全面了解,整體掌握
——HIV命題多視角分析
2017-08-08 03:01:37貴州鄭小毛
教學考試(高考生物) 2017年2期
貴州 鄭小毛
全面了解,整體掌握
——HIV命題多視角分析
貴州 鄭小毛
隨著經濟和社會的發展,健康逐漸成為了生活的主題,國家和個人都越來越關注疾病(特別是傳染病)的預防和治療。艾滋病,自從出現以來,一直都是人類社會關注的熱點,作為當前人類健康的頭號殺手,預防和治療艾滋病,一直都是科學追求的目標。
把健康貫穿于教育教學,用考試指揮棒引導公民更加關注健康知識,是當下教育變革的方向。因此,健康、疾病、HIV……這些知識,也就成了高考命題取材的重點和熱點。
一、HIV的高考考查及知識點歸納
1. HIV的高考考查
從過去幾年高考試題統計分析來看,考查HIV的試題主要有:2016年高考江蘇選擇題13,2015年高考浙江選擇題1,2015年全國課標卷Ⅱ選擇題5,2015年全國課標卷Ⅰ非選擇題40,2013年全國課標卷Ⅰ選擇題4, 2013年全國大綱卷選擇題3,2007年廣東和海南都進行了非選擇題的考查。
“艾滋病的流行和預防”在2017年《考試大綱》中,要求層級由Ⅰ上升到Ⅱ,意味著在2017年,在各套高考試題中,依然會出現HIV相關的試題。
2. HIV的知識點歸納
2.1 HIV科普知識
HIV,人類免疫缺陷病毒,俗稱艾滋病病毒,屬于RNA病毒,主要攻擊人體T細胞,引起免疫缺陷病,是造成人類免疫系統缺陷的一種病毒。1981年,人類免疫缺陷病毒在美國首次發現。它是一種感染人類免疫系統細胞的慢病毒(Lentivirus),屬逆轉錄病毒的一種。

HIV通過破壞人體的T淋巴細胞,進而阻斷細胞免疫和體液免疫過程,導致免疫系統癱瘓,致使各種疾病在人體內蔓延,最終導致艾滋病。由于HIV的變異極其迅速,難以生產特異性疫苗,至今無有效治療方法,對人類健康造成極大威脅。

2.2 HIV的致病機理2.3 HIV及其他病毒拓展
(1)有關病毒的幾個常識
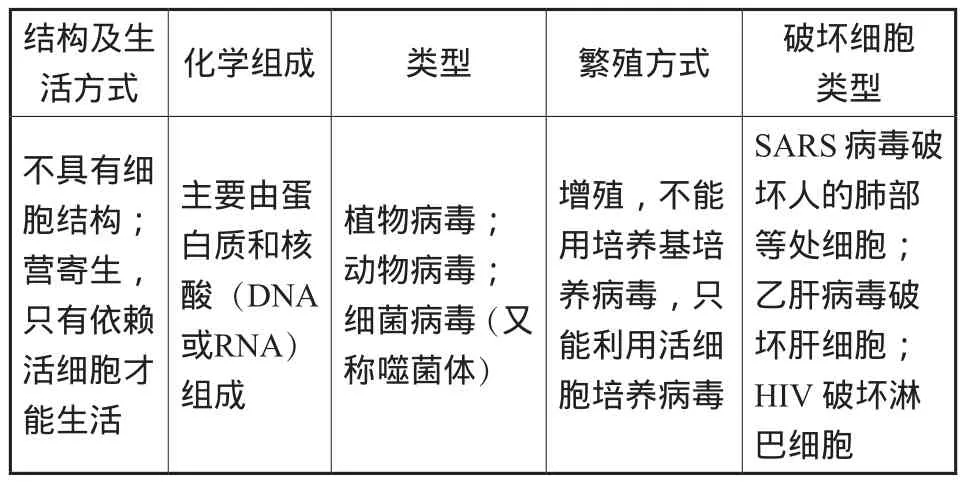
_活方式_____________________________________化學組成 類型 繁殖方式 破壞細胞類______型不具有細胞結構;營寄生,只有依賴活細胞才能生活結構及生主要由蛋白質和核酸(DNA或RNA)組成植物病毒;動物病毒;細菌病毒(又稱噬菌體)增殖,不能用培養基培養病毒,只能利用活細胞培養病毒SARS病毒破壞人的肺部等處細胞;乙肝病毒破壞肝細胞;HIV破壞淋________________________巴細胞
(2)病毒與免疫調節的聯系
病毒作為一種抗原,決定其抗原特異性的是衣殼(蛋白質),因此可利用病毒衣殼的抗原特異性功能制備疫苗,進行免疫預防。病毒侵入機體后,先進行體液免疫,再進行細胞免疫。治療病毒性感染不能使用抗生素類藥物,應該使用干擾素、病毒靈等抗病毒藥物。
(3)病毒與中心法則的拓展
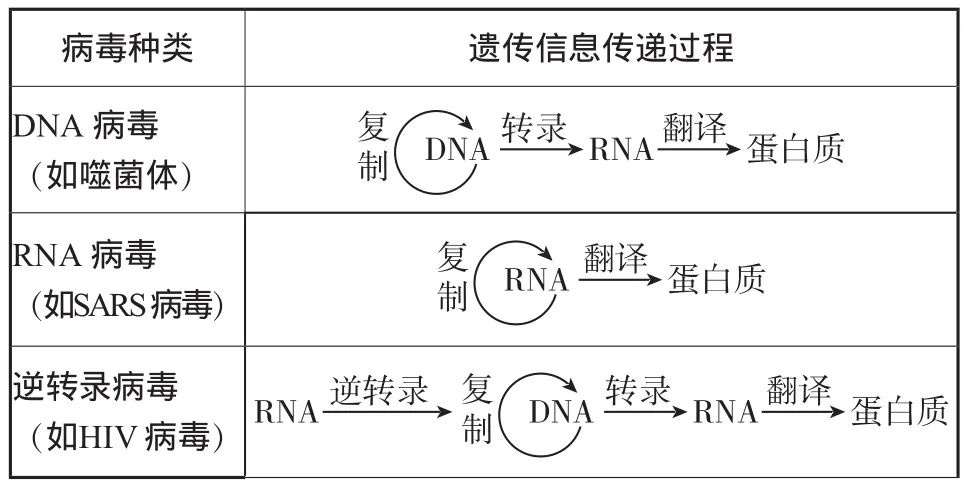
__病毒種類 _____________遺傳信息傳遞過程DNA病毒(如噬菌體)_復制DNA RNA轉錄 翻譯 蛋白質RNA病毒(如SARS病毒)復制RNA 翻譯 蛋白質逆轉錄病毒(如HIV病毒)RNA逆轉錄 復制DNA RNA轉錄 翻譯 蛋白質
二、HIV的命題視角及命題預測
1. HIV的命題視角分析
1.1 結合病毒與原核、真核生物的比較考查
HIV屬于病毒,沒有細胞結構,由RNA和蛋白質組成,與原核、真核生物的細胞結構差別較大,預測2017年高考可能聯系HIV與原核、真核細胞結構上的區別考查HIV組成。
例1(2013·大綱卷)關于HIV的敘述,正確的是 ( )
A. HIV在活細胞外能大量增殖
B. HIV僅含有核糖體這一種細胞器
C. HIV主要攻擊B細胞,使人體無法產生抗體
D. 艾滋病患者的血液中可以檢出HIV這種病毒
【解析】HIV是病毒,必須依賴活細胞才能大量繁殖,A錯誤;HIV不含有核糖體,B錯誤;HIV主要攻擊T細胞,C錯誤;HIV通過體液傳播,所以在患者的血液中能夠檢出HIV,D正確。
【答案】D
1.2 結合中心法則考查
HIV侵入機體后會進行侵染、增殖等一系列生命活動,如下圖所示,這些生命活動都與中心法則息息相關,預測2017年高考可能聯系中心法則,考查HIV在人體細胞中的增殖過程。
例2 下圖是HIV感染人體細胞過程的示意圖。請回答下列一組問題。
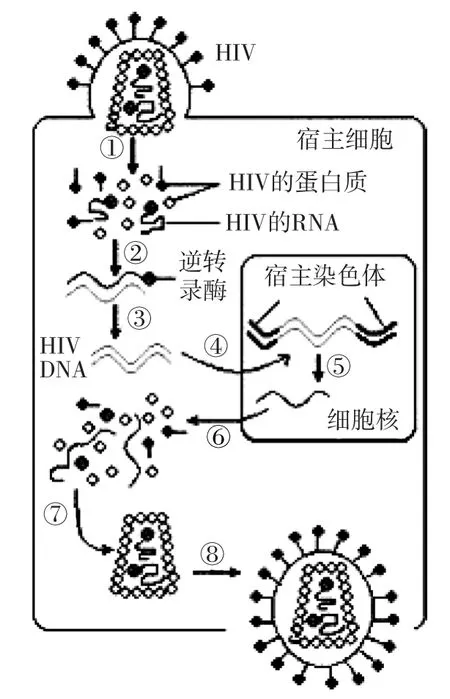
(1)由于病毒在結構和習性上的特殊性,人們對生物進化過程中是先有病毒后有細胞還是先有細胞后有病毒這一問題,存在兩種截然不同的觀點。你的觀點是________,理由是_______。
(2)HIV進入人體后,由于與T細胞表面的CD4具有較強親和力而容易侵染T細胞。CD4的化學成分最可能是_______。
(3)完成圖中的②過程,需要宿主細胞提供的原料是________。直接參與⑥過程的兩種細胞器是________。
(4)圖中①~⑧所示的過程中,遵循堿基互補配對原則的是_______。
(5)大多數艾滋病患者會出現記憶喪失等精神障礙,此時HIV已感染了_______。
(6)人的免疫系統是如何清除進入T細胞內的HIV的?請簡要說明。
【解析】本題考查中心法則中的DNA復制、轉錄、翻譯、逆轉錄所需的模版、原料、產物等問題,熟悉中心法則的各個過程是正確解答該題的關鍵。
【答案】(1)觀點:先有病毒后有細胞;理由:病毒的結構較細胞的簡單。(或觀點:先有細胞后有病毒; 理由:病毒不能離開細胞而獨立生存和繁殖。)
(2)蛋白質
(3)4種脫氧核苷酸 核糖體、線粒體
(4)②③⑤⑥
(5)大腦
(6)首先由相應的效應T細胞與靶細胞密切接觸而使靶細胞裂解,釋放出HIV;然后由體液中的特定抗體與HIV結合,再由吞噬細胞吞噬、清除(須說明效應T細胞、抗體、吞噬細胞的作用)
1.3 對HIV與免疫的考查
HIV主要攻擊人體T細胞,進而阻斷細胞免疫和體液免疫過程,導致免疫系統癱瘓,引起免疫缺陷病,對人類健康造成極大威脅。預測2017年高考可能結合免疫調節考查HIV的致病機理及引起的免疫反應。
例3(2016·江蘇卷)人類免疫缺陷病毒(HIV)有高度變異性,感染機體后可損傷多種免疫細胞,并通過多種機制逃避免疫系統識別和攻擊。下列相關敘述錯誤的是 ( )
A. HIV感染人群比健康人群更易患甲型H1N1流感
B. HIV的高度變異性,致使疫苗效果難以持久
C. 被HIV潛伏感染的細胞表面沒有HIV蛋白,利于病毒逃避免疫系統識別和攻擊
D. HIV破壞免疫系統,機體無體液免疫應答,不能通過檢測抗體來診斷HIV感染
【解析】HIV感染人群中多種免疫細胞損傷,對H1N1流感的抵抗力下降,A正確;接種疫苗產生的抗體和記憶細胞具有特異性,HIV具有高度變異性,導致記憶細胞和抗體不能消滅HIV,B正確;被HIV潛伏感染的細胞表面沒有HIV蛋白,有利于病毒逃避免疫系統識別和攻擊,C正確;HIV感染初期,機體通過體液免疫產生大量抗體,可通過檢測抗體來診斷HIV感染,D錯誤。
【答案】D
1.4 對HIV的綜合考查
由于HIV所涉及的知識點比較雜,涉及病毒的結構、病毒的繁殖、病毒的感染與免疫等有關內容。在高考考查中許多時候進行綜合考查,試題與各方面相關聯,主要為了引起對HIV的關注和重視。
例4(2015·新課標卷Ⅰ)HIV屬于逆轉錄病毒,是艾滋病的病原體。回答下列問題:
(1)用基因工程方法制備HIV的某蛋白(目的蛋白)時,可先提取HIV中的_______,以其作為模板,在_______的作用下合成________。獲取該目的蛋白的基因,構建重組表達載體,隨后導入受體細胞。
(2)從受體細胞中分離純化出目的蛋白,該蛋白作為抗原注入機體后,刺激機體產生的可與此蛋白結合的相應分泌蛋白是________。該分泌蛋白可用于檢測受試者血清中的HIV,檢測的原理是________。
(3)已知某種菌導致的肺炎在健康人群中罕見,但是在艾滋病患者中卻多發。引起這種現象的根本原因是HIV主要感染和破壞了患者的部分________細胞,降低了患者免疫系統的防衛功能。
(4)人的免疫系統有_______癌細胞的功能,艾滋病患者由于免疫功能缺陷,易發生惡性腫瘤。
【解析】(1)HIV屬于逆轉錄病毒,它的遺傳物質是RNA,在逆轉錄酶的作用下可以逆轉錄為cDNA,用于構建基因表達載體,從而制備HIV的某蛋白。(2)HIV的某蛋白作為抗原進入機體后,能刺激人體產生針對該抗原的一種特殊的分泌蛋白——抗體;可用該抗體進行抗原—抗體進行雜交來檢測血清中是否含有HIV。(3)HIV病毒營寄生生活,寄生在T淋巴細胞內,T細胞參與體液免疫和細胞免疫,因此少了T細胞,特異性免疫幾乎全部喪失,降低了機體的免疫功能。(4)人體的免疫系統具有防衛、監控和清除的功能,可及時清除體內產生的癌細胞,艾滋病患者的整個免疫功能缺陷,機體腫瘤的發生機會增加。
【答案】(1)RNA 逆轉錄酶 cDNA(或DNA)
(2)抗體 抗原抗體特異性結合
(3)T(或T淋巴)
(4)監控和清除
2.HIV的命題預測
艾滋病作為危害人類健康的頭號殺手,也是近年高考命題的熱點專題之一,以艾滋病病毒的結構、繁殖過程考查免疫、基因的表達是常見的題型。掌握教材相關知識,加以拓展,認真回顧高考真題,強化訓練是全面了解、整體把握該部分知識的重要措施。
(作者單位:貴州省興義中學)
