同步分泌高效纖維素酶和木聚糖酶菌株YB的鑒定及其酶學性質研究
2020-02-22 04:48:28楊彬李小波周林區佩渝金小寶
生物技術通報 2020年2期
楊彬 李小波 周林 區佩渝 金小寶
(廣東藥科大學生命科學與生物制藥學院 廣東省生物活性藥物研究重點實驗室,廣州 510006)
木質纖維素主要由纖維素和半纖維素組成[1],因其結構的復雜性,幾乎不能被非反芻動物直接吸收利用。我國是畜牧養殖大國,每年需要大量的動物飼料,而飼料中豐富的木質纖維素卻未能得到有效的利用,降低飼料的營養價值,如果能將其轉化為易吸收能源物質,不僅能提高飼料的營養價值,而且向非反芻動物飼料中引入農業秸稈及副產物變廢為寶可降低飼料的生產成本,對解決我國糧食危機和飼料短缺有著重要的意義[2]。然而商業酶的高成本和活力偏低,造成我國木質纖維素生物轉化的瓶頸[3]。
纖維素酶和木聚糖酶是能夠水解木質纖維素的重要工具酶。纖維素酶能夠將纖維素水解為葡萄糖;木聚糖酶能夠將半纖維素水解成為木寡糖、木糖等小分子化合物,而木寡糖和木糖作為雙歧因子,可以提高動物體內雙歧桿菌的繁殖能力,改善胃腸道微環境,增強免疫力[4]。研究發現半纖維素緊緊地包圍著纖維素,限制了纖維素酶與纖維素的接觸,產生阻塞效應,木聚糖酶在纖維素酶降解纖維素過程中可以起協同作用,促進纖維素酶的水解效率[5]。因此,篩選同步分泌高效纖維素酶和木聚糖酶菌株,可以提高木質纖維素的生物轉化,降低工業生產成本,提高飼料的營養價值。本研究從荔枝林腐敗落葉中分離出一株能同步分泌纖維素酶和木聚糖酶的菌株,對其進行碳源篩選和酶學性質測定,旨為木質纖維素酶菌株的生產應用提供一定理論依據。
1 材料與方法
1.1 材料
1.1.1 樣品與試劑 用于菌株分離的荔枝腐敗落葉取自廣州市番禺區廣州大學城荔枝林。木聚糖(xylan)、D(+)-木糖分析純(AR)和剛果紅指示劑均購買于Maicklin公司。
1.1.2 培養基
1.1.2.1 固體活化培養基[6](g/L) 葡萄糖 20 g、酵母浸粉5 g、CaCl20.2 g、KH2PO40.5 g、維生素B110 mg、MgSO4·7H2O 0.1 g、瓊脂 18 g。
1.1.2.2 剛果紅木質纖維素選擇培養基[7](g/L) 剛果紅3.0 g、CMC-Na 或木聚糖2.0 g、硫酸銨0.5 g、NaCl 0.5 g,瓊脂 18 g。
1.1.2.3 液體產酶培養基[8](g/L) 碳源 20 g、硫酸銨 6 g、酵母浸粉 0.4 g、MgSO4·7H2O 0.3 g、CaCl20.5 g、KH2PO41.0 g、FeSO4·7H2O 5.0 mg、MnSO4·H2O 1.6 mg、ZnSO4·7H2O 1.4 mg、CoCl2·6H2O 2.0 mg、 維生素 B110 mg、0.05% Tween 80。
1.2 方法
1.2.1 木質纖維素降解菌的分離與鑒定 將荔枝腐葉在無菌環境下研碎,稱取1.0 g樣品,加入50 mL的無菌水,振蕩2 h后,梯度稀釋[9],取0.1 mL上清液涂布在剛果紅CMC-Na選擇培養基上,進行纖維素降解菌篩選,于28℃培養48 h,用無菌牙簽挑取產生透明圈的不同形態的菌落,接種到剛果紅木聚糖選擇培養基上,于28℃培養48 h,將具有透明圈的菌落進行液體產酶發酵,通過測定酶活比較各菌株的纖維素酶和木聚糖酶的活力,選擇兩種酶活力均最高的菌株進行下一步研究。菌種的形態學觀察,鑒定參照《真菌鑒定手冊》進行。采用DNA提取試劑盒進行DNA提取。應用真菌核糖體DNA內轉錄間隔區引物ITS1(5'-TCCGTAGGTGAACCTGCGG-3') 和 ITS4(5'-TCCTCGCCTTATTGATATGC-3')為引物進行PCR擴增,PCR產物由北京睿博興科生物技術公司測定,將菌株的ITS-rDNA 序列在NCBI上用Blast程序與已知的相關序列進行比對,并利用Mega 5.0軟件構建系統發育樹[10]。
1.2.2 不同碳源對菌株產酶的影響 用接種環刮取約1 cm2活化瓊脂培養基上目標菌株的孢子,接種到種子液搖瓶中,28℃,120 r/min條件下培養24 h,作為后續試驗的種子液。移取2 mL種子液到碳源分別為麥皮、玉米芯、玉米秸稈、CMC-Na等粉末(過18目篩)的液體產酶培養基中,28℃,120 r/min條件下培養120 h。發酵結束后,先將發酵液過120目濾布,除去菌絲體和大部分顆粒較大的碳源粉末,再4 000 r/min離心10 min,上清液即為粗酶液,纖維素酶活力的測定參考王剛等[11]的方法,木聚糖酶活力的測定參考吳萍等[12]的方法。
1.2.3 發酵時間對菌株產酶的影響 移取2 mL種子液到玉米秸稈液體產酶培養基中,28℃,120 r/min條件下培養,每24 h取樣測定酶活力。
1.2.4 酶的分離純化
1.2.4.1 SDS-PAGE與酶譜分析 粗酶液進行酶譜分析參考李源濤等[13]的方法:電泳前將粗酶液與2×上樣緩沖液混合,沸水浴5 min,低溫電泳,結束后將膠體分為兩部分,含蛋白質分子量標記部分用考馬斯亮藍染色,另一部分用30%異丙醇乙酸緩沖液浸泡45 min(每15 min更換一次),蒸餾水沖洗2次,浸泡在乙酸緩沖溶液中,50℃,孵育20 min,剛果紅染色后,用NaCl溶液脫色,直到出現透明條帶時,用5%(V/V)的乙酸浸泡停止反應[14]。
1.2.4.2 不同飽和度的硫酸銨對酶部分純化的影響 取7只試管,每只試管中加入5 mL粗酶液,依次加入不同質量的硫酸銨,緩慢振蕩溶解,使硫酸銨的飽和度分別為:30%、40%、50%、60%、70%、80%和90%。放入4℃冰箱保存2 h,4 000 r/min離心10 min。棄上清,沉淀用0.5 mL乙酸緩沖液(0.05 mol/L,pH 5.0)溶解,即得到部分純化的粗酶濃縮液,測定相對酶活力。
1.2.4.3 DEAE Sepharose Fast Flow陰離子交換層析 將1.2.4.2中50%-80%硫酸銨沉淀所得酶液經冰水浴透析除鹽(過0.45 μm濾膜),上樣于經 0.02 mol/L、pH 8.0的Tris -HCl緩沖液平衡后的 DEAE Sepharose Fast Flow陰離子交換層析柱(1 cm×70 cm)。先用緩沖液進行洗脫,再用含0.1-0.5 mol/L的 NaCl緩沖液進行梯度洗脫,流速為0.5 mL/min,每6 min收集一管(流動相全程冰水浴)。測定每個蛋白質峰的酶活力,收集有酶活力的洗脫峰,對其進行SDS-PAGE純度檢測。
1.2.4.4 Sephadex G-100凝膠層析 將1.2.4.3中分離不純的目標酶液透析、冷凍干燥、濃縮,過0.45 μm濾膜得清液上樣于0.02 mol/L pH5.0檸檬酸緩沖液平衡后的SephadexG-100層析柱(0.5 cm×30 cm),流速為0.5 mL/min,2 min收集一管(流動相全程冰水浴),測定每個蛋白質峰的酶活力,收集有酶活的洗脫峰,并對其進行純度檢測。
1.2.5 酶學性質測定
1.2.5.1 純化酶的最適反應溫度和pH值 最適反應溫度:將酶液分別置于30、40、50、60和70℃的水浴條件進行酶促反應,以最高酶活力為100%,測定相對酶活力。最適反應pH:用pH值為3.0、4.0、5.0、6.0、7.0和8.0的乙酸緩沖液配制1% CMC-Na或1%木聚糖溶液為底物,在最適溫度下測定相對酶活力。
1.2.5.2 純化酶熱穩定性和酸堿穩定性 熱穩定性:將酶液分別在30、40、50、60和70℃的條件下水浴2 h,快速冷卻至室溫,在最適反應條件下測定相對酶活力。酸堿穩定性:將粗酶液分別置于pH值分別為3.0、4.0、5.0、6.0、7.0和8.0的乙酸緩沖液中室溫放置 2 h,在最適反應條件下測定相對酶活力。
1.2.5.3 玉米芯木聚糖樣品制備及酶解產物薄層分析 采用酸堿處理醇沉法[15]從玉米芯中提取木聚糖,將制備的木聚糖放入60℃烘箱中烘干,研磨成粉末,得到玉米芯木聚糖樣品,用粗酶液降解玉米芯木聚糖樣品,水解產物進行薄層色譜分析[15-16]。
2 結果
2.1 菌株的篩選與鑒定
通過剛果紅CMC-Na選擇培養基,從荔枝腐葉中共獲得5株不同形態的單菌落,分別將其接種在剛果紅木聚糖選擇培養基上,28℃培養48 h,獲得一株生長旺盛透明圈較大的菌株YB。表明菌株YB能分泌纖維素酶和木聚糖酶。菌株YB在剛果紅CMC-Na選擇培養基上的生長狀態(圖1-A),在木聚糖選擇培養基上的生長狀態(圖1-B)。菌株YB在PDA培養基上生長旺盛,菌絲潔白,呈放射束狀生長,中心菌絲變黃出現同心圓(圖1-C),隨后菌絲體都逐漸變黃(圖1-D),到第10 天菌絲體漸漸消失,產生大量綠色孢子(圖1-E)。顯微鏡下觀察菌絲體粗壯有分枝,孢子為卵圓形(圖1-F),根據《中國真菌志》初步判定菌株為木霉屬。目標菌株 PCR擴增片段序列長度為592 bp(圖2)。將菌株的ITS-rDNA基因序列輸入NCBI 基因庫中進行同源性比較,與綠木霉(Trichoderma virens)KX343100的相似度為99%,初步鑒定該菌為一株Trichoderma virens,命名為Trichoderma virensYB(圖3)。
2.2 不同碳源對菌株YB產酶的影響
菌株YB在4種碳源中,以玉米秸稈為碳源時,兩種酶的活力都遠高于其它碳源,木聚糖酶活力為:18 218.36±432.56 U/mL。纖維素酶活力為:220.59±18.73 U/mL。木聚糖酶活力順序:玉米秸稈>麥皮>玉米芯粉>CMC-Na。纖維素酶活力順序:玉米秸稈>玉米芯粉>麥皮>CMC-Na(圖4),因此選擇玉米秸稈為碳源進行下一步試驗。

圖1 菌株YB的形態學觀察

圖2 菌株YB的18S rDNA PCR產物電泳圖
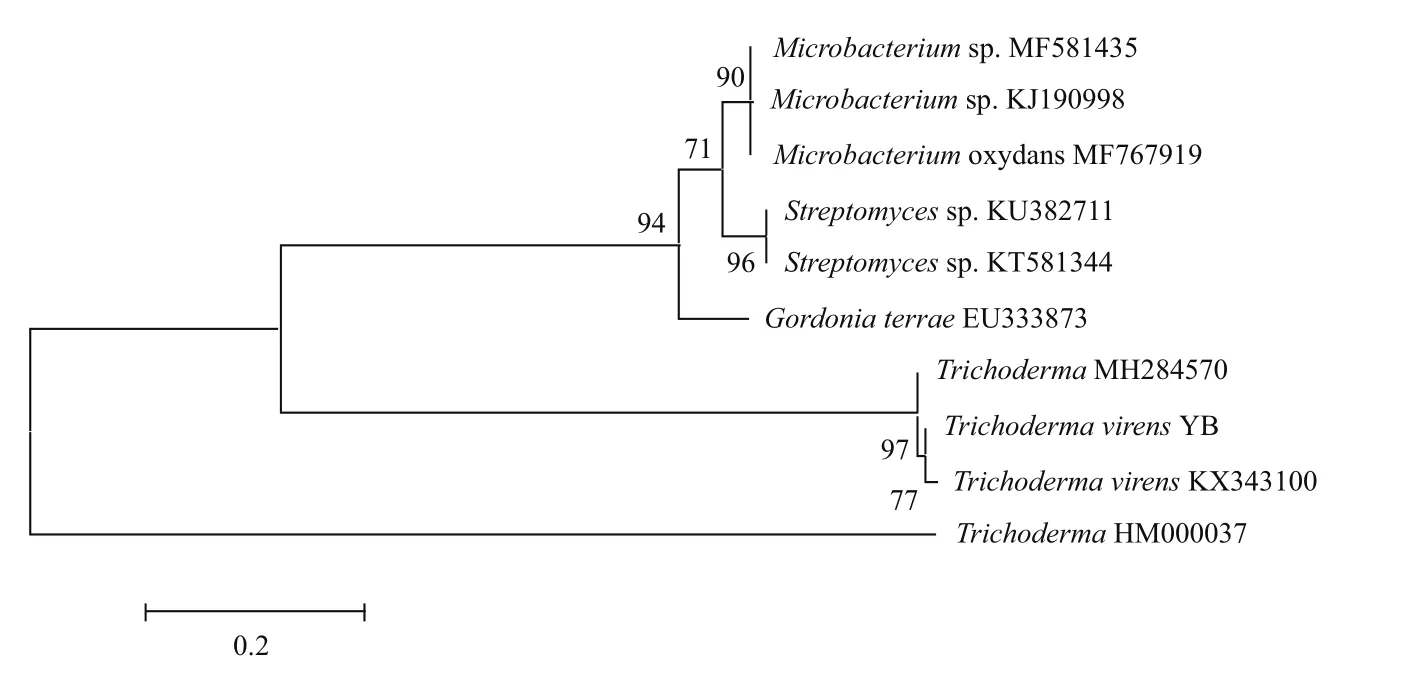
圖3 菌株YB的系統發育樹

圖4 不同碳源對菌株YB產酶的影響
2.3 發酵時間對產酶的影響
在發酵過程中菌株YB在前72 h,纖維素酶活力快速增長,到72 h纖維素酶活力達到最大313.53±26.78 U/mL,72 h后纖維素酶活力逐漸緩慢下降。木聚糖酶活力在前72 h快速增長,72-120 h緩慢增加達到18 120.87±500.37 U/mL,120 h后木聚糖酶活力降低。綜合考慮兩種酶的總酶活力,故選擇發酵周期為72 h(圖5)。

圖5 發酵時間對菌株YB產酶的影響
2.4 酶的分離純化
2.4.1 纖維素酶與木聚糖酶SDS-PAGE及酶譜分析 菌株YB能分泌多種纖維素酶組分,其中有3種為耐熱性質的纖維素酶組分;3種不耐熱纖維素酶組分(圖6-A箭頭指示)。從粗酶液中提取到1種具有耐熱性質的木聚糖酶活力的蛋白帶,未經高溫處理的粗酶液(稀釋10倍),冰浴電泳的條件下有一段非常透明的譜帶(圖6-B)。

圖6 菌株YB發酵上清液的SDS-PAGE與酶譜分析
2.4.2 不同飽和度硫酸銨鹽析對酶活力的影響 硫酸銨鹽的飽和度由30%增加到40%時,濃縮酶液的酶活力均急劇上升,當飽和度為50%時,木聚糖酶活力最大。當飽和度為70%時,纖維素酶活力最大,因此,鹽析木聚糖酶和纖維素酶(CMC酶)的最佳硫酸銨飽和度分別為50%和70%(圖7)。

圖7 不同飽和度硫酸銨鹽析對酶活力的影響
2.4.3 DEAE Sepharose Fast Flow陰離子交換層析由圖8可知,洗脫出5個明顯的蛋白質峰,經酶活力檢測發現,Ⅰ既有CMC酶活力又有木聚糖活力,峰形不對稱,需要進一步分離純化;Ⅱ、III只具有較低的CMC酶活力和酶含量;Ⅳ具有較高CMC酶活力且峰形對稱性較好,表明含雜蛋白少可進行純度鑒定;V不具有酶活性;故只收集Ⅰ和Ⅳ組分,有利于下一步的分離純化。

圖8 粗酶液DEAE Sepharose Fast Flow陰離子交換層析洗脫圖譜
2.4.4 Sephadex G-100凝膠層析 由圖9可知,圖8中Ⅰ組分經Sephadex G-100凝膠層析洗脫出3個比較明顯的蛋白質峰,分別命名為Ⅰa、Ⅰb、Ⅰc;經酶活力檢測發現,Ⅰa只具有木聚糖酶活力;Ⅰb只具有CMC酶活力;Ⅰc未檢測出酶活性,為雜蛋白質。故收集Ⅰa和Ⅰb組分進行蛋白質純度鑒定。
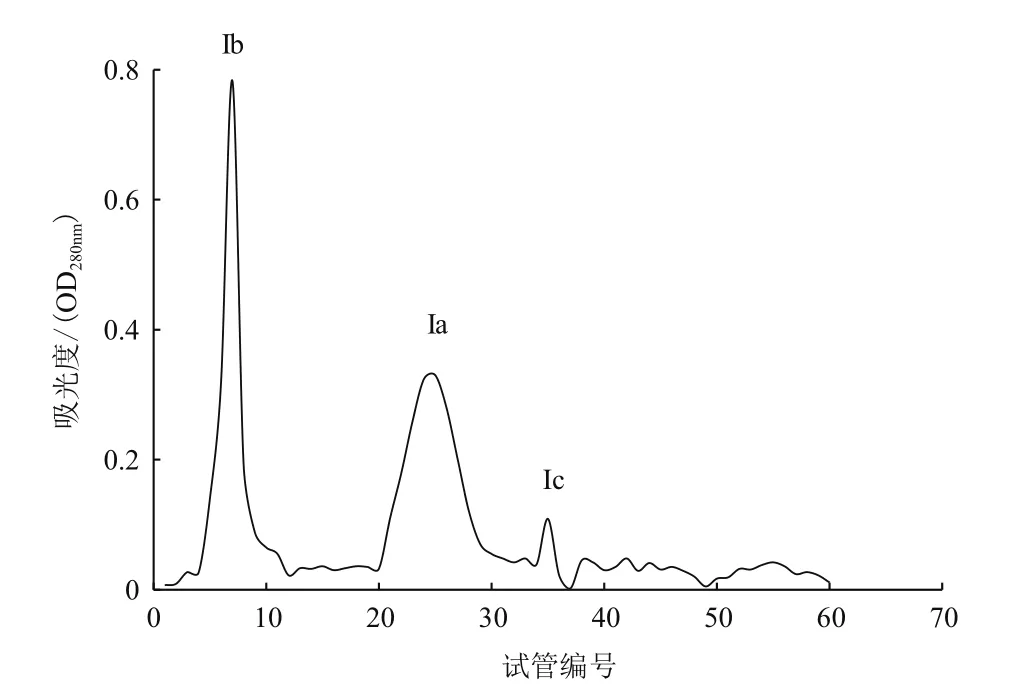
圖9 組分|Sephadex G-100凝膠層析洗脫圖譜
2.4.5 SDS-PAGE電泳鑒定及蛋白質分子量的測定 繪制標準蛋白分子量曲線,計算標準蛋白Marker的相對遷移率,并將其作為橫坐標,其分子量大小的對數為縱坐標,繪制標準曲線得y=-0.848 9x+2.068 3,R2=0.98。圖10為純化后的酶液進行SDS-PAGE電泳鑒定。由電泳結果可知,Ⅳ組分中含有少量的雜蛋白,通過與圖6-A纖維素酶活性電泳對照,可以排除少量雜蛋白的干擾。CMC酶Ⅰb、木聚糖酶Ⅰa均為單一條帶,純度較高。通過標準蛋白分子量曲線計算得到Ⅳ、Ⅰb、Ⅰa分子質量分別為22.8 kD、58.5 kD和44.5 kD。

圖10 目的蛋白酶純化后SDS-PAGE電泳圖
2.4.6 菌株YB纖維素酶和木聚糖酶純化結果 由表1可知,對木質纖維素酶進行3步純化:(NH4)2SO4分級沉淀、DEAE Sepharose Fast Flow、Sephadex G-100凝膠層析,最終CMC酶Ⅳ、Ⅰb、木聚糖酶Ⅰa純化倍數分別為3.65、5.45和11.33倍。回收率分別為 37.45%、10.09%和28.73%。比酶活分別為9.06 U/mg、13.53 U/mg、1595.93 U/mg。其中(NH4)2SO4分級沉淀的純化倍數較低,可能是選擇的沉淀區間較寬造成的。

表1 菌株YB的CMC酶和木聚糖酶的純化結果
2.5 酶學性質研究
2.5.1 酶促反應的最適溫度和pH值 3種酶的最適酶促反應條件均為:50℃,pH 5.0。溫度對酶活力的影響:在30℃時,3種酶的活力較低,隨著溫度的升高,3種酶的活力快速上升,當溫度達到50℃時,3種酶的活力均達到最大,隨著溫度的繼續上升,3種酶的活力均迅速下降(圖11-A)。pH對酶活力的影響:pH小于5.0時隨著pH值的增加,3種酶的活力均逐漸上升,當pH為5.0時,3種酶的活力均達到最大,當pH超過5.0時,3種酶的活力陡然下降(圖11-B)。
2.5.2 酶的熱穩定性和pH穩定性 木聚糖酶Ⅰa與CMC酶Ⅳ在30-50℃較穩定;而CMC酶Ⅰb耐熱性較差,當溫度高于50℃時Ⅰb殘余酶活力快速下降(圖12-A)。Ⅰb在pH4-6范圍內穩定性較強,其酶活力在80%左右;當pH值高于6.0時,Ⅳ其殘余酶活力快速下降,在pH值為8.0時,Ⅳ酶活力大約為15%。而Ⅰa只在pH 5.0有較好的酶活力,當pH值高于或者低于5.0時,Ⅰa酶活力損失較大(圖12-B)。
2.5.3 木聚糖的制備與粗酶液降解木聚糖薄層色譜分析 玉米芯粉經酸堿處理醇沉,獲得大量的玉米芯木聚糖(圖13-A)。菌株YB分泌的木聚糖酶對玉米芯木聚糖水解產物薄層分析。在前0.5 h木聚糖被降解成為木糖和極少的木寡糖,當水解1 h時,產物中逐漸積累不同大小的木寡糖,隨著水解時間的延長,產物中的木寡糖種類也不斷的增加。到第4 h時,木聚糖被水解成了多種木寡糖(圖13-B)。
3 討論
研究發現自然界中存在多種木質纖維素酶的產酶菌,主要有細菌、放線菌和絲狀真菌。其中真菌分泌的木質纖維素酶具有酶活高,酶系全,多為胞外酶利于分離提取等優點,廣泛地用于工業生產。工業生產纖維素酶的主要真菌多為木霉屬、青霉屬、曲霉屬等。在木質纖維素生物轉化的研究中,研究發現木聚糖酶在纖維素酶降解纖維素過程中起協同作用,促進纖維素酶的水解效率,同時木聚糖酶降解半纖維素過程中生成的生物活性小分子化合物(木寡糖),能夠提高動物的免疫力,降低疾病的發生[5]。因此,人們越來越關注于能夠產生多種木質纖維素復合酶的新型微生物來降解木質纖維素。劉新育等[17]以斜臥青霉為出發菌,經紫外誘變獲得了高產纖維素酶(19.71 IU/mL)和木聚糖酶(215.4 IU/mL)的菌株;Jampala等[18]對里氏木霉NCIM 1186同步生產纖維素酶和木聚糖酶發酵條件進行優化(培養周期為5 d,產纖維素酶3 055.65 U/L,產木聚糖酶444.94 U/L);Tian等[19]從15株真菌中篩選到一株能同步高產纖維素酶(28.81 U/g)和木聚糖酶(213 U/g)的黑曲霉菌株,吳萍[12]篩選到一株高產木聚糖酶菌株,其酶活力達到7 088.54 U/mL。倪大偉等[20]篩選到一株高產木聚糖酶的黑曲霉菌株,其酶活為12 664 U/mL。本次試驗從荔枝林腐敗落葉中篩選到一株能同步分泌纖維素酶和木聚糖酶的菌株,經形態學和分子生物學鑒定為Trichoderma virens,命名為Trichoderma virensYB。菌株YB利用玉米秸稈進行液體發酵,培養周期為3 d,其纖維素酶活達到313.53 U/mL,木聚糖酶活高達18 120.87 U/mL。可知菌株YB分泌木聚糖酶的能力較強,屬于高產木質纖維素酶,具有生長快,培養周期短等優點,具有一定的應用前景。
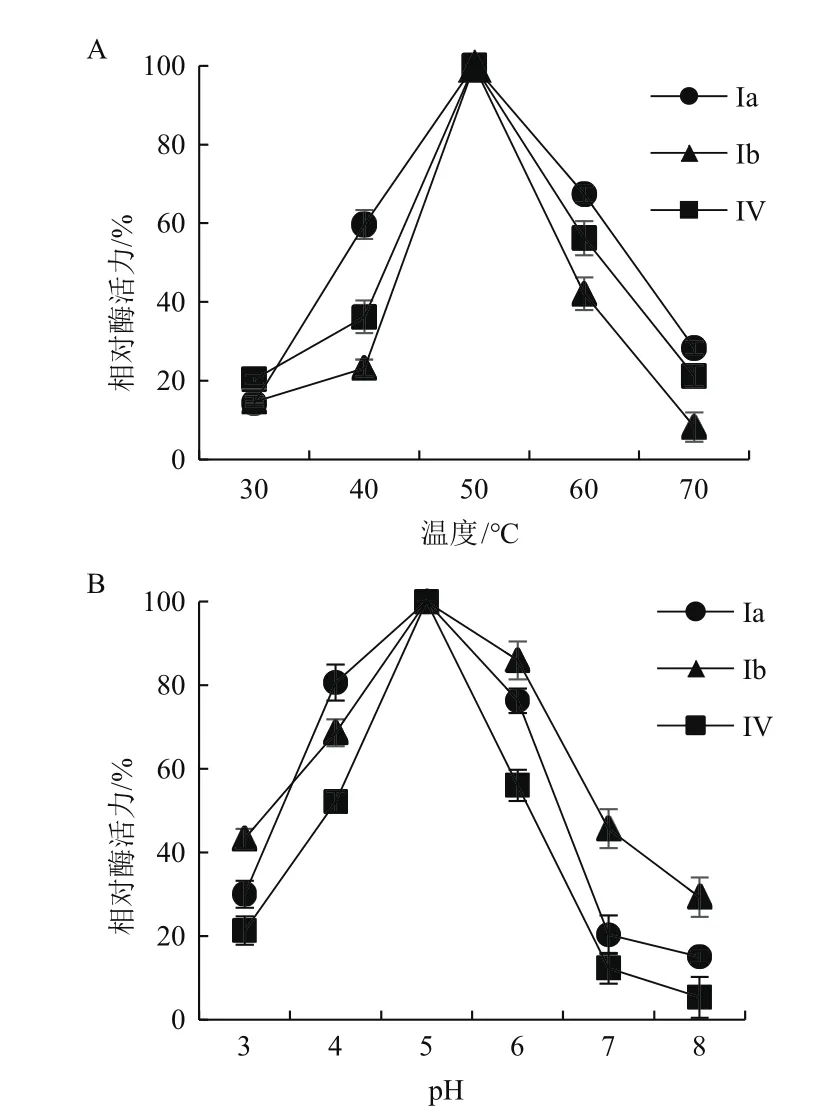
圖11 溫度與pH值對酶促反應酶活力的影響
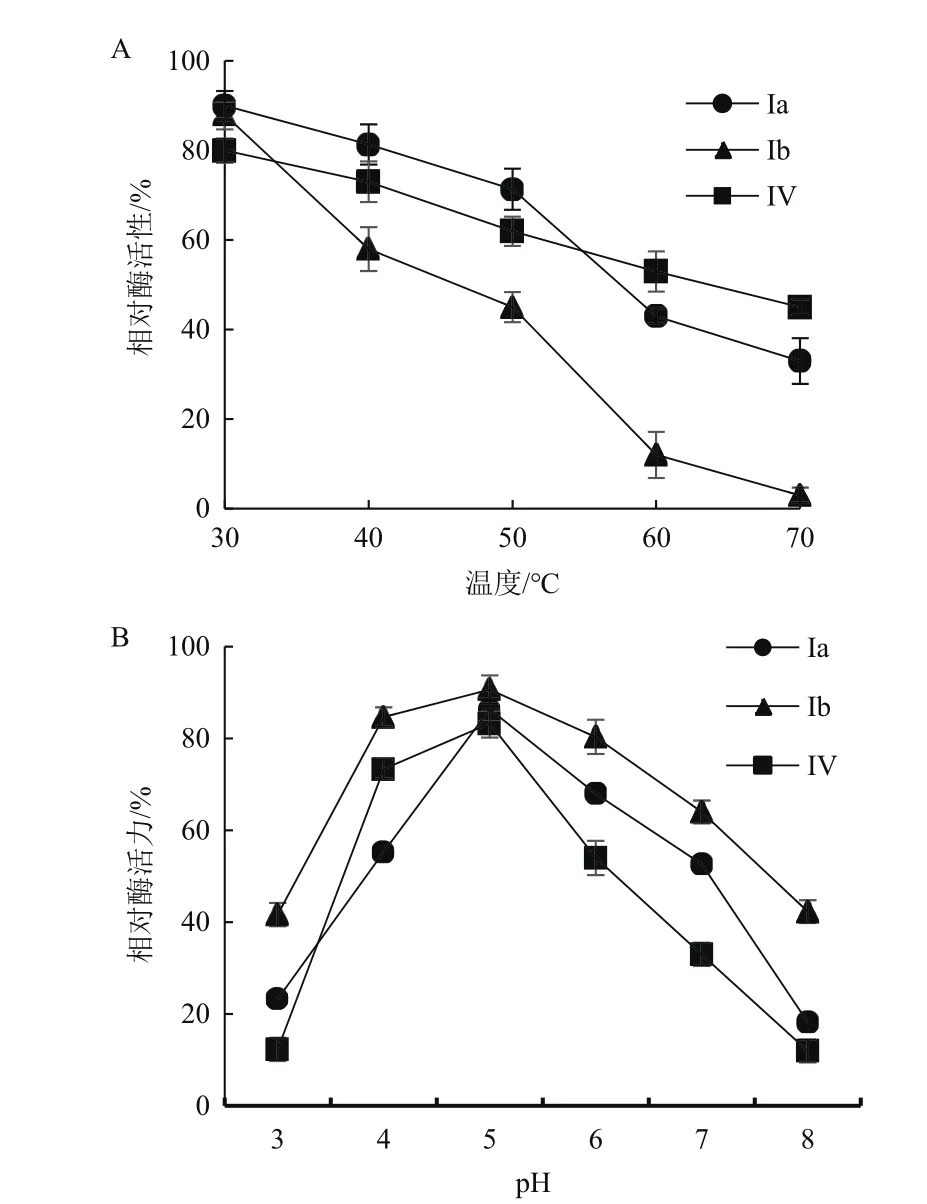
圖12 酶的熱穩定性和pH穩定性

圖13 粗酶液降解木聚糖薄層色譜
碳源篩選結果發現,菌株YB以玉米秸稈為碳源時,兩種酶活力均遠高于其它碳源的酶活。麥皮、玉米芯、CMC-Na為碳源時產生的酶活性偏低,其可能的原是因麥皮中含有大量的淀粉和蛋白質,麥皮既作碳源又作氮源導致菌株YB快速進行營養生長,酶分泌量降低。玉米芯粉中含有的木聚糖含量高達35%[21]遠高于玉米秸稈,一般微生物易于吸收葡萄糖,導致培養基中,木聚糖酶降解木聚糖產生的木糖在發酵液中積累,木糖的積累抑制了木聚糖酶的分泌。CMC-Na為碳源時,培養基經高溫滅菌后,發酵液黏度較大,可能造成發酵液中的菌體缺氧,酶的分泌能力降低。
通過酶譜法可知,菌株YB可以分泌多種纖維素酶,在分離純化過程中,只進行了內切葡聚糖甘酶和木聚糖酶的分離,并未對纖維素外切酶進行分離,最終分離出一種木聚糖酶(Ⅰa)和兩種CMC酶(Ⅰb,Ⅳ)和,其分子量分別為44.5 kD、58.5 kD和22.8 kD;其中分離得到的CMC酶(Ⅰb 58.5 kD,Ⅳ 22.8 kD)分子量與Zhou等[22]從綠色木霉t100-14突變株分離得到的CMC酶(EGⅠ 56.5 kD,EGШ 21 kD)大小相近。酶學性質研究發現,纖維素酶和木聚糖酶在pH5.0時穩定性最強,表明菌株YB分泌的為弱酸性的纖維素酶和木聚糖酶,可以廣泛的運用于飼料生產、果汁和食醋的釀造。菌株YB能夠利用廉價易得的玉米秸稈為碳源,能夠同時分泌酶活性較高的纖維素酶和木聚糖酶,并能將玉米芯木聚糖降解為木糖和多種木寡糖,這對降低工業上生產木質纖維素酶、木寡糖的成本和提高飼料的營養價值有著重大的意義。本實驗只進行了碳源的篩選和酶學性質的研究,并未對其具體的發酵培養條件進行優化,這將是我們下一步要展開的試驗。PAGE 檢測,其分子量分別為58.5 kD、22.8 kD和44.5 kD。3種酶的最適酶促條件均為:pH 5.0,溫度50℃。鹽析木聚糖酶和CMC酶的最佳硫酸銨飽和度分別為50%和70%。菌株YB分泌的木聚糖酶,能將玉米芯木聚糖降解為木糖和多種木寡糖。
4 結論
從荔枝林腐敗落葉中篩選出一株能同步分泌高效纖維素酶和木聚糖酶的綠木霉菌株,命名為Trichoderma virensYB,其最佳產酶碳源為玉米秸稈,酶譜分析可知Trichoderma virensYB能夠分泌高效木聚糖酶和多種纖維素酶組分。用DEAE Sepharose Fast Flow陰離子交換層析和Sephadex G-100凝膠層析分離純化得到CMC酶Ⅰb、Ⅳ和木聚糖酶Ⅰa;純化倍數分別為3.65、5.45和11.33倍。回收率分別為 37.45%、10.09%和28.73%。比酶活分別為9.06 U/mg、13.53 U/mg和 1595.93 U/mg。通過 SDS-
