芒棒頭草生物量和根冠比的影響
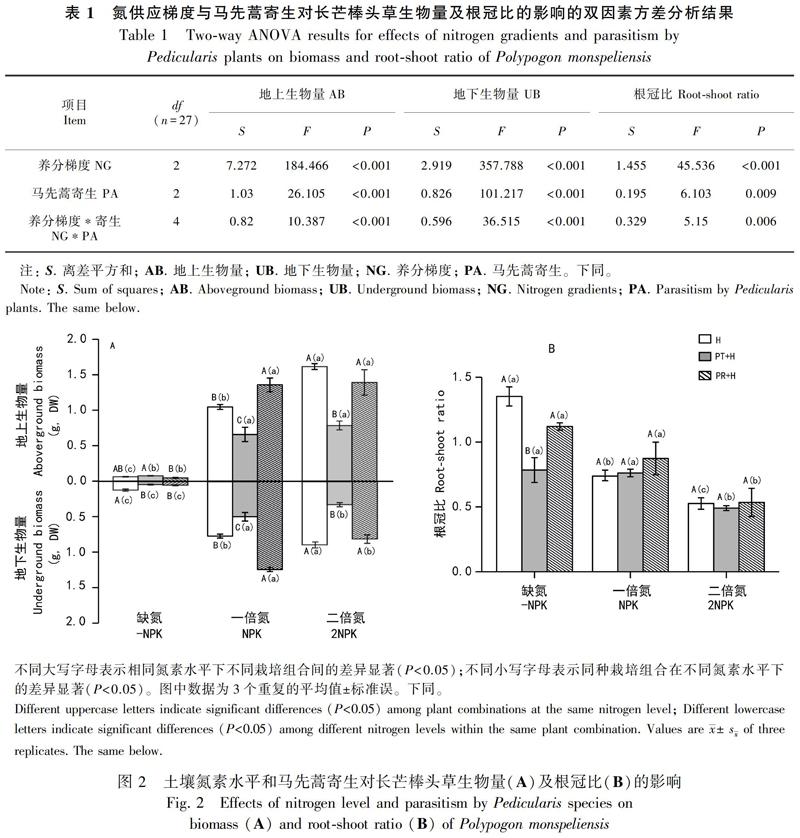
雙因素方差分析結果顯示,養分梯度與馬先蒿寄生對長芒棒頭草生物量和根冠比均存在顯著的交互影響,且養分梯度變化對長芒棒頭草地上部生物量、地下部生物量和根冠比的離差平方和(S養分梯度)分別為7.278、2.919和1.455,寄生對其所導致的離差平方和(S寄生)分別為1.030、0.826和0.195,均表現為S養分梯度>S寄生(表1)。
2.1.1 寄主(長芒棒頭草)生物量?養分添加對未被寄生的長芒棒頭草地上和地下生物量顯著影響,且隨著基質中氮素水平的增加,長芒棒頭草地上和地下部分生物量均顯著增加(圖2:A)。在不同養分條件下,兩種馬先蒿寄生對長芒棒頭草的影響表現出明顯的種間差異。未施加外源氮(缺氮,-NPK)處理下,三色馬先蒿寄生對長芒棒頭草地上部分生物量影響不明顯,但有增加趨勢,達22.7%;而大王馬先蒿寄生時,長芒棒頭草地上生物量只有0.046 g,較未被寄生時顯著降低了25.2%;兩種馬先蒿寄生均顯著降低長芒棒頭草地下生物量(PT:65.5%和PR:57.1%)。在NPK處理中,三色馬先蒿寄生時,長芒棒頭草地上和地下生物量分別為0.657 g和0.494 g,較未被寄生時顯著降低了37.1%和35.6%;而大王馬先蒿寄生卻使其地上和地下生物量分別達1.358 g和1.237 g,較未被寄生時顯著增加了29.9%和61.2%。2NPK處理下,被三色馬先蒿寄生的長芒棒頭草地上和地下生物量分別為0.784 g和0.325 g,較未被寄生時顯著降低了51.5%和63.5%;被大王馬先蒿寄生的長芒棒頭草地上、地下生物量分別為1.391 g和0.81 g,較未被寄生時降低13.9%和9%,但未達到顯著水平。
2.1.2 寄主(長芒棒頭草)根冠比?隨著土壤氮素水平的增加,未被寄生的長芒棒頭草根冠比顯著降低(圖2:B)。馬先蒿寄生對長芒棒頭草根冠比的影響因寄生植物種類和養分變化的不同而異。在未施加外源氮(缺氮,-NPK)處理下,三色馬先蒿寄生時長芒棒頭草根冠比為0.784,較未被寄生時顯著降低了42%;而大王馬先蒿寄生時長芒棒頭草根冠比為1.12,較未被寄生時僅降低了17.2%,未達統計學顯著水平。NPK處理下,三色馬先蒿和大王馬先蒿分別寄生使長芒棒頭草根冠比達0.761和0.873,較未被寄生時增加了3%和 18.5%,均未達統計學顯著水平。2NPK處理下,被三色馬先蒿寄生的長芒棒頭草根冠比為0.49,較未被寄生時僅降低了6.7%;而大王馬先蒿寄生時根冠比為0.534,較未被寄生時僅增加了1.6%,均未達統計學顯著水平(圖2:B)。
2.2 分室兩側寄主根系生物量分配對養分供應及馬先蒿寄生的響應
雙因素方差分析結果表明,馬先蒿寄生與養分供應狀況對套盆兩側長芒棒頭草根系生物量存在顯著交互影響(表2)。套盆未添加養分一側養分梯度貢獻的離差平方和(S養分梯度)為 0.336,寄生貢獻的離差平方和(S寄生)為0.125,在套盆添加養分一側S養分梯度為1.516,S寄生為0.261,套盆兩側均顯示出S養分梯度>S寄生。說明影響分室兩側根系生物量的主要因素為養分梯度。
T-test結果顯示,在未施加外源氮(缺氮,-NPK)處理下,三色馬先蒿寄生時,套盆兩側長芒棒頭草的根系生物量在未添加養分一側和添加養分一側分別為0.027 g和0.048 g,較未被寄生時分別增加了33%和2.2%;大王馬先蒿寄生時套盆兩側的長芒棒頭草根系生物量在未添加養分一側為0.021 g,較未被寄生時僅增加了3%,在添加養分一側為0.030 g,較未被寄生時卻降低了30%,但均未達統計學顯著水平。在NPK處理下,三色馬先蒿寄生時套盆兩側長芒棒頭草根系生物量在未添加養分一側和添加養分一側分別達0.146 g和0.348 g,較未被寄生時顯著降低45%和31%;大王馬先蒿寄生時套盆兩側根系生物量在未添加養分一側和添加養分一側分別達0.431 g和0.845 g,較未被寄生時顯著增加了63%和70%。在2NPK處理下,三色馬先蒿寄生時套盆兩側長芒棒頭草根系生物量在添加養分一側和添加養分一側分別為0.104 g和0.325 g,較未被寄生時顯著降低了60%和53%;大王馬先蒿寄生時套盆兩側根系生物量在未添加養分一側為0.324 g,較未被寄生時增加了24%,在添加養分一側為0.538 g,較未被寄生時降低了23%,但均未達到統計學顯著水平(圖3,表3)。不同養分梯度處理下各栽培組合(H、PT+H和PR+H)中長芒棒頭草根系在添加養分一側的生物量高于未添加養分一側的生物量(圖3)。未施加外源氮(缺氮,-NPK)處理下,H、PT+H和PR+H組合中在套盆中添加養分一側的長芒棒頭草根系生物量分別達0.046、0.048和0.030 g,較未添加養分的一側分別增加了132%、78.6%和46.4%。
NPK處理下,H、PT+H和PR+H組合中在套盆中添加養分一側的根系生物量分別達0.503、0.348和0.845 g,較未添加養分的一側分別增加了90.7%、139%和96.1%。2NPK處理下,H、PT+H和PR+H組合中在套盆中添加養分一側的根系生物量分別達0.695、0.325和0.538 g,較未添加養分的一側分別增加了165%、211%和66.2%(圖3)。
3?討論
本研究利用分室分根盆栽試驗,探討了寄主植物長芒棒頭草根系空間分布對土壤養分不均衡分布及馬先蒿寄生脅迫的響應,并分析了養分供應變化對這一響應的調節作用和對寄主生長的影響。寄主植物根系的生長和空間分布同時受養分空間分布和根部半寄生植物脅迫程度的影響,是寄主整合養分脅迫和寄生脅迫信息后主動調節根系生長和分配的結果。
三種養分梯度下添加養分一側長芒棒頭草根系生物量比未添加養分一側大,說明其根系趨向于養分較豐富區域生長。這與前人的研究結果一致,即根系更趨向于養分富集區域分布,以此來增加養分吸收面積進而吸取更多養分供其生長(Farley & Fitter 1999; Li et al., 2012b)。在未被馬先蒿寄生時,氮素水平增加使得添加營養液一側長芒棒頭草根系生物量顯著增加,并明顯增加養分貧瘠一側的根系生物量;進一步提升氮素水平雖然仍能明顯提高添加營養液一側的長芒棒頭草根系生物量,但養分貧瘠一側的根系生物量并未得到顯著提高。這說明長芒棒頭草根系具有主動的養分水平識別和根系生長調控能力,以減少低效率投資、避免資源浪費。
兩種馬先蒿對寄主植物生長的影響存在顯著差異。本研究中雖然三色馬先蒿寄生時間較短(僅3周),但其在NPK和2NPK處理下對長芒棒頭草生物量的抑制程度均大于寄生時間較長(約6周)的大王馬先蒿,這與Li et al.(2012a)研究結果一致,即前者較后者對寄主的依賴程度更強、造成的危害更大。未施加外源氮(缺氮,-NPK)處理下,三色馬先蒿和大王馬先蒿寄生導致長芒棒頭草向有馬先蒿一側的根系分配比例降低,提示當兩側根室均缺乏氮素時,長芒棒頭草根系有躲避馬先蒿寄生脅迫的傾向。然而,隨著氮素供應水平的提高,即便添加營養液一側存在馬先蒿寄生脅迫,長芒棒頭草也將更多的根系分配給了這一側。這可能是因為相對于養分貧瘠土壤的脅迫而言,在養分較為豐富的土壤環境下,寄生脅迫所造成的壓力相對較小。盡管沒有在保持兩側根室土壤養分完全均等的條件下探討長芒棒頭草根系分布對馬先蒿寄生脅迫的躲避行為,但根據其在缺氮處理中的表現及其他養分條件下被不同馬先蒿寄生時的根系生長反應差異來看,長芒棒頭草根系生長對寄生脅迫會表現出主動響應行為。
在未施加外源氮(缺氮,-NPK)處理下,大王馬先蒿寄生顯著抑制長芒棒頭草地上部分和根系的生物量,但在NPK處理時,卻顯著增加了地上及根系生物量。大王馬先蒿的氮素需求較高且較發達的根系可與寄主競爭養分(Li et al., 2013b),對同樣氮素需求較高的長芒棒頭草而言,在缺氮和寄生雙重脅迫條件下其生長受到了明顯的抑制。研究結果顯示,在土壤養分匱乏時,植物會通過加大對根系的投資來獲取更多的養分資源(Fransen et al., 1999; Craine 2006; Schenk, 2006)。在適當氮素供應條件下,大王馬先蒿對氮素的競爭導致栽培基質中氮素含量降低,從而促進了長芒棒頭草根系生長,再加上后期馬先蒿的死亡又在一定程度上緩解了其所受的寄生壓力,這可能是NPK處理下被大王馬先蒿寄生的長芒棒頭草生物量超過了未被寄生植株的重要原因。Cechin & Press(1993, 1994)認為合適的氮素供應可以降低寄生植物的寄生對寄主生物量的抑制甚至可以消除寄生效應。一般情況下,適量供應氮素可促進根系形態和質量的增加(Forde & Lorenzo, 2001),但過量的氮素供應會抑制根系生長(董桂春等,2001)。這也解釋了在NPK基礎上增加氮素供應反而降低了根系生物量的現象。
本研究中雖然兩種馬先蒿寄生的時間較短(三色馬先蒿寄生3周,大王馬先蒿寄生6周),但對長芒棒頭草生物量和根系發育的影響仍十分顯著。說明即便是短暫的寄生脅迫,也可對寄主植物生長造成嚴重危害。前人的研究顯示,根部半寄生植物可以通過吸器大量掠奪寄主植物資源、影響寄主植物光合能力、抑制寄主植物生長,對農業和畜牧業造成嚴重危害(Parker & Riches 1993; Shamoun, 2009; 隋曉琳,2013)。Suetsugu et al.(2012)與Bao et al.(2015)通過剔除根部半寄生植物證明了根部半寄生植物對禾草類和豆類優良寄主生長的抑制性。然而,這些試驗都未曾關注寄生植物幼苗對寄主植物生長的影響。由于一些根部半寄生植物種類種子產量大且存在一定程度的生理休眠和種子庫效應(柳妍妍等,2011;隋曉琳等,2013),因此,在野外生境中的整個植物生長季節內均有大量幼苗存在。由于幼苗個體不大、不易于觀察,其對生態系統的影響往往容易被忽視。本研究中,雖然大王馬先蒿和三色馬先蒿的寄生時間短且尚處于幼苗階段,但其對長芒棒頭草生長的影響依然存在。因此,在針對根部半寄生雜草防治過程中也要注意寄生植物幼苗的危害。
4?結論
(1)土壤氮水平與馬先蒿寄生均可顯著影響長芒棒頭草生物量及根冠比,且兩者存在顯著交互作用,其中土壤氮水平為主要影響因子。無論是否被馬先蒿寄生,提高土壤氮水平都會增加長芒棒頭草生物量,并有降低根冠比的趨勢。
(2)兩種馬先蒿短期寄生即可對長芒棒頭草造成明顯危害,但危害程度不同,且受土壤氮素水平調節的規律存在明顯種間差異。三色馬先蒿寄生在NPK和2NPK處理時顯著降低其生物量(莖葉:37.1%,51.5%;根系:35.6%,63.6%);而大王馬先蒿寄生在NPK處理時顯著增加其生物量(莖葉:29.9%,根系:61.2%)。
(3)長芒棒頭草可感應土壤氮素脅迫及馬先蒿寄生脅迫并相應調節根系空間分布。無論是否存在馬先蒿寄生脅迫,長芒棒頭草根系總是傾向于土壤氮水平較高的一側分布。在根室兩側均受氮素脅迫條件下,長芒棒頭草遭遇大王馬先蒿或三色馬先蒿寄生危害時,有向未被寄生根室增加根系生物量投資比例的傾向;但當被寄生一側的根室土壤氮水平較高時,長芒棒頭草根系躲避馬先蒿寄生的行為不明顯。長芒棒頭草根系空間分布變化對其適應土壤養分異質環境及馬先蒿寄生脅迫具有重要意義。
參考文獻:
AFLAKPUI GKS, GREGORY PJ, FROUD WILLIAMS RJ, 2002.Growth and biomass partitioning of maize during vegetative growth in response to Striga hermonthica infection and nitrogen supply[J].Exp Agric, 38:265-276.
BAO GS, SUETSUGU K, WANG XY, et al., 2015.Effects of the hemiparasitic plant Pedicularis kansuensis on plant community structure in a degraded grassland[J].Ecol Res, 30:507-515.
BENNING TL, SEASTEDT TR, 1997.Effects of fire, mowing and nitrogen addition on root characteristics in tall-grass prairie[J].J Veg Sci, 8:541-546.
CECHIN I, PRESS MC, 1993.Nitrogen relations of the sorghum Striga hermonthica host parasite association germination, attachment and early growth[J].New Phytol, 124:681-687.
CECHIN I, PRESS MC, 1994.Influence of nitrogen on growth and photosynthesis of a C-3 cereal,Oryza sativa, infected with the root hemiparasite Striga hermonthica[J].J Exp Bot, 45:925-930.
CRAINE JM, 2006.Competition for nutrients and optimal root allocation[J].Plant Soil, 285:171-185.
DAY KJ, JOHN EA, HUTCHINGS MJ, 2003.The effects of spatially heterogeneous nutrient supply on yield, intensity of competition and root placement patterns in Briza media and Festuca ovina[J].Funct Ecol, 17:454-463.
DONG GC,WANG YL, WU H, et al., 2001.Effect of nitrogen supplying levels on the development of roots in rice(Oryza satival)[J].Jiangsu Agric Res, (4):9-13.[董桂春,王余龍,吳華,等,2001.供N濃度對水稻根系生長的影響[J].江蘇農業研究,(4):9-13.]
FARLEY RA, FITTER AH, 1999.Temporal and spatial variation in soil resources in a deciduous woodland[J].J Ecol, 87:688-696.
FORDE B, LORENZO H, 2001.The nutritional control of root development[J].Plant Soil, 232:51-68.
FRANSEN B, BLIJJENBERG J, DEKROON H, 1999.Root morphological and physiological mplasticity of perennial grass species and the exploitation of spatial and temporal heterogeneous nutrient patches[J].Plant Soil, 211:179-189.
FROST DL, GURNEY AL, PRESS MC, et al., 1997.Striga hermonthica reduces photosynthesis in sorghum:The importance of stomatal limitations and a potential role for ABA?[J].Plant Cell Environ, 20:483-492.
GIBSON CC, WATKINSON AR, 1991.Host selectivity and the mediation of competition by the root hemiparasite Rhinanthus minor[J].Oecologia, 86:81-87.
HEARNE SJ, 2009.Control the Striga conundrum[J].Pest Manag Sci, 65:603-614.
JACKSON RB, CALDWELL MM, 1993.The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics[J].Ecology, 74:612-614.
LI AR, GUAN KY, STONOR R, et al., 2013a.Direct and indirect influences of arbuscular mycorrhizal fungi on phosphorus uptake by two root hemiparasitic Pedicularis species:Do the fungal partners matter at low colonization levels?[J].Ann Bot, 112:1089-1098.
LI AR, LI YJ, SMITH SE, et al., 2013b.Nutrient requirements differ in two Pedicularis species in the absence of a host plant:Implication for driving forces in the evolution of host preference of root hemiparasitic plants[J].Ann Bot, 112:1099-1106.
LI AR, SMITH FA, SMITH SE, et al., 2012a.Two sympatric root hemiparasitic Pedicularis species differ in host dependency and selectivity under phosphorus limitation[J].Funct Plant Biol, 39:784-794.
LI HB, ZHANG FS, SHEN JB, 2012b.Contribution of root proliferation in nutrient-rich soil patches to nutrient uptake and growth of maize[J].Pedosphere, 22:776-784.
LI HB, 2014.The dynamic process of plant responses to heterogeneous nutrient environments and the strategies of root management[D].Beijing:China Agricultural University.[李洪波, 2014.植物響應養分空間異質性分布的動態過程及調控根系獲取養分的策略研究[D].北京:中國農業大學.]
LIU YY, HU YK, WANG X, et al., 2011.Studies on characteristics of soil seed bank of Pedicularis verticilata community[J].J Nat Res, 26(1):48-57.[柳妍妍,胡玉昆,王鑫,等,2011.輪葉馬先蒿群落土壤種子庫特征研究[J].自然資源學報,26(1):48-57.]
LIU YY, TAXIPULATI T, GONG YM, et al., 2017.N-P fertilization inhibits growth of root hemiparasite Pedicularis kansuensis in natural grassland[J].Front Plant Sci, 8:2088.
PARKER C, RICHES CR, 1993.Parasitic weeds of the world:Biology and control[M].Wallingford:CAB International:1-332.
PRESS MC, 1989.Autotrophy and heterotrophy in root hemiparasites[J].Trends Ecol Evol, 4:258-263.
PRESS MC, PHOENIX GK, 2005.Impacts of parasitic plants on natural communities[J].New Phytol, 166:737-751.
SCHENK HJ, 2006.Root competition:Beyond resource depletion[J].J Ecol, 94:725-739.
SCHIFFERS K, TIELBORGER K, TIETJEN B, et al., 2011.Root plasticity buffers competition among plants:Theory meets experimental data[J].Ecology, 92:610-620.
SHAMOUN SF, 2009.Introduction to stem and shoot fungal pathogens and parasitic plants:the values of biological diversity introduction[J].Botany, 87:5-6.
SHAO YQ, ZHAO J, LIU ZL, et al., 2006.Effects of mowing frequency on soil microbial quantity and plant underground biomass in Leymus chinensis steppe[J].Chin J Ecol, (10):1191-1195.[邵玉琴,趙吉,劉鐘齡,等,2006.割草頻率對土壤微生物數量和植物地下生物量的影響[J].生態學雜志,(10):1191-1195.]
SMALING EMA, STEIN A, SLOOT PHM, 1991.A statistical analysis of the influence of Striga hermonthica on maize yields in fertilizer trials in southwestern Kenya[J].Plant Soil, 138:1-8.
STEWART GR, PRESS MC, 1990.The physiology and biochemistry of parasitic angiosperms[J].Ann Rev Plant Physiol Plant Mol Biol, 41:127-151.
SUETSUGU K, TAKEUCHI Y, FUTAI K, et al., 2012.Host selectivity, haustorial anatomy and impact of the invasive parasite Parentucellia viscosa on Floodplain vegetative communities in Japan[J].Bot J Linn Soc, 170:69-78.
SUI XL, 2013.Eco-physiological mechanism for the spatial expansion of Pedicularis kansuensis in Bayanbulak grassland of Xinjiang[D].Beijing:University?of Chinese Academy Science.[隋曉琳, 2013.甘肅馬先蒿在新疆巴音布魯克草原蔓延的生理生態學機理[D].北京:中國科學院大學.]
SUI XL, LI AR, GUAN KY, 2013.Impacts of climatic changes as well as seed germination characteristics on the population expansion of Pedicularis verticillata [J].Ecol Environ Sci, 22(7):1099-1104.[隋曉琳,李愛榮,管開云,2013.氣候變化和種子萌發特性對輪葉馬先蒿種群擴張的影響[J].生態環境學報,22(7):1099-1104.]
SUI XL, ZHANG T, TIAN YQ, et al., 2019.A neglected alliance in battles against parasitic plants:Arbuscular mycorrhizal and rhizobial symbioses alleviate damage to a legume host by root hemiparasitic Pedicularis species[J].New Phytol, 221:470-481.
WANG W, LIANG CZ, LIU ZL, et al., 2000.Analysis of the plant individual behavior during the degradation and restoring succession in steppe community[J].Chin J Plant Ecol, 24(3):268-274.[王煒,梁存柱,劉鐘齡,等,2000.草原群落退化與恢復演替中的植物個體行為分析[J].植物生態學報,24(3):268-274.]
WANG WX, SANG GJ, LI L, 2009.Study on the control techniques of poisons grass Pedicularis in Xinjiang Bayanbulak prairie[J].Grass-Feeding Livestock, (2):49-50.[王文香,桑格吉,李莉,2009.新疆巴音布魯克草原毒害草馬先蒿防治技術研究[J].草食家畜,(2):49-50.]
WIJESINGHE DK, JOHN EA, BEURSKENS S,et al., 2001.Root system size and precision in nutrient foraging:Responses to spatial pattern of nutrient supply in six herbaceous species[J].J Ecol, 89:972-983.
ZHANG HM, FORDE BG, 1998.An arabidopsis MADS box gene that controls nutrient induced changes in root architecture[J].Science, 279:407-409.
(責任編輯?何永艷)
