參附益心方對缺氧原代心肌細胞脂肪酸利用的影響
2020-02-24 07:04:54喬利杰李彬#王新陸謝世陽高原朱明軍王永霞
中國藥房 2020年2期
喬利杰 李彬# 王新陸 謝世陽 高原 朱明軍 王永霞

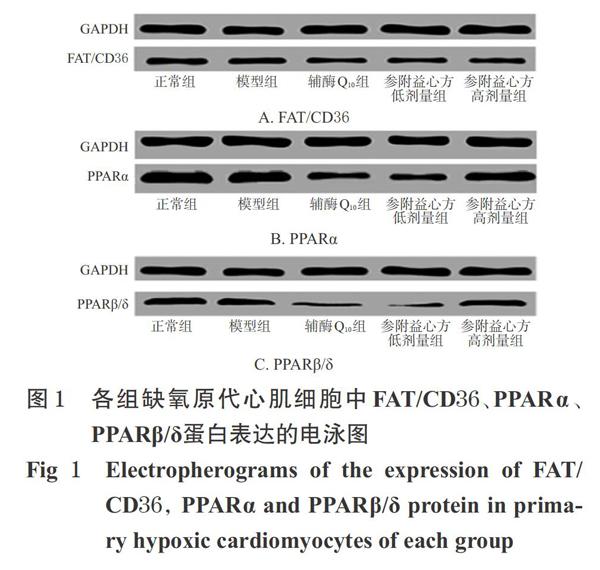
中圖分類號R285.5
文獻標志碼A
文章編號 1001-0408(2020)02-0149-05
DOI l0.6039/j.issn.1001-0408.2020.02.05
摘要 目的:探討參附益心方對缺氧原代心肌細胞脂肪酸利用的影響及其可能機制。方法:取新生l-3d的SD大鼠心尖部組織,進行原代心肌細胞的分離、培養與鑒定,并將細胞隨機分為正常組、模型組、輔酶Q.。組(陽性對照,1X10-4mol/L)和參附益心方低、高劑量組(0.25、0.5 mg/mL)。除正常組外,其余各組細胞均于5%02、5qoCO2、90%N2條件下培養6h以復制缺氧損傷模型。缺氧6h后,采用熒光素酶發光法檢測各組細胞中三磷酸腺苷(ATP)的含量,采用Westem blotting法檢測其脂肪酸轉位酶(FAT/CD36)、過氧化物酶體增殖物激活受體(PPARa、PPARp/8)的表達情況。結果:與正常組比較,模型組細胞中ATP含量以及FAT/CD36蛋白的相對表達量均顯著降低(P<0.05)。與模型組比較,輔酶Q10組和參附益心方高劑量組細胞中ATP含量均顯著升高,而輔酶010組細胞中FAT/CD36、PPARa蛋白,參附益心方高劑量組細胞中FAT/CD36蛋白以及參附益心方各劑量組細胞中PPARa、PPARp/8蛋白的相對表達量均顯著降低(P<0.05)。結論:參附益心方可通過抑制FAT/CD36、PPARa、PPARp/8蛋白的表達來抑制缺氧原代心肌細胞的脂肪酸利用,改善其能量代謝。
關鍵詞 缺氧;原代心肌細胞;脂肪酸;代謝;參附益心方
心力衰竭(以下簡稱“心衰”)是由于心臟收縮或舒張功能異常,導致排血量不能滿足機體正常需求而引發的以氣喘、乏力、水腫為主要表現的一系列臨床綜合征,為多種心臟疾病發展的最終階段。有學者認為,心肌維持正常生理活動所需的能量得不到滿足或代謝失于平衡,可引起心臟結構與功能受損從而導致超負荷心肌損害,進一步引發心衰[1]。由此可見,改善心肌能量代謝對緩解心衰患者的癥狀至關重要。脂肪酸是心肌細胞產生三磷酸腺苷(ATP)的主要底物,有動物實驗及臨床研究證實,抑制脂肪酸利用可改善心肌能量代謝,延緩心衰病程進展[2-3]。
參附益心方為治療心衰的經驗方。有不少研究證明,該方可改善慢性心衰模型大鼠的心功能,降低其血清心房鈉尿肽(ANP)、B型腦鈉肽(BNP)和心肌血管緊張素Ⅱ(AngⅡ)水平,下調心肌細胞及其線粒體中活性氧(ROS)的表達,并上調ATP、肌酸激酶(CK)的含量,提高心肌細胞膜Na+-K+-ATP酶活性,從而達到緩解心衰癥狀的目的[4-6]。但目前有關該方對脂肪酸利用的影響尚未明確。為此,本研究擬建立缺氧原代心肌細胞模型,觀察參附益心方對其脂肪酸利用的影響,并初步探討可能的作用機制,以期為該方治療慢性心衰提供更有力的理論依據。
l 材料
1.1 儀器
BCM-IOOOA型生物超凈工作臺(蘇凈集團蘇州安泰空氣技術有限公司);RC0-3000TVBB型C02培養箱(美國Revco公司);JA203型電子分析天平(上海海康電子儀器廠);TDL-50B型低速臺式大容量離心機(北京醫用離心機廠);CK40型倒置顯微鏡(日本Olympus公司);DMI3006型熒光倒置顯微鏡(德國Leica公司);MK3型酶標儀(美國Thermo Fisher Scientific公司);ChemiDoc XRS+型化學發光成像分析系統(美國Bio-Rad公司)。
1.2 藥品與試劑
參附益心方浸膏干粉(山東步長制藥股份有限公司,批號:131101,規格:1g干粉相當于生藥總量9.5 g);輔酶Q10片[衛材(中國)藥業有限公司,批號:H10930021,規格:10 mg];心肌細胞消化液(含胰蛋白酶0.16 g、氯化鈉1.6 g、碳酸氫鈉0.070 6 g、葡萄糖0.198 2g、氯化鉀0.059 6 g、羥乙基哌嗪乙磺酸0.4 g.ddH20 200mL,批號:T1320)、DMEM高糖培養基(批號:12100)、磷酸鹽緩沖液(PBS,pH 7.4,批號:P1020)、RIPA蛋白裂解液(批號:ROOIO)、蛋白上樣緩沖液(批號:P1015)、CCK-8試劑(批號:CA1210)、BCA蛋白濃度測定試劑盒(批號:PC0020)、ECL發光液(批號:PEOOIO)均購自北京索萊寶科技有限公司;溴脫氧尿核苷(Brdu,批號:B9285)、4 ,6-二脒基-2-苯基吲哚(DAPI,批號:D9542)均購自美國Sigma公司;YOYO-I染料(批號:Y3601)、逆轉錄試劑盒(批號:11752050)、擴增試劑盒(批號:11744500)均購自美國Invitrogen公司;Opti-MEM培養基(批號:31985088)、胰蛋白酶(批號:27250018)均購自美國Gibco公司;兔源心肌肌鈣蛋白I(cTn I)多克隆抗體(批號:BM1765)、SABC-FITC標記山羊抗兔IgG二抗(批號:SA1064)均購自武漢博士德生物工程有限公司;0.4%臺盼藍染色液(華美生物工程有限公司,批號:C0040);ATP檢測試劑盒(瑞士Roche公司,批號:11699695001);兔抗大鼠脂肪酸轉位酶(FATICD36)多克隆抗體(批號:ab64014)、兔抗大鼠過氧化物酶體增殖物激活受體α(PPARa)多克隆抗體(批號:ab24509)、兔抗大鼠PPARβ/δ多克隆抗體(批號:ab23673)均購自艾博抗(上海)貿易有限公司;兔抗大鼠GAPDH多克隆抗體(批號:10494-1-AP)、辣根過氧化物酶標記的山羊抗兔IgG=抗(批號:SA00001-2)均購自武漢三鷹生物技術有限公司;胎牛血清(FBS,以色列Biological Industries公司,批號:04-OO1-1A);其余試劑均為分析純,水為雙蒸水。
1.3 動物
SPF級SD大鼠,新生1~3 d,雌雄不限,體質量約5~6 g,由河南省動物實驗中心提供,動物生產許可證號:SCXK(豫)2015-0005。
2 方法
2.1 藥液制備
2.1.1參附益心方藥液參考本課題組已發表文獻[6],稱取參附益心方浸膏干粉0.25 g,用DMEM高糖培養基10 mL稀釋,經0.2 μm濾膜濾過后,制得質量濃度為25mg/mL(以浸膏干粉質量計)的藥液。需現用現配。2.1.2輔酶Q.。藥液稱取輔酶Q10片50 mg,用DMEM高糖培養基5.8 mL稀釋,經0.2 μm濾膜濾過后,制得濃度為1X10-2mol/L的藥液,備用。
2.2 原代心肌細胞分離、培養與鑒定
于無菌條件下取出乳鼠的心尖部,以4℃的PBS清洗3次,隨后將其剪成約1 mm3大小的組織塊,經含0.08%胰蛋白酶的心肌細胞消化液吹打混勻后,轉移至玻璃瓶內,于37℃水浴中孵育6 min,自然沉淀,棄去上清液;再同法反復消化約7~8次,直到組織塊變成白色透亮狀。取各次自然沉淀后的上清液與等體積含20%FBS的DMEM高糖培養基混勻,以1 000 r/min離心5min,棄去上清液,沉淀與含10% FBS的DMEM高糖培養基(以下簡稱“完全培養基”)1 mL混合,用200目篩網濾過,去除未完全消化的組織塊,得細胞懸液。取上述細胞懸液適量,于37℃、5%C02培養箱(下同)中差速貼壁培養90 min后,吸取細胞懸液,以1 000 r/min離心5min,棄去上清液,沉淀與完全培養基充分混合,制成細胞懸液。取少量該細胞懸液,用0.4%臺盼藍染色液染色,在倒置顯微鏡下計數以計算活細胞比率(染色后,死細胞呈藍色):活細胞比率=活細胞數/(活細胞數+死細胞數)XlOO%。用完全培養基將細胞濃度調整為4x105個/mL,加入濃度為0.1 mmol/L的Brdu溶液中以抑制成纖維細胞增殖,培養48h后,棄去上清液,細胞用不含血清的DMEM高糖培養基同步培養24 h;棄去上清液,用PBS清洗3遍,置于4%多聚甲醛溶液中固定10 min,加入cTnI多克隆抗體(稀釋度為1:50),4℃孵育過夜;加入SABC-FITC標記山羊抗兔IgG二抗(稀釋度為1:100),避光孵育2h;加入DAPI避光染色5min,用水溶性封片劑封孔,于熒光倒置顯微鏡下觀察、拍照。隨機選取6個視野,記錄視野內細胞總數目(N1)和心肌細胞數(n1,即熒光陽性細胞數),計算心肌細胞比例:心肌細胞比例=n1/N1×1OO%。當該比例超過85%,表明原代心肌細胞純度達到試驗要求,可用以進行后續研究[6]。
2.3 分組、造模與給藥
取“2.2”項下原代心肌細胞適量接種于96孔板中,將其隨機分成正常組、模型組、輔酶Q.。組(陽性對照,IX10-4 mol/L,劑量參考本課題組前期CCK-8試驗結果設置)和參附益心方低、高劑量組(0.25、0.5 mg/mL,以浸膏干粉質量計,劑量參考本課題組前期CCK-8試驗結果設置),每組設3個復孔。正常組和模型組加入完全培養基2 mL,各給藥組加入含相應藥物的完全培養基2 mL。除正常組外,其余各組細胞均于5% 02、5%C02、90%N2條件下培養6h(缺氧時間根據本課題組前期YOYO-1染色試驗結果確定),復制缺氧損傷模型;正常組細胞則于37℃、5%C02條件下培養6h。
2.4 缺氧原代心肌細胞中ATP含量檢測
按“2.2”項下方法分離、培養、鑒定原代心肌細胞后,再按“2.3”項下方法分組、給藥、造模。缺氧6h后,吸棄各孔上清液,用常溫PBS清洗2~3次,每次1min,然后采用熒光素酶發光法以酶標儀檢測細胞中ATP的含量,嚴格按照相應試劑盒說明書重復操作3次。
2.5 缺氧原代心肌細胞中FAT/CD36、PPARα、PPARβ/δ蛋白表達水平檢測
按“2.2”項下方法分離、培養、鑒定原代心肌細胞后,再按“2.3”項下方法分組、給藥、造模。缺氧6h后,采用Western blotting法檢測心肌細胞中FAT/CD36、PPARa、PPARp/8蛋白的表達情況。用RIPA蛋白裂解液300 μL反復吹打破碎細胞,于4℃、12 000 r/min離心10 min后,取上清液。采用BCA法測定各組細胞的蛋白濃度后,加入4x蛋白上樣緩沖液適量,于100℃煮沸變性5min。取變性后的蛋白進行SDS-PAGE電泳(先于80 V下電泳20 min,再于120V下電泳lh),濕法轉膜(恒流:250 mA,時間:2h),以5%脫脂奶粉室溫下封閉2h后,加入相應一抗( FATICD36、PPARa、PPARp/8、GAPDH,稀釋度分別為1:1 000、1:1 000、1:1 000、1:4 000),4 cC孵育過夜;然后加入相應二抗(稀釋度為1:5 000),室溫孵育2h后,經ECL發光液孵育后于化學發光成像系統上顯影并拍照。采用Image Pro Plus 6.O軟件分析各蛋白條帶的灰度值,以目的蛋白與內參GAPDH條帶的灰度值比值來表示蛋白的相對表達量。上述試驗重復3次。
2.6 統計學方法
采用SPSS 19.O軟件對數據進行統計分析。計量資料以x±s表示,組間比較采用單因素方差分析或f檢驗。P<0.05為差異有統計學意義。
3 結果
3.1 原代心肌細胞的鑒定
顯微鏡下可見,原代心肌細胞呈星形或者梭形生長,相互接觸交織成網,且呈現同步搏動,搏動頻率、節律、強度穩定,頻率為70~130次/min。原代心肌鑒定結果見本課題組已發表文獻[6]。
3.2 參附益心方對缺氧原代心肌細胞ATP含量的影響
與正常組比較,模型組細胞ATP含量顯著降低(P<0.05);與模型組比較,輔酶Q10組和參附益心方高劑量組細胞ATP含量均顯著升高(P<0.05),詳見表1。
3.3 參附益心方對缺氧原代心肌細胞中FAT/CD36、PPARα、PPARβ/δ蛋白表達的影響
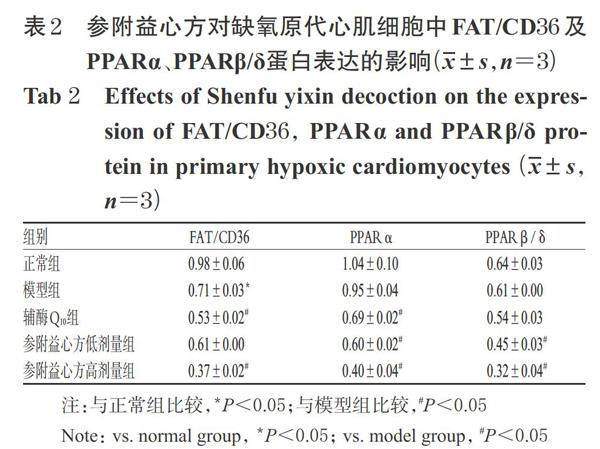
與正常組比較,模型組細胞中FAT/CD36蛋白的相對表達量顯著降低(P<0.05);而PPARa、PPARβ/δ蛋白的相對表達量與正常組比較差異均無統計學意義(P>0.05)。與模型組比較,輔酶Q.。組細胞中FAT/CD36、PPARa蛋白,參附益心方高劑量組細胞中FAT/CD36蛋白以及參附益心方各劑量組細胞中PPARα、PPARβ/δ蛋白的相對表達量均顯著降低(P<0.05),詳見圖1、表2。
4 討論
正常生理條件下,心臟運動所需的ATP約有60%~80%來自于脂肪酸代謝,20%~40%來自于碳水化合物(葡萄糖、乳酸、酮體)代謝;在消耗等量氧氣的情況下,脂肪酸代謝途徑比碳水化合物(葡萄糖)代謝途徑產生的ATP少[7]。可見,在心衰發生時,心肌細胞缺血缺氧能量生成不足,而抑制脂肪酸代謝、增加葡萄糖氧化供能將更有助于心肌獲益。
ATP是心臟可直接利用的能量形式,對維持心肌細胞正常代謝和功能具有重要意義;若ATP生產不足,將導致心肌收縮和舒張功能減弱[8]。輔酶Q.。是線粒體氧化磷酸化的輔酶之一,對線粒體氧化磷酸化和細胞ATP的形成具有重要作用;同時,其可增加心輸出量,降低外周阻力,是治療心衰的常用藥物[9]。因此,本研究選擇輔酶Q10作為陽性對照。
FAT/CD36作為脂肪酸轉位酶,存在于細胞內和細胞膜上,參與脂肪酸的跨膜轉運,并影響機體對脂肪酸的利用[10]。有研究證實,FAT/CD36編碼基因缺失大鼠的脂肪酸攝取率明顯降低,而葡萄糖利用度明顯升高[11]。由此可見,可通過抑制心衰模型大鼠FAT/CD36蛋白的表達來抑制心肌細胞對脂肪酸的利用。PPAR家族為過氧化氫酶增殖物激動受體,其生理功能主要涉及脂肪酸代謝、糖代謝、細胞增殖與分化等[12]。其中,PPARa為機體調控脂肪酸代謝的重要因子,可參與脂肪酸B氧化過程,活化的PPARa可提高肉毒堿棕櫚酰轉移酶I(CPT-I)編碼基因的表達,后者可促使脂肪酸通過線粒體內膜轉移至線粒體基質內,然后被機體代謝[13]。Morgan EE等[14]研究發現,心衰患者心肌組織脂肪酸氧化率下調與PPARa蛋白表達減少有關。PPARβ/δ可通過調控CPT-I、過氧化物酶體增殖物激活受體y輔助活化因子la(PGC-1a)等編碼基因的表達,進一步調節脂肪酸B氧化過程,其反應強度與機體脂肪酸利用呈正比[15]。有研究證實,心衰患者心肌PPARβ/δ蛋白表達下降,脂肪酸氧化將受到抑制,而葡萄糖有氧代謝則會增加[16]。由此可見,抑制PPARa、PPARβ/δ蛋白及其編碼基因的表達對延緩心衰患者的疾病進展至關重要。
中醫認為,心衰多因外感風寒、內傷飲食、情志失暢、溫熱疫毒日久導致氣血虧虛、心脈失養而引起,病位在心,涉及肺、脾、腎三臟,病機總屬本虛標實、虛實夾雜,本虛為氣虛、陽虛,標實為痰飲、血瘀、水停,治法宜益氣溫陽、活血利水[17]。河南中醫藥大學孫建芝教授以此法立方,得參附益心方。方中以人參大補元氣為君,使氣旺血行,水飲痰濕無以停聚;附子、桂枝為臣,溫腎陽、通心脈,使元陽充盛正氣存內;丹參、赤芍、益母草活血化瘀利水,豬苓、澤瀉、車前子利水滲濕,諸藥配伍使瘀祛水行;葶藶子瀉肺平喘、利水消腫,砂仁、大棗溫補中焦,保護胃氣,為佐使之藥。全方補瀉兼施,標本兼治,扶正不滯邪,利水不傷陰[18]。
本研究選用缺氧細胞模型,模擬心衰時心肌細胞的缺氧狀態,從ATP含量、脂肪酸利用相關因子FAT/CD36以及PPARa、PPARp/8蛋白表達等方面來探討參附益心方對缺氧原代心肌細胞脂肪酸利用的影響及可能機制。結果顯示,模型組細胞中ATP含量及FAT/CD36蛋白的表達均較正常組顯著降低;PPARa、PPARβ/δ蛋白的表達雖有降低,但差異無統計學意義。這提示心衰發生時,心肌細胞的能量代謝底物發生了變化,由優先利用脂肪酸轉變為優先利用葡萄糖[7],脂肪酸氧化率降低,產能總體減少。與模型組比較,輔酶Q10組和參附益心方高劑量組細胞中ATP含量均顯著升高,輔酶Q10組細胞中FAT/CD36、PPARa蛋白,參附益心方高劑量組細胞中FAT/CD36蛋白以及參附益心方各劑量組細胞中PPARα、PPARβ/δ蛋白的相對表達量均顯著降低。這提示參附益心方可進一步抑制缺氧條件下原代心肌細胞的脂肪酸利用,改善其能量供應。
綜上所述,參附益心方可通過抑制FAT/CD36及PPARa、PPARβ/δ蛋白的表達來抑制缺氧原代心肌細胞的脂肪酸利用,改善其能量代謝,延緩心衰過程。由于本研究的缺氧時間設定為6h,處于缺氧早期,而參附益心方對缺氧中晚期原代心肌細胞脂肪酸利用的抑制作用尚未可知,故本課題組后續將延長缺氧時間來檢測FAT/CD36蛋白、PPARa、PPARβ/δ蛋白及其mRNA的表達,對上述結論進行驗證;此外,參附益心方對脂肪酸利用其他相關因子的調控作用及機制,以及對葡萄糖利用的調控作用尚未明確,仍有待進一步深入研究。
參考文獻
[1] DOENST T.NGUYEN TD. ABEL ED. Cardiac metabo-lism in heart failure: implications beyond ATP produc-tion[J]. Circ Res,2013 ,113(6): 709-724.
[2]鄒亞興,吳文宇,羅莉,等.苓桂術甘湯對多柔比星所致心力衰竭大鼠的改善作用及其機制[J].中國中醫基礎醫學雜志,2018,24(7):912-915、954.
[3]
FRAGASSO G.PALLOSHI A.PUCETTI P,et al.A ran-domized clinical trial of trimetazidine,a partial free fattyacid oxidation inhibitor, in patients with heart failure[J]J,Am Coll Cardiol,2006,48(5):992-998.
[4]王永霞,任紅杰,朱明軍,等.參附益心顆粒對慢性心力衰竭大鼠血清心鈉素、腦鈉素的影響[J].中國實驗方劑學雜志,2011,17(4):118-120.
[5]王永霞,任紅杰,李彬,等,參附益心顆粒對慢性心力衰竭大鼠心肌血管緊張素Ⅱ1型受體和2型受體表達的影響[J].中華中醫藥雜志,2013,28(9):2710-2712.
[6]李彬,王新陸,謝世陽,等.參附益心方對缺氧原代心肌細胞活性氧和能量代謝的影響[J].中國藥房,2019,30(8):1018-1024.
[7]陶志強,高想,姜衛東,等.曲美他嗪對心衰大鼠心臟功能和脂肪酸代謝的影響[J],中國老年學雜志,2014,34(17):4913-4915.
[8]鄧潔,倪晶宇,黃鈺婷,等,心力衰竭與心肌能量代謝關系的研究進展[J].中國臨床藥理學雜志,2019,35 (11):1213-1216.
[9]李玲巧.瑞舒伐他汀聯合輔酶Q10治療對慢性心衰心功能的影響[J/CD].臨床醫藥文獻電子雜志,2018,5(A1):89-90.
[10]薄俊霞,李敬達,劉慶平.脂肪酸移位酶FAT/CD36與長鏈脂肪酸的結合機制[J].中國動脈硬化雜志,2018,26
[11]張治宇,李振東,殷仁富.脂肪酸轉運體FAT/CD36在心肌脂肪酸代謝中的作用[J].同濟大學學報(醫學版),2006,27(S):25-27.
[12]張云,何秋霞,侯海榮,等.過氧化物酶體增殖物激活受體α的研究進展[J].現代生物醫學進展,2013,13 (29):5798-5800.
[13]胡鳴旭,參芪益心方對H9C2心肌細胞能量代謝的影響及機制研究[D].哈爾濱:黑龍江中醫藥大學,2014.
[14]MORGAN EE, CHANDLER MP, YOUNG ME, et al.Dissociation between gene and protein expression of meta-bolic enzymes in a rodent model of heart failure[J]. Eurl,Heart Fail,2006,8(7):687-693.
[15]胡承,賈偉平.PPAR8與脂代謝及胰島素抵抗[J].國際內分泌代謝雜志,2006,26(6):385-387.
[16]CHENG L, DING G, QIN Q, et al.Cardiomyocyte-re-stricted peroxisome proliferator-activated receptor-deltadeletion perturbs myocardial fatty acid oxidation andleads to cardiomyopathy[J]. Nat Med, 2004, 10 (11):1245-1250.
[17鄧悅,中醫藥在診治心力衰竭中的地位[J].中國中西醫結合雜志,2017,37(10):1162-1163.
[18]謝世陽,王幼平,王小曉,等.參附益心顆粒對心力衰竭大鼠心肌ATP含量及解偶聯蛋白2的影響[J].中華中醫藥雜志,2017,32(10):4412-4415.
(收稿日期:2019-05-08修回日期:2019-09-02)
(編輯:張元媛)
△基金項目:國家自然科學基金資助項目( No.81503419,No.813 73853, No.81603466, No.81603432)
*碩士研究生。研究方向:中醫藥防治心血管疾病。E-mail:qlj000826@163.com
#通信作者:主治醫師。研究方向:中西醫結合治療心血管疾病的基礎及臨床。電話:0371-66264771。E-mail:libinnvhai@163.com
