魚蝦腸道菌群代謝產物短鏈脂肪酸研究進展
2020-02-22 04:48:24劉宇丁倩雯冉超楊雅麟王安然張洪玲張進雄李解RolfErikOlsenEinarRing張震周志剛
生物技術通報 2020年2期
關鍵詞:生長
劉宇 丁倩雯 冉超 楊雅麟 王安然 張洪玲 張進雄 李解 Rolf Erik Olsen Einar Ring? 張震 周志剛
(1. 中國-挪威魚類消化道微生物聯合實驗室,北京 100081;2. 中國農業科學院飼料研究所 農業部飼料生物技術重點實驗室,北京 100081;3. 挪威-中國魚類消化道微生物聯合實驗室,特隆赫姆,挪威;4. 北京市畜牧業環境監測站,北京 102200)
腸道菌群是指存在于動物腸道中的數量龐大的微生物群體,其中超過99%的都屬于細菌,其個體總數為100兆個,約為體細胞數目的10倍,其編碼基因數目是人類基因組編碼數的100-150 倍[1]。腸道菌群也被稱為腸道微生態系統,作為最龐大、最復雜的微生態系統,定殖于動物腸道內,與宿主共生并互相影響,參與調控宿主營養代謝和免疫抗病等生理過程[2]。近些年來,腸道菌群代謝產物逐漸走進研究者的視野,研究發現腸道菌群代謝產物也會進一步對宿主生理產生重要影響。短鏈脂肪酸(Short-chain fatty acids,SCFAs)作為腸道微生物重要的代謝物質之一,也受到越來越多的關注。
改革開放以來,我國水產養殖行業發展迅速,據統計,2018年我國水產品養殖總產量已經達到5×107t,已經占到我國全年水產品總產量的78%以上,目前我國是世界上唯一一個水產品總產量中養殖量超過捕撈量的主要漁業國家[3]。我國水產養殖業的發展勢頭迅猛,規模化、規范化、集約化的水產養殖除了帶來可觀的經濟效益外,也造成了水產養殖動物的應激加劇,養殖環境日趨惡化,病害發生率越來越高,抗生素濫用等問題。水產養殖過程中濫用的抗生素等化學藥物不僅增加病原菌的耐藥性,破壞水產動物的腸道菌群穩態,也造成養殖環境抗生素超標;同時,殘留在養殖水產動物中的抗生素也會通過食物鏈富集作用進入人體,危害到人類的身體健康[4]。因此,人們對無毒、無副作用、無污染的綠色安全替代物質的需求越來越迫切,微生態制劑近些年逐漸應用于水產養殖行業,具有綠色環保的優點,但微生態制劑在水產養殖應用上也存在不足,包括運輸保存困難,菌體易失活,穩定性差等缺點。為實現水產養殖無抗無憂,筆者所在團隊提出益生元件是水產養殖綠色發展的優勢策略[5-9]。“益生元件”即益生菌效應分子,這些益生菌微生物相關的模式分子和特定結構的代謝產物通過影響微生物與宿主代謝信號傳導途徑,亦或優化土著微生物菌群的結構,調控宿主營養代謝及免疫[10]。SCFAs是腸道菌群發酵產生的重要代謝物質之一,是近年來的研究熱點,對宿主的營養代謝與免疫抗病有重要影響,有望彌補微生態制劑的不足[11]。
1 水產動物消化道短鏈脂肪酸的種類、分布與吸收
SCFAs一般是不易消化的碳水化合物經腸道菌群厭氧發酵產生,包括甲酸、乙酸、丙酸、異丁酸、丁酸、異戊酸和戊酸,其結構圖如表1。據報道,哺乳動物近端結腸SCFAs含量約為120 mol/L,遠端結腸SCFAs含量為90 mol/L,其中乙、丙、丁酸三者比例大致為3∶1∶1,共占總量的95%,是主要的 SCFAs[12]。Mountfort等[13]研究發現,水產動物中主要存在的SCFAs也為乙酸、丙酸,丁酸。海鯉Crinodus lophodon)腸道中乙酸、丙酸、丁酸比例為8∶3∶1,鯡魚(Odax cyanomelas)腸道中乙、丙、丁酸比例為37∶13∶2,鯧魚(O. pullus)腸道中三者比例為20∶4∶1。SCFAs在3種溫水海洋魚類的不同腸道分區比例變化接近,與哺乳動物相同的是,3種魚類中主要SCFAs也為乙酸,在魚類中濃度范圍為20-29 mmol/L,丙酸和丁酸含量較低[14]。不同的是,水產動物腸道中丙酸的比例較哺乳動物有所提高,丁酸含量比哺乳動物低。SCFAs可通過兩種方式進入腸道,包括簡單擴散方式和主動運輸方式,進而對宿主產生調控作用。其中非游離SCFAs通過簡單擴散方式經腸上皮屏障進入腸道細胞,被腸道細胞吸收利用,游離SCFAs經單羧酸轉運蛋白1(Monocarboxylatetransporter1,MCT-1)和鈉耦聯單羧酸轉運蛋白1(Sodium-coupled monocarboxylate transporter1,SMCT-1)這兩種主要的轉運體進入腸道,發揮作用[15]。SCFAs也可通過G蛋白偶聯受體(Gprotein-coupled receptor)發揮生物學效應,G蛋白偶聯受體也稱為游離脂肪酸受體,能夠識別游離脂肪酸,目前已知的SCFAs受體有GPR41、GPR43和GPR109,它們可通過短鏈脂肪酸,調控動物腸道對營養物質的吸收、能量的吸收利用,激素分泌、免疫抗病等多種生物學過程[16]。
2 水產動物消化道短鏈脂肪酸的生理效應
SCFAs主要由乙酸鹽、丙酸鹽和丁酸鹽組成,在調節宿主代謝、免疫系統和細胞增殖方面具有關鍵作用。SCFAs在盲腸和近端結腸中濃度最高,是結腸細胞能量來源(尤其是丁酸鹽),也可以通過門靜脈轉運到外周循環中作用于肝臟和外周組織。盡管外周循環中SCFAs水平較低,但它們作為信號分子參與宿主不同的生物過程,發揮調控作用[17]。
2.1 營養代謝調控效應
SCFAs在腸道中濃度很高,腸上皮細胞通過被動和主動機制將SCFAs吸收至細胞質中,參與宿主能量代謝。哺乳動物中研究表明,SCFAs是腸上皮細胞重要的能量來源,SCFAs氧化后可為結腸黏膜細胞提供近70%的能量[18],對混合SCFAs 產生的二氧化碳的研究表明,細胞利用SCFAs的順序是丁酸、丙酸、乙酸[19]。乙酸作為動物腸道內含量最多的SCFAs,可隨血液進入全身循環,肝可利用乙酸合成谷氨酰胺、長鏈脂肪酸、谷氨酸和β-羥丁酸;乙酸鹽也可通過中樞穩態機制穿過血腦屏障并降低食欲。丙酸可被肝臟吸收,參與糖異生并抑制膽固醇合成,約50%的丙酸在肝內是糖異生的底物。丁酸除了是結腸黏膜上皮細胞的主要能量來源,也參與糖類代謝和脂類代謝。盡管丙酸和丁酸在外周循環的濃度較低,但它們通過激素和神經系統間接影響外周器官[20]。總之,作為腸道菌群產生的一種重要的代謝物質,SCFAs對宿主的營養代謝有著重要的調控作用。
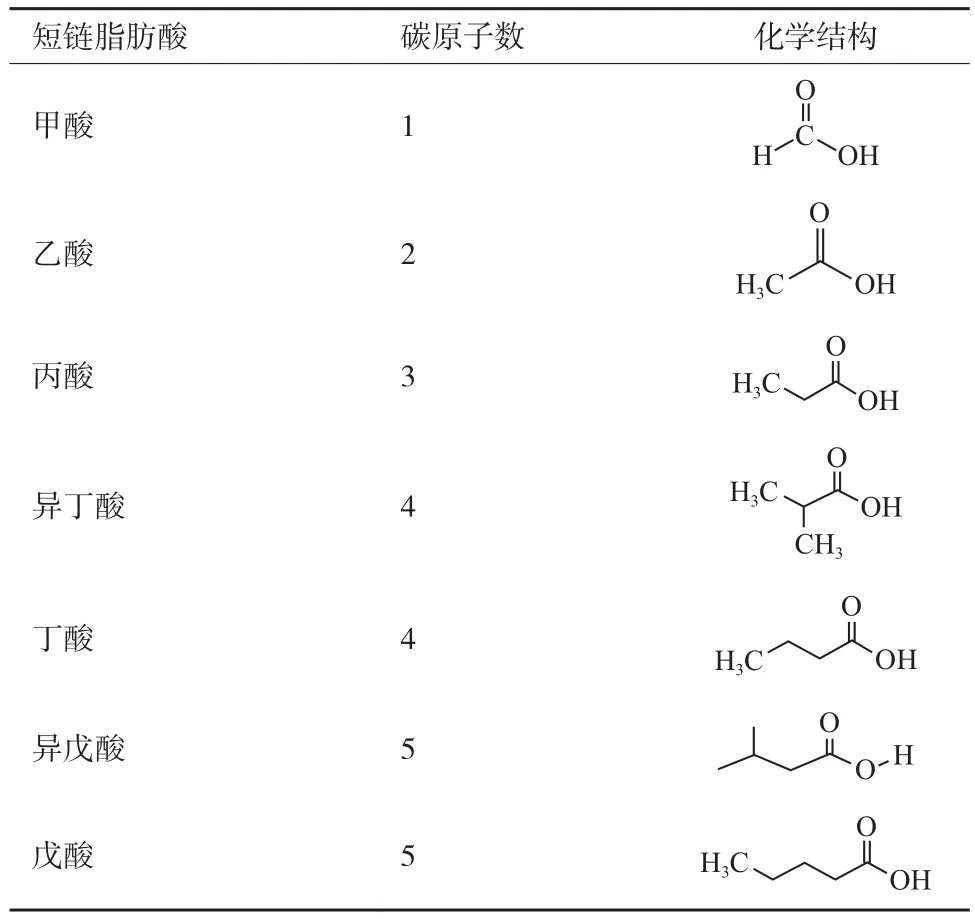
表1 短鏈脂肪酸化學結構式
在水產動物飼料中添加不同種類的SCFAs,也可促進水產動物生長,調控水產動物營養代謝。如乙酸,飼料中添加1 g/kg的乙酸鈉可以顯著提高貝爾湖紅點鮭(Salvelinus alpinus)增重率[21]。如丙酸,在水產動物飼料中添加丙酸鹽,結果發現丙酸鹽在改善水產動物能量吸收方面具有重要作用,據報道,丙酸鈉添加可增加南美白對蝦(Litopenaeus vannamei)對能量和磷的表觀消化系數,改變其腸道微生物并促進南美白對蝦生長[22-23],同時,在羅非魚(Oreochromisspp.)的研究中發現丙酸鹽能提高羅非魚飼料利用效率,蛋白質產值和蛋白質效率等[24],另外丙酸鹽的添加也會提高水產動物生長表現,促進水產動物生長。研究表明丙酸鹽的添加可促進鯉魚(Cyprinus carpioL.)生長,提高鯉魚生長基因表達水平[25],顯著性提高里海白魚(Rutilus frisii kutum)飼料利用效率,促進里海白魚生長等[25],并能提高斑馬魚生長與促進食欲調節相關基因的表達水平,促進其生長[27]。在水產動物飼料中添加乙酸鹽、丙酸鹽能夠促進水產動物對營養物質的吸收利用,促進水產動物的生長。在丁酸鹽的研究中發現,丁酸鹽對不同水產動物生長也有著促進作用,如在海鯛(Sparus aurata)日糧中添加丁酸發現,丁酸會顯著性提高其增重率,通過對海鯛生長的機理進行進一步研究,發現丁酸添加組核苷酸衍生物和幾種常見氨基酸的利用率有所提高[28],同樣地,研究表明丁酸鹽的添加能夠顯著提高商業蝦采食量,改善其生長[29],并顯著提升羅非魚(Oreochromisspp)終末體重、增重率和特定生長率[30]。并且其他形式的丁酸鹽也能提高水產動物生產性能,如 3%(W/W)的PHB(Poly-b-hydroxybutyrate)添加可改善太平洋白蝦的生長性能,腸道消化酶活性及免疫功能,并增加太平洋白蝦腸道SCFAs含量[31]。在藍蟹飼料中添加乙酸鈉、檸檬酸鈉、丁酸鈉或2%丙酸鈉這幾種不同種類的有機酸,均能夠促進藍蟹生長,顯著提高其終末干重和特定生長率,并能夠加速螃蟹蛻皮,同時也能改善藍蟹存活率[32]。
總之,在水產動物飼料中添加3種主要的SCFAs,對提高魚蝦類增重率和促進飼料轉化均具有積極作用,促進其生長。但是也存在與其相悖的研究結果,如北極嘉魚(Salvelinus alpinus)飼料中添加10 g/kg的丙酸鈉,發現丙酸鈉的添加抑制北極嘉魚生長[33]。將2 g/kg丁酸鈉添加到鯰魚(Silurus asotus)飼料中,結果發現丁酸鈉添加組并不能顯著性改善鯰魚生長,并伴隨著鯰魚飼料轉化效率的降低,此外,脫脂大豆飼料中添加丁酸鈉則沒有正向效果[34]。不同的實驗結果可能與試驗動物種類,SCFAs飼喂濃度以及飼喂方式的差異有關。盡管有較多的實驗結果證實SCFAs在介導水產動物營養調控方面起著重要作用,但SCFAs在水產動物中營養調控途徑、調控機理的研究還是相對匱乏因而有待于進一步研究,以便為未來SCFAs在飼料添加中的應用提供理論支撐,進而促進水產養殖行業的健康可持續發展。
2.2 免疫抗病調控效應
在哺乳動物中,SCFAs也可以作為免疫刺激分子,通過激活G蛋白偶聯受體GPR41,GPR43和GPR109A提高淋巴細胞功能,從而發揮免疫相關作用[35]。SCFAs作用于腸上皮細胞、單核吞噬細胞、淋巴細胞等,可以修飾包括基因表達、趨化、分化、增殖和凋亡在內的多種細胞過程,是溝通腸道菌群和腸免疫系統的重要分子[36]。研究發現,生理劑量的乙酸、丙酸、丁酸可對結直腸腫瘤細胞的生長增殖起抑制作用,促進腫瘤細胞的分化,通過多種途徑誘導腫瘤細胞凋亡,起到抗腫瘤作用[37]。目前,SCFAs在臨床上已經開始應用于腸道炎癥性疾病的治療,通過補充食物中膳食纖維的含量,增加腸道內SCFAs的水平,能有效治療潰瘍性結腸炎,控制腹部癥狀。
水產動物腸道致病菌是引起水產動物死亡的主要因素,每年給水產動物養殖造成巨大損失。隨著水產動物SCFAs研究的不斷深入,大量研究者也發現SCFAs對水產動物免疫抗病方面有著重要的調控作用。如甲酸,在太平洋白蝦(Litopenaeus vannamei)飼料中添加甲酸,可以提高太平洋白蝦對副溶血性弧菌的抵抗能力[38]。如乙酸,發現其能夠較好的抑制南美白對蝦(L. vannamei)腸道弧菌屬,改善其腸道健康[39]。如丙酸,斑馬魚(Danio rerio)日糧中添加丙酸鈉能改善黏膜免疫參數[26],提高免疫和抗氧化生長相關基因的表達[40],也能顯著提高里海白魚(Rutilus frisii kutum)的血清溶菌酶和免疫球蛋白水平,并提升補體活性[41],改善鯉魚(Cyprinus carpio)皮膚黏液免疫參數以及血清免疫參數[25,40]。并且丙酸也能夠減少南美白對蝦(L. vannamei)腸道弧菌數目,降低蝦血清凝集系數[42-43]。如丁酸,研究發現鯉魚(C.carpio)飼料中添加丁酸鈉與氧化豆油(Microencapsulated sodium butyrate,MSB),其中丁酸鈉以裝入微型膠囊的形式添加,發現飼喂此飼料會顯著提高促炎因子(IL-β、TNF-ɑ)、抑炎因子(TGF-β)和腸道熱休克蛋白的表達水平,而且丁酸鈉能夠保護腸道免受傷害和修復細胞組織損傷[44]。通過乙酸、丙酸、丁酸在水產動物中的研究可以發現,SCFAs在水產動物中主要表現為對腸道致病菌有一定的抑制能力,并且對于非特異性免疫參數有一定程度的改善,最終改善水產動物的生長。對于其他形式短鏈脂肪酸:如二甲酸鉀和羥基丁酸,研究表明將不同濃度二甲酸鉀添加到羅非魚日糧中,隨著添加劑量的提升羅非魚存活率也顯著提高,而且二甲酸鉀的添加也伴隨著羅非魚攝食量,飼料轉化系數,蛋白質能效比的提高[45-46]。同時,歐洲黑鱸(Dicentrarchus labrax)日糧中添加20%羥基丁酸,使黑鱸腸道微生物組成發生改變[47]。河蟹幼苗飼喂封裝有羥基丁酸的鹵蟲和輪蟲,羥基丁酸的添加使河蟹幼苗存活率、孵化率及滲透脅迫顯著提高[48]。其他形式的SCFAs對水產動物的生長也具有促進作用,未來還需要我們對不同種類的SCFAs進行探索研究,闡明SCFAs免疫抗病的具體機理,以期在生產上實現更廣泛的應用。通過以上實驗發現SCFAs在水產動物中主要通過抑制有害病原菌生長,提高水產動物非特異性免疫能力,從而改善腸道健康,調控水產動物免疫抗病。
3 總結與展望
SCFAs對水產動物生長和免疫抗病有著重要作用,了解SCFAs的具體生理功能以及調控方式對SCFAs的高效利用至關重要。哺乳動物研究表明,SCFAs可作為信號分子發揮作用,SCFAs的受體主要有GPCR41,GPCR43,GPCR109A。G蛋白偶聯受體(G protein coupled receptors,GPCRs)是指與G蛋白偶聯的一大類膜蛋白受體的統稱,有7個跨膜α螺旋,且其肽鏈的C端以及連接第5和第6個跨膜螺旋的胞內環上都有G蛋白(鳥苷酸結合蛋白)的結合位點[49]。GPCRs能夠跨越細胞膜,激活細胞外的信號分子(如蛋白酶、核酸、肽類,生物胺和脂質等),然后觸發細胞內部反應,介導了包括正常和疾病在內的眾多生理反應[50-51]。目前,哺乳動物SCFAs研究相對較多,水產動物的研究主要集中于表型,對其調控機制的研究相對較少,因此,SCFAs的調控機制是未來水產研究上重要方向。
飲食、腸道微生物與SCFAs三者相互聯系。SCFAs調控哺乳動物以及水產動物營養代謝、免疫抗病等生理過程即通過調控動物飲食組成,改變動物腸道內SCFAs含量從而調控魚類和哺乳動物的健康。研究表明膳食纖維的添加可影響動物體內SCFAs含量,盡管不同實驗研究膳食纖維投喂的濃度和實驗動物的飼料組成成分信息不同,但是我們也可以從中得出一些結論。據報道,不同濃度的菊粉添加到小鼠半純化飼料中,結果發現菊粉添加會引起小鼠腸道SFCAs濃度升高,但當菊粉添加量增加到20%時,反而會減少小鼠腸道內SCFAs濃度。與其他類型纖維不同的是,在小鼠飼料中添加菊粉會改變小鼠腸道內不同類型SCFAs比例,促進乙酸向丙酸和丁酸轉化[52-53]。在小鼠日糧中添加微晶纖維素,微晶纖維素和低聚果糖、微晶纖維素和果寡糖,微晶纖維素和纖維寡糖,與基礎飼料組及僅微晶纖維素添加組相比,低聚果糖、果寡糖以及纖維寡糖添加組的小鼠盲腸SCFAs含量以及總SCFAs含量(盲腸以及糞便SCFAs之和)均顯著增加,此外纖維的類型以及纖維的日攝入量同樣會影響SCFAs的含量[52]。小鼠日糧中添加纖維素、果膠、抗性淀粉和大麥外殼等不同類型的不易消化碳水化合物,結果表明纖維素、果膠、抗性淀粉添加組小鼠盲腸和空腸SCFAs含量都顯著提高[54]。因此可以推測,在供給動物富含蛋白質或脂肪的飲食的同時補充膳食纖維會促進有益微生物的生長,改變腸道微生物組成,促進SCFAs的產生,改善動物的生長。但是目前水產動物上相關研究還比較缺乏,因此,這可能是未來SCFAs研究的方向之一,也是推進水產養殖的綠色可持續發展的有效方式。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
