非淀粉多糖在豬腸道菌群調控中的研究進展
2020-02-22 04:48:24楊立杰曾祥芳譙仕彥
生物技術通報 2020年2期
關鍵詞:研究
楊立杰 曾祥芳 譙仕彥
(中國農業大學動物科技學院 動物營養學國家重點實驗室,北京 100193)
日糧中纖維水平過高,將影響能量及蛋白質等營養物質的消化率,降低飼料轉化效率[1-2]。然而,纖維在動物營養調節方面的研究結果存在不一致性。近期研究表明,日糧中適宜的纖維水平,對促進畜禽腸道功能發育及其微生態平衡具有積極作用[3-6]。纖維源及其中非淀粉多糖(Non-starch polysaccharides,NSP)組分含量的差異,是導致其應用效果呈現多樣性的重要原因,亦是其功能性評價成敗的關鍵[7-8]。眾多研究表明,日糧纖維可以通過調節腸道菌群結構[9-11],減少斷奶仔豬腹瀉,進而提高生產性能[12],是促生長型抗生素的理想替代物[13-14]。而NSP作為膳食纖維的重要組成部分,其組分對于纖維的功能性反饋及其發揮的程度都至關重要。另有報道顯示,NSP對豬腸道菌群正面調控效應的體現,主要與腸道微生物對NSP的利用形式及其代謝產物密切相關[15]。豬腸道微生物通過自身糖苷酶的分泌,降解NSP結構中的β或α糖苷鍵,從而產生單糖和寡糖供微生物利用[16]。此外,日糧NSP可為豬后腸段微生物提供良好的培養基質[17-18],同時發酵產生的乙酸、丙酸、丁酸等短鏈脂肪酸(Short chain fatty acid,SCFA)可通過降低腸道pH以抵御感染及致病菌的侵襲[19-21]。
綜上所述,腸道菌群調控的機制探索,已成為整個微生物研究最活躍的熱點之一。然而纖維對豬腸道菌群調控機制闡釋才剛剛開始,許多問題亟待解決。由此,重新定位并積極探索NSP對豬腸道菌群的調控機制,無疑是對NSP與菌群間對話機制深入認識的重中之重。
1 NSP的構成及其評價體系
1.1 NSP的構成
作為高等植物細胞壁的主體,纖維的組成復雜多樣,作用和功能各不相同,主要由NSP和木質素組成。其中NSP的占比可高達90%以上[22],主要包括纖維素、半纖維素、寡聚糖、抗性淀粉及果膠類等(圖1),是纖維中相對容易被動物消化的類型。根據NSP是否具有溶解于水或弱堿溶液的特性,又可分為可溶性非淀粉多糖(Soluble nonstarch polysaccharides,SNSP)和不可溶性非淀粉多糖(Insoluble non-starch polysaccharides,INSP)。

圖1 植物碳水化合物的組成
纖維素主要由葡萄糖通過β-(1,4)糖苷鍵彼此鏈接而成,而NSP中的非纖維素部分則由不同的單糖通過 β-(1,4)、β-(1,3)、β-(2,1)及 α-(1,6)等糖苷鍵異構性連接而成,是目前發現異構型及締合分子最多的物質之一(圖2)。
1.2 NSP評價體系
以往報道中,研究者普遍采用粗纖維(Crude fiber,CF)、中性洗滌纖維(Neutral detergent fiber,NDF)及酸性洗滌纖維(Acid detergent fiber,ADF)等作為評價日糧纖維水平的參考指標。然而,近幾年隨著精準營養及新舊動能轉換行動的倡導,合理開發并利用“纖維”,這一地球上儲量最多的可再生資源,便成為了當前科技工作者亟待解決的首要任務。原始的纖維評價體系,已遠遠無法適應當前精準營養的需求。
CF評價體系中,酸性洗滌劑和堿性洗滌劑的處理,將分別使得樣品中半纖維素和木質素含量大量損失,而半纖維素中的阿拉伯木聚糖、β-葡聚糖、甘露聚糖等作為其主要組成部分,均具有可溶于水的特性,亦屬于可溶性NSP范疇。因此,在評價含可溶性纖維較多的原料時,使得CF檢測值遠低于真實值。報道表明,《中國飼料成分及營養價值表(27版)》中,NY/T 1級豆粕中CF含量為5.9%(風干基礎),而在黃慶華[8]的研究中顯示,豆粕中僅NSP含量就已達到20.3%(風干基礎,水分以10%計),相似的結果在Choct等[23]的研究中也得到了證實。此外,在NDF和ADF體系中,研究者盡管能夠對纖維含量進行相對客觀的評價,但對纖維組分構成,仍不能進行進一步檢測,且在檢測過程中果膠等可溶性纖維的損失,難以避免。由此,“酶-氣相色譜-比色-重量法”應運而生。該法將NSP酶解后,酸性條件下水解為單糖,中性糖衍生為糖醇乙酸酯,糖醛酸在酸性條件下經3,5-二甲基酚顯色后定量,可對纖維及其組分的實際含量進行準確測定,是揭開纖維研究黑匣子的重要研究手段。20%[29-31]。NSP被后豬腸段微生物利用,同時產生乙酸、丙酸和丁酸等SCFA被腸上皮細胞吸收利用。研究表明,豬體內丁酸主要被結腸上皮細胞吸收,而丙酸、戊酸等被吸收入血后進入肝臟進行丙酸糖異生過程,乙酸則被肌肉和脂肪組織利用[32-33]。此外,有研究表明,日糧NSP可以通過參與激活腸道中相關通路進行腸道免疫和代謝的調控,且其對機體的影響包括但不僅限于其自身的能量供應[34]。
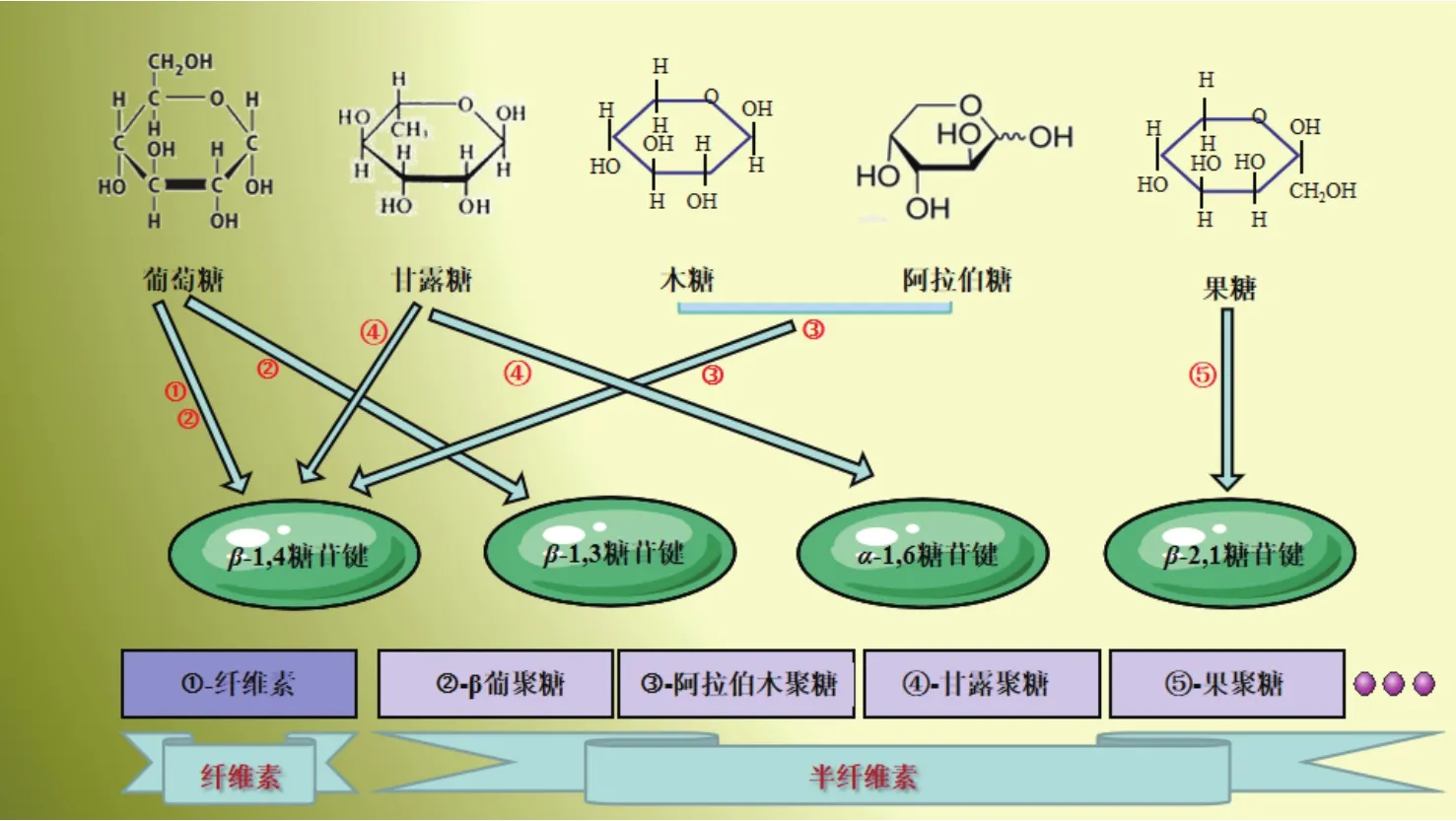
圖2 纖維素及半纖維素的構成
2 NSP的營養學功能
豬自身不具備分泌纖維素酶、半纖維素酶和果膠酶的能力,因此日糧NSP不能被豬自身分泌的消化酶降解,而主要依賴于腸道微生物的發酵和非酶水解作用[24],其發酵程度與日糧中ADF水平呈負相關[25-26]。隨著日齡的增加,豬大腸內具有NSP降解能力的菌群數量逐漸增加,對日糧NSP的降解能力亦將逐步提高[27]。研究表明,生長豬腸道內的NSP分解菌數量約為1×108CFU/g,而成年母豬大腸中的NSP分解菌數量是生長豬的6倍[28]。
豬后腸道微生物以日糧NSP作為主要碳源并最終將其轉化為SCFA,為宿主提供所需總能量的5%-
3 NSP對豬腸道微生物組成及代謝的影響
3.1 NSP研究的必要性
近年來,隨著醫學領域對肥胖、糖尿病與日糧纖維及腸道菌群間互作關系的深入研究,日糧NSP及其代謝產物與腸道菌群間關系的探討備受眾多研究者青睞[35-38]。研究表明,NSP在調節動物腸道菌群結構、抑制病源微生物的定植及維持腸道健康等諸多方面日益體現出不可替代的作用[39-41]。NSP及木質素的構成、來源及其對動物代謝的影響,見表1。豬腸道微生物菌群數量及豐度,依賴于日糧結構及營養素變化[42-45],其中NSP是其后腸段微生物的主要能量來源[46]。日糧中不同含量的NSP被豬攝入后,隨食糜進入后腸段被微生物利用,對豬后腸段微生物菌群結構及其代謝活性造成可逆性影響,從而改變菌群代謝產物[47]。對于豬腸道微生物而言,NSP是除木質素外唯一一種幾乎不受上消化道及前腸段干擾,組分幾近無損作用于腸道菌群的營養素,推測可嘗試作為腸道微生物區系調控的標志性營養素進行深入探究。近期眾多研究中,多采用微晶纖維素及羧甲基纖維素作為日糧纖維源,研究其對動物腸道健康的影響[8,48]。以上兩種纖維素,組成單糖均為葡萄糖,盡管能夠在一定程度上說明纖維素對機體的影響,但由于忽略了纖維中NSP及其組分含量和比例的多樣性,故結論并不嚴謹。

表1 纖維亞型的構成、來源及功能[49-52]
3.2 NSP對斷奶仔豬及生長育肥豬腸道菌群的調控
斷奶應激是促使仔豬腸道菌群建立穩態及多樣性的重要階段,而不合理的營養調控無疑是導致仔豬腹瀉的始作俑者。研究表明,日糧中NSP可黏附于腸黏膜表面,對外源病菌的吸附和定植起到一定的防御作用[53]。此外,不同腸段微生物具備定向選擇NSP并利用其不同組分的能力,所以可根據NSP類型及其組分的不同,對豬腸道菌群結構進行調控[54]。
根據溶解性,NSP可分為INSP和SNSP。其中,SNSP最早可在回腸被微生物開始利用,而INSP卻能夠為直至大腸末端的微生物提供培養基[55],提示NSP對豬腸道微生物的調控,不僅與NSP來源及含量有關,還與INSP/SNSP,甚至NSP組分密切相關。SNSP可在后腸菌群的作用下迅速發酵并產生SCFA,且不同NSP源產生的SCFA種類和濃度完全不同。而INSP可縮短食糜在腸道中的停留時間,促進腸蠕動,減少糞便在體內的滯留時間,是營養素調控腸道健康及菌群結構的重要手段[56-57]。研究表明,日糧中添加10%小麥麩,可顯著提高斷奶仔豬回腸乳酸桿菌和結腸中雙歧桿菌的數量,降低腹瀉率[58]。另有報道表明,選擇大豆皮為NSP源,使得豬回腸段乳酸桿菌數量顯著降低,而盲腸中大腸桿菌數量顯著升高[59]。推測原因除了大豆皮NSP高于小麥麩外(56.7%>30.6%),也與兩者INSP/SNSP(小麥:26.5∶4.1;大豆皮 :48.3∶8.4)有關[60]。此外,小麥麩中NSP主要以阿拉伯木聚糖為主,而大豆皮中NSP主要為半乳甘露聚糖和果膠[61],暗示NSP組分的不同是兩者發揮不同功能性的重要潛在原因之一。另有報道顯示,高NSP水平日糧能夠顯著增加腸道中具有高效纖維分解能力的菌群豐度[62]。SNSP可通過增加食糜黏稠性,影響機體消化功能[63-64]。同時,腸道厭氧環境使其蠕動遲緩,為后腸段微生物提供了穩定的發酵環境[65]。龔敏[66]試驗表明,日糧中小麥NSP的含量對斷奶仔豬回腸、結腸、盲腸中雙歧桿菌、乳酸桿菌、大腸桿菌及鏈球菌數量影響并不顯著。而小麥日糧中木聚糖酶的添加使雙歧桿菌和乳酸桿菌數量呈現出一定的升高趨勢,大腸桿菌及鏈球菌呈現降低趨勢。提示斷奶仔豬腸道發育不完善,NSP酶缺乏,可能是導致日糧中小麥NSP難以發揮功能性的重要原因,推測育肥豬日糧中添加適宜劑量的NSP,預期將獲得積極效應,這在高陽等[67]的研究中得到證實。豬育肥階段腸道發育完善,菌群相對穩定,當采食高NSP日糧時,腸道菌群需要一定時間進行自我調整,以抵御原有微生物區系的冗余特性并適應新的發酵基質。有研究表明,育肥豬飼糧中添加50%苜蓿,其后腸道微生物總數呈現出先減少后增加的趨勢,且具有纖維素高效降解能力的菌屬比例不斷增加[68]。采用不同纖維類型及玉米粉碎粒度作為自變量,對育肥豬日糧進行處理后,支鏈及直鏈淀粉酶在試驗第1天出現,而木聚糖酶等相關NSP酶在試驗第7天才開始出現[69],提示豬腸道菌群對日糧NSP的反饋,是一個相對緩慢的過程。
3.3 NSP對妊娠母豬腸道菌群的調控
在非洲豬瘟疫情的影響下,我國能繁母豬存欄量驟減,生豬價格翻倍,對民眾的生活造成了一定的影響。全面的認識NSP在母豬營養調控中的作用并合理利用,對于指導生產和提高其生產性能具有重要的意義。母豬后腸段具有龐大的微生物群(1×1010-11CFU/g),是生長豬的6-7倍,且食糜在其后腸段滯留時間長(20 h左右),因而母豬對NSP的利用具有相較于生長豬更大的潛力。妊娠期母豬腸道微生物區系的健康穩定,是提高仔豬成活率及生產效率的有力保障。前人研究大多集中高NSP特別是高INSP日糧對緩解妊娠母豬便秘具有積極的作用[70-71],而對NSP在母豬妊娠階段腸道微生物區系的調控卻罕有報道。研究表明,母豬可通過乳汁和糞便等母源微生物的垂直傳遞,影響仔豬粘膜微生物的定植,且表現出空間生態位點的選擇特異性[72]。另有報道表明,經健康人類糞菌移植后的無菌小鼠,飼喂低NSP日糧后,腸道菌群多樣性顯著降低,此時改用高NSP日糧進行飼喂,可在一定程度上恢復原有的菌群結構。低NSP水平日糧持續飼喂3代后,高NSP日糧對其菌群結構的恢復效應大幅度降低[73]。提示隨著世代間母子傳遞,微生物的冗余特性逐漸降低直至消亡,預示著妊娠期母豬高NSP型飲食干預,在新生仔豬腸道菌群多樣性的早期建立方面具有重要意義。此外,一項來自斯坦福大學的研究表明,抗生素使用后,人體微生物菌群具有很強的自我恢復能力,而不同的抗生素使用后,小鼠腸道菌群的反應具有共性。值得一提的是,低NSP日糧會加重腸道菌群的失衡,延緩環丙沙星使用后的恢復,而高NSP日糧可加快菌群的恢復,提示高NSP日糧對于緩解抗生素對腸道菌群的負面影響具有積極作用[74]。
4 展望
迄今為止,國內外有關日糧NSP在動物上的應用研究多側重于不同來源及濃度NSP的飼喂效果及其對表觀指標的影響。盡管近年來部分研究對日糧NSP與腸道健康的關系進行了探討,但多集中于菌群數量及其代謝產物的調節方面,缺乏宏基因組、宏轉錄組等現代微生物技術研究手段。此外,目前有關NSP調控豬腸道菌群的核心微生物,以及該微生物群在NSP調控腸道健康中的貢獻及機制尚不十分明確。因此,深入認識日糧NSP組分的營養作用,探討豬腸道微生物與NSP及其組分間的對話機制,消除或緩解NSP對機體的不利影響,充分發揮其積極作用,對促進動物健康、提高飼料的有效能值以及緩解我國飼料資源短缺局面大有裨益。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
