敲減cGAS基因對人食管鱗癌細胞TE-1增殖、遷移和凋亡的影響
2020-03-02 09:13:52王云帆戴東方傅聰陳德玉
江蘇大學學報(醫學版) 2020年1期
關鍵詞:檢測
王云帆,戴東方,傅聰,陳德玉
(1.江蘇大學醫學院,江蘇 鎮江 212013;2.江蘇大學附屬醫院放療科,江蘇 鎮江 212001)
環鳥苷酸-腺苷酸合成酶(cyclic guanosine monophosphate-adenosine monophosphate synthase,cGAS)是一種DNA傳感器,在DNA識別領域有重要意義[1]。cGAS能夠識別異常存在于細胞質內的雙鏈DNA,催化生成環鳥腺苷酸(cyclic GMP-AMP, cGAMP),隨后結合并激活STING蛋白,通過干擾素調節因子3(IRF3)和TANK結合激酶1(TBK1)促進Ⅰ型干擾素(IFN)表達發揮抗病毒作用[2]。越來越多的證據表明cGAS在抗病毒免疫、腫瘤發生、自身免疫性疾病中發揮了重要作用[3-5]。但對于實體腫瘤中cGAS如何影響腫瘤生物學行為及分子機制卻少有研究。
食管癌是世界范圍內最常見的腫瘤之一,在我國發病人數超過了全世界的50%[6]。我國90%以上的食管癌病理類型是食管鱗狀細胞癌(esophageal squamous cell carcinoma,ESCC)[7]。雖然手術、放療、化療等三大主要治療方法都在進步,但食管癌的5年生存率仍不高[8]。cGAS既是DNA受體,也是一種酶類,它可以與藥物小分子特異性結合,也許能成為抗腫瘤藥物的作用靶點。本研究利用免疫組化分析了60例食管鱗癌患者的癌組織與癌旁組織中cGAS的表達情況;使用慢病毒轉染食管鱗癌細胞株TE-1,構建穩定敲減cGAS基因的TE-1細胞系,探索敲減cGAS對細胞增殖、遷移和凋亡等生物學行為的影響。
1 材料與方法
1.1 材料
1.1.1 組織標本 收集2018年7月至12月在江蘇大學附屬醫院胸外科手術切除且經病理證實的60例食管鱗癌標本,患者均簽署知情同意書。所有入選患者術前均未行抗癌治療。取癌組織與癌旁組織進行甲醛固定及石蠟包埋切片。
1.1.2 質粒與細胞 包裝慢病毒的質粒pLKO.1 TRC 克隆載體、pMD2.G和psPAX2,HEK-293T、Het-1A、TE-1、KYSE-150細胞由本實驗室保存。HEK-293T細胞用含有10%胎牛血清(Gibco公司)的 DMEM 培養基(Gibco公司)培養;Het-1A、TE-1、KYSE-150細胞用含10%胎牛血清的1640培養基(Gibco公司)培養。
1.1.3 主要試劑與儀器 cGAS、β-肌動蛋白、HRP標記的山羊抗兔 IgG抗體購自CST公司;CCK8試劑購自日本同仁公司;凋亡檢測試劑盒購自福麥斯生物技術有限公司;質粒小提試劑盒購自天根生化科技有限公司。
1.2 方法
1.2.1 免疫組織化學染色觀察食管鱗癌組織中cGAS表達 將石蠟包埋的食管鱗癌組織切片常規脫蠟,水化,抗原修復,3%過氧化氫孵育培養,加入1 ∶250稀釋的cGAS一抗4 ℃孵育過夜。陰性對照用磷酸鹽緩沖液(PBS)代替一抗孵育。HRP山羊抗兔二抗室溫孵育后DAB顯色,蘇木素復染,脫水后封片。兩名病理醫師在顯微鏡下隨機挑選5個視野(×200倍)觀察,cGAS表達強度評分如下,① 陽性染色細胞百分比:陽性細胞的比例<5%為0分,5%~20%為1分,21%~50%為2分,51%~75%為3分,76%~100%為4分;② 染色強度:無顯色為0分,淺棕色為1分,棕色為 2 分,深棕色為 3 分。若兩項評分的乘積≥6分則定義為cGAS高表達,若兩項評分的乘積<6分則定義為低表達。
1.2.2 實時熒光定量PCR檢測cGAS mRNA表達水平 提取Het-1A、TE-1、KYSE-150細胞以及cGAS敲減和空載對照TE-1細胞總RNA,測RNA樣品濃度和純度,使得D(260 nm/280 nm)在1.8 ~ 2.0間。反轉錄得到cDNA。引物序列如下:cGAS上游引物5′-CACGAAGCCAAGACCTCCG-3′,下游引物5′-GTCGCACTTCAGTCTGAGCA-3′;β-肌動蛋白上游引物5′-ACTCTTCCAGCCTTCCTTCC-3′,下游引物5′-CGTACAGGTCTTTGCGGATG-3′。反應條件:95℃ 2 min預變性;95 ℃ 15 s 變性,60 ℃ 20 s 退火/延伸,40個循環。采用2-△△Ct法計算cGAS的mRNA相對表達量。
1.2.3 蛋白質印跡檢測食管鱗癌細胞cGAS蛋白的表達 分別取Het-1A、TE-1、KYSE-150細胞,以及cGAS敲減和空載對照TE-1細胞各2×106個,用RIPA裂解細胞,提取蛋白質,BCA試劑盒蛋白定量。SDS-PAGE電泳后,恒流轉膜2 h,脫脂奶粉封閉1 h,4 ℃ cGAS抗體(1 ∶1000)孵育過夜,TBST洗膜3次,每次10 min。HRP標記的山羊抗兔二抗(1 ∶5 000)室溫孵育1 h,TBST洗膜3次,每次10 min。蛋白顯像并重復3次實驗。
1.2.4 慢病毒包裝及感染TE-1細胞 設計cGAS基因的敲減靶序列1:CTTTGATAACTGCGTGACATA,靶序列2:GATGCTGTCAAAGTTTAGGAA,將對照組空載質粒pLKO.1 TRC克隆載體、cGAS敲減的質粒與pMD2.G和psPAX2共轉染至HEK-293T,36 h后收集上清病毒液。取1×106個處于對數生長期的TE-1細胞種于6孔板中,貼壁后血清培養基1.75 mL、病毒液750 μL和聚凝胺20 μg混合感染TE-1細胞,10 h后換正常培養基。感染48 h后,以流式分選將GFP陽性細胞收集后培養。使用熒光顯微鏡觀察細胞轉染陽性率。分別命名穩定敲減TE-1細胞為sh1-cGAS、sh2-cGAS。
1.2.5 CCK8檢測敲減cGAS后TE-1細胞的增殖 取穩定敲減和對照細胞消化計數,按每孔5 000個細胞接種在96孔板中,每組設3個復孔,于24、48和72 h檢測。檢測前每孔加10 μL CCK8,37 ℃、5% CO2培養箱中4 h后酶標儀450 nm測定光密度,繪制生長曲線。
1.2.6 克隆形成實驗檢測敲減cGAS后TE-1細胞的增殖 取穩定敲減和對照細胞按每孔300個細胞接種在6孔板中,每組設3個復孔。完全培養基連續培養14 d,定期換液后,棄培養基,用甲醇固定15 min,隨后結晶紫染色15 min,用PBS輕輕漂洗3次。拍照后用Image J軟件計克隆數分析。
1.2.7 細胞劃痕實驗檢測敲減cGAS后TE-1細胞的遷移 將穩定敲減和對照細胞接種至24孔板,取5×105個細胞均勻鋪入,人工劃筆直的劃痕約2 mm,劃痕后0 h、24 h拍照,重復3次實驗。
1.2.8 Transwell實驗檢測敲減cGAS后TE-1細胞的遷移 無血清培養基重懸穩定敲減和對照細胞。調整細胞密度至2×105/mL。500 μL 含10%胎牛血清的完全培養基加入小室下部,小室上部加入100 μL細胞懸液,細胞培養箱中培養24 h。取出小室,用棉簽擦去殘留在上室內的細胞,甲醇固定細胞15 min,0.1%結晶紫染色20 min,鏡下隨機選取5個視野,計數穿膜細胞數。
1.2.9 流式細胞術檢測敲減cGAS后TE-1細胞的凋亡 將穩定敲減和對照細胞種入6孔板,每孔5×105個細胞,每組細胞設置3個復孔。過夜貼壁24 h使用Annexin V/PI雙染后進行流式檢測,使用Flowjo分析數據。
1.3 統計學分析
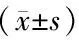
2 結果
2.1 cGAS在食管鱗癌組織中的表達
免疫組化結果顯示,cGAS主要在食管鱗癌的細胞胞質中表達,呈現為棕黃色顆粒。在食管鱗癌組織中,有39例(65%)cGAS呈現高表達;21例(35%)cGAS呈現低表達。在癌旁組織中,cGAS均呈現為低表達甚至陰性。見圖1。

圖1 cGAS在食管鱗癌組織中的表達(免疫組化×200倍)
2.2 cGAS在食管鱗癌細胞中的表達
實時定量PCR結果顯示,食管鱗癌細胞系TE-1、KYSE-150中cGAS mRNA的表達明顯高于食管正常上皮細胞Het-1A(t值分別為10.35、6.86,P<0.01,圖2A);蛋白質印跡結果顯示TE-1、KYSE-150細胞cGAS蛋白表達高于Het-1A(圖2B)。

A:熒光定量PCR;B:蛋白質印跡;*:P<0.01,與Het-1A細胞比較
圖2 cGAS在食管正常上皮細胞和食管鱗癌細胞系中的表達
2.3 重組慢病毒質粒細胞感染及cGAS敲減的鑒定
選取cGAS表達量高的細胞系TE-1,將對照空載體病毒和敲減cGAS病毒分別感染TE-1細胞后72 h,流式分選收集GFP陽性細胞培養,傳代3次,在熒光顯微鏡觀察,可見各組細胞GFP陽性率均在90%以上。熒光定量PCR結果顯示,sh1-cGAS、sh2-cGAS細胞cGAS mRNA均明顯降低(t分別為6.46、5.96,P<0.01)。蛋白質印跡結果亦表明敲減后TE-1細胞cGAS蛋白表達降低。結果表明成功構建了穩定的sh1-cGAS、sh2-cGAS細胞系。見圖3。

A:熒光轉染效率(×200倍);B:熒光定量PCR;C:蛋白質印跡;*:P<0.01,與空載體對照組比較
圖3 驗證TE-1細胞中cGAS敲減效果
2.4 敲減cGAS對TE-1細胞增殖能力的影響
取食管鱗癌細胞TE-1穩轉細胞系sh1-cGAS、sh2-cGAS,CCK8增殖實驗顯示,敲減組細胞增殖較對照組顯著抑制(P<0.01,圖4)。克隆形成實驗表明,sh1-cGAS、sh2-cGAS組克隆形成數目較對照組明顯降低(t分別為27.02、21.19,P<0.01),見圖5。

*:P <0.01,與空載體對照組比較

*:P<0.01,與空載體對照組比較
2.5 敲減cGAS對TE-1細胞遷移能力的影響
細胞劃痕和Transwell遷移實驗結果顯示,sh1-cGAS、sh2-cGAS組傷痕愈合率明顯降低(t分別為10.55、5.74,P<0.01),穿膜細胞數量明顯減少(t分別為11.46、8.59,P<0.01)。見圖6和圖7。

*:P<0.01,與空載體對照組比較

*:P<0.01,與空載體對照組比較
2.6 敲減cGAS對TE-1細胞凋亡的影響
流式細胞術檢測結果顯示,sh1-cGAS、sh2-cGAS組TE-1細胞早期凋亡和晚期凋亡細胞所占百分比較對照組明顯增加(t分別為16.16、11.68,P<0.01),見圖8。

*:P<0.01,與空載體對照組比較
3 討論
基因組不穩定性是導致腫瘤發生發展的關鍵因素,通過同源重組準確修復DNA雙鏈斷裂可以保留基因組完整性并抑制腫瘤發生,cGAS是一種胞質DNA傳感器,可通過啟動STING-IRF3-Ⅰ型IFN信號傳導級聯來激活先天免疫[1,9]。以往的研究大多集中于探索cGAS在固有免疫細胞中的功能與調節機制,近年來的研究也發現其在腫瘤細胞中發揮了一定的調節作用。cGAS在細胞質內可結合因損傷而積累的雙鏈DNA,進而激活下游的cGAS-STING信號通路,并最終促進包括促炎因子在內的多種基因表達,將MHC-Ⅰ分子結合肽呈遞在腫瘤細胞表面,使腫瘤細胞可被CD8+T細胞識別[10],進而促進了抗腫瘤免疫的功效。也有研究表明腫瘤放射線處理后可以通過cGAS-STING途徑誘導細胞表面PD-L1表達的上調,進而改變腫瘤微環境并抑制腫瘤殺傷性T細胞的功能[11]。本研究結果顯示,cGAS在食管鱗癌組織中的表達較癌旁組織明顯升高,且進一步在永生化的食管鱗癌細胞系和食管正常上皮細胞中對應地證明了這一點,提示其在食管鱗癌中可能發揮了調節功能。
最近的研究表明,在腫瘤細胞發生DNA損傷的情況下,cGAS會進入到細胞核中,并被招募至DNA雙鏈斷裂部位,并直接與組蛋白變體γH2AX發生直接相互作用,進而參與斷裂DNA的同源重組與損傷修復。對應的,過度表達的cGAS增加了體外腫瘤細胞的惡性潛能,而肺癌細胞中敲除cGAS后減少了小鼠中形成的腫瘤體積[4,12]。本研究中cGAS敲減后顯著抑制了食管鱗癌細胞的增殖與遷移能力,且敲減的細胞凋亡明顯增加。我們推測cGAS可能作為一種新的癌基因促進腫瘤的發生發展,因此cGAS可作為癌癥治療新的潛在分子靶點[13-14]。但是cGAS在食管鱗癌細胞中是否也參與了DNA的損傷修復還有待進一步的實驗來確定。后續我們將進一步擴大臨床樣本,深入地研究cGAS在食管鱗癌中的調節功能與相關的分子機制,并評估cGAS與臨床食管鱗癌治療效果和預后的關系。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
