海參體壁自溶機制及其控制技術的研究進展
2020-03-03 10:05:42
食品工業科技 2020年3期
(青島科技大學化工學院,山東青島 266042)
海參是一種海洋棘皮動物,其體內含有多種生物活性物質,如巖藻糖基硫酸軟骨素、巖藻聚糖、皂苷、腦苷脂、糖基鞘磷脂和膠原蛋白等[1-2],具有抗癌[3]、抗凝血[4]、抗氧化[5]的作用。因此海參具有較高的商業價值,是中國、日本、韓國、越南、印度等許多亞洲國家重要的養殖水產物種之一,也是澳洲以及美國、加拿大、墨西哥等歐美國家亟待開發的水產資源。鮮活海參如仿刺參(Stichopusjaponicus)的體態一般呈平圓筒狀,表皮整體完好,有彈性;背部略隆起,具有圓錐狀肉刺,海參的主要食用部分為海參體壁組織,然而,海參在收貨、運輸、貯藏和加工過程中極易發生自溶。自溶時海參會發生“吐腸”現象,這是一種自我保護的條件反射;海參體表松弛,白色真皮外露,出現脫皮現象;海參體壁的可變結締組織中的膠原蛋白纖維、微纖維間的間隙加大,膠原纖維分解為膠原原纖維和膠原纖維束,纖維間的蛋白聚糖橋消失,纖維網絡解體,繼而膠原纖維的形態和輪廓變模糊,部分原本堅固的體壁形成粘稠團,局部發生“溶解”,甚至最終“融化”為液態,其成分主要為蛋白、多糖等[6-7]。海參自溶速度極其迅速,快者可在幾分鐘內致使海參體壁發生局部液化,從而導致海參品質嚴重下降,進而造成極大的經濟損失。為此,本文針對海參自溶,綜述了海參體壁的組成、海參自溶機制和自溶控制技術,為進一步研究海參自溶機制和控制技術提供理論依據。
1 海參體壁的組成及其自溶過程中的變化
海參體壁由角質層、皮層、肌肉層組成,其中皮層中的真皮占比最大,為可變結締組織。該組織是由膠原纖維和微纖維網絡構成的纖維網絡結構。如圖1所示,膠原纖維呈草繩狀平行緊密排列,通過蛋白聚糖橋連接[6-8],被微纖維網絡包圍,微纖維網絡維持組織并提供遠程恢復力[9]。膠原纖維的主要結構單元是膠原原纖維和膠原纖維束。膠原纖維及其網絡結構的主要組成是海參多糖和膠原蛋白。
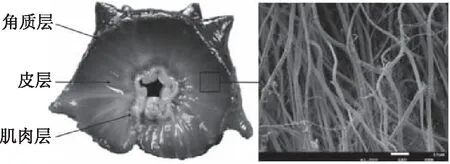
圖1 海參體壁剖面圖及其體壁膠原纖維Fig.1 Contraverse section and collagen fibers of body wall of sea cucumber
海參多糖主要由海參硫酸軟骨素和巖藻聚糖構成,其相對分子質量分別為4×104~5×104、8×104~10×104[10]。Chen[11]等得到了巖藻硫酸軟骨素的結構,如圖2(a)所示;Yu[12]等在刺參體壁中得到了海參巖藻多糖,其結構如圖2(b)所示。目前發現的海參多糖的單糖組成為N-乙酰半乳糖胺、半乳糖胺、半乳糖、巖藻糖、葡萄糖胺、葡萄糖醛酸、甘露糖等[13-14],糖苷鍵的連接方式為α-1,3、α-1,4[12-15]。硫酸基團在糖單元中的結合位點(2、3、4、6位)和數量(19.54%~29.95%)因海參種類而異[16]。海參多糖在自溶過程中可以釋放68%的硫酸多糖,被分解并轉化成脫氧半乳聚糖[17],其結構如圖2(c)所示。
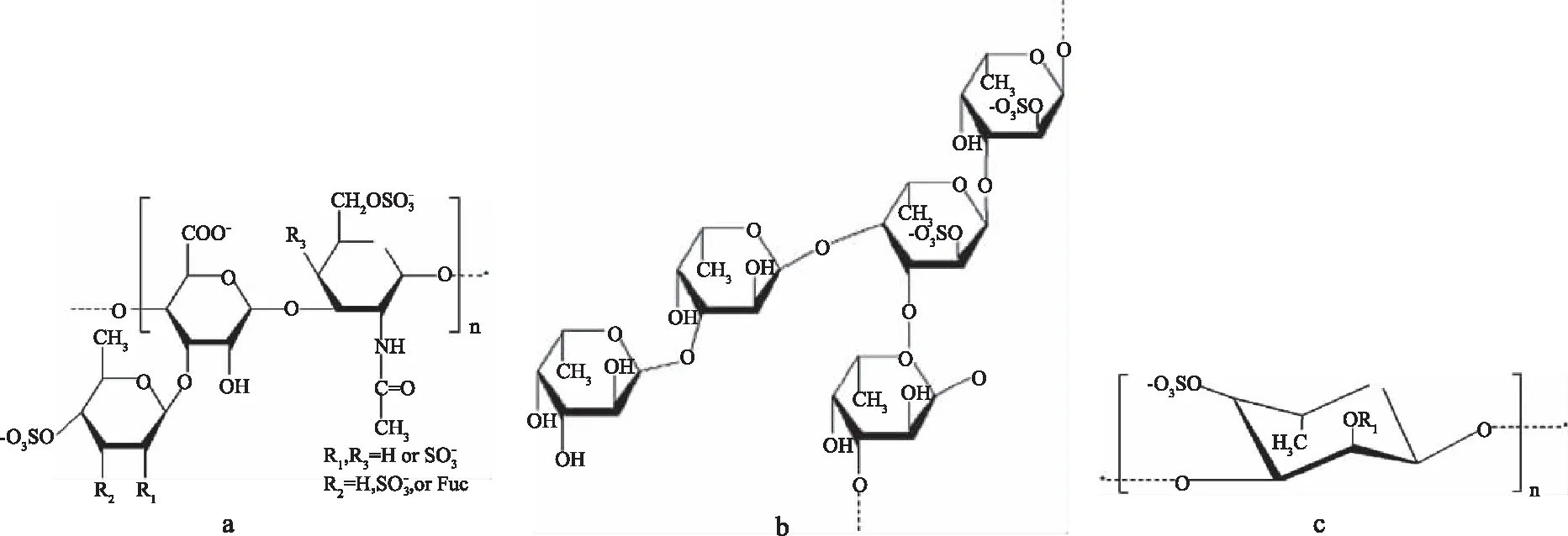
圖2 海參體壁中的巖藻硫酸軟骨素(a)和海參巖藻多糖(b)以及自溶產物脫氧半乳聚糖(c)Fig.2 Chondroitin sulfate(a)and polysaccharide(b)in the body wall of sea cucumber and the resultant deoxygalactosan(c)after autolysis
海參膠原蛋白是結締組織中的主要組成成分。每個膠原蛋白分子由三條肽鏈(三條肽鏈可以彼此相同,也可以彼此不同)相互纏繞成草繩狀而成,呈右手超螺旋結構。膠原蛋白的相對分子質量約為300 kDa,長300 nm,直徑1.5 nm,其中每條肽鏈約有1000個氨基酸殘基,相對分子質量為95~100 kDa[18]。在自溶過程中,海參體壁中TCA可溶性寡肽的溶出率隨之逐漸增大[19],說明體壁中的蛋白結構可能發生酶促降解。
2 海參自溶的機制
海參體壁自溶過程極其迅速,基本上可歸因于海參體壁內源酶。當然,外源微生物的存在也受到關注。
2.1 海參內源酶
2.1.1 海參體壁酶 海參體壁自溶與即食海參的體壁失穩與海參體壁中海參多糖和膠原蛋白降解有關,而這些降解與海參體壁中的多糖酶和蛋白酶有關[20-21]。
目前已發現的多糖水解酶有α-糖苷鍵水解酶,如以α-淀粉和果膠等為底物可探知其具有淀粉酶和果膠酶等活性[22-23]。張杰[22]等通過從海參體壁中分離純化并得到了α-1,4淀粉酶,其分子質量約為420 kDa。劉征東[23]等從海參體壁中分離純化了果膠酶,并發現該酶是一種耐強熱酶,在90 ℃條件下保溫20 min后活力無顯著性變化,可水解高溫處理的海參體壁組織,產生更多的還原端。海參經過煮沸或者高壓蒸汽滅菌后,其體壁中的果膠酶仍有較高的活力[24],即具有較高的熱穩定性。
海參體壁蛋白酶主要有組織蛋白酶L、半胱天冬酶、基質金屬蛋白酶和組織蛋白酶K。Zhou[25]等從海參體壁中分離得到的組織蛋白酶L可催化水解酰胺鍵(以Z-Phe-Arg-MCA為底物)。該酶存在于細胞液泡內表面,在紫外照射下液泡膜與細胞膜融合,使該酶從細胞中釋放出來,并擴散到組織中,與細胞外基質如膠原纖維接觸并降解[6]。Qi[26]等發現,細胞凋亡可誘導增強該酶活性,加劇海參的自溶。可能的原因是,絲裂原活化蛋白激酶與應激誘導凋亡信號通路密切相關,調控細胞因子產生、炎癥、生長、分化、存活、凋亡等關鍵下游基因的轉錄。紫外照射會增強海參中自由基的形成,激活絲裂原活化蛋白激酶家族中的應激活化蛋白激酶通路和p-38絲裂原活化蛋白激酶通路,從而導致降解膠原蛋白,并激活半胱天冬酶-3,而后者激活組織蛋白酶L進一步降解膠原蛋白。此外,該通路也可直接導致DNA的損傷。因此,最終導致海參自溶。基質金屬蛋白酶是一類廣泛存在于動物體內的含鈣鋅內肽酶,主要包括膠原酶和明膠酶。膠原酶降解膠原蛋白,明膠酶降解明膠和一些基底膜蛋白。基質金屬蛋白酶在海參可變結締組織的合成和水解中發揮著重要作用,Sun[27]等發現基質金屬蛋白酶是海參自溶過程中的關鍵酶,參與了海參自溶過程中蛋白質的降解,其作用集中在膠原纖維的高級結構[28]。海參體壁中組織蛋白酶K也參與了海參自溶,組織蛋白酶在酸性條件下可降解膠原蛋白,海參組織的堿性環境可隨著自溶的進行,溶酶體的溶解和組織蛋白酶的釋放,變為酸性環境,從而使組織蛋白酶K發揮活性。組織蛋白酶K能夠對具有穩定三螺旋結構的膠原進行多位點降解,可降解Ⅰ和Ⅱ型膠原蛋白以及非膠原蛋白[29]。總之,海參體壁自溶與其體內的蛋白酶關系密切。
2.1.2 海參腸道酶 半胱天冬酶可在紫外照射下被激活,參與自溶。Su[30]等發現紫外刺激海參可使線粒體膜功能障礙,細胞色素c從線粒體釋放到胞漿中激活半胱天冬酶并參與海參自溶。半胱天冬酶抑制劑可以不可逆地結合到蛋白酶的催化位點,抑制蛋白酶的活化,從而降低海參自溶。海參腸壁多糖水解酶可以作用于α-1,4糖苷鍵和β-1,3糖苷鍵,其中淀粉酶活力最高,是一種金屬蛋白酶,在60 ℃內活力穩定[31]。然而,由于海參體壁自溶極其迅速,其與海參內臟酶的相關性尚需進一步研究。
2.2 外源微生物
除了海參自身能夠產生多種多糖酶和蛋白酶外,海參腸道中的多種微生物也可以產生多糖酶和蛋白酶。海參腸道微生物已被分離和鑒定,如芽孢桿菌和枝芽孢桿菌[32]、放線菌[33]和立克次氏體[34],可產生多種外源性消化酶,如淀粉酶、蛋白酶、纖維素酶,對海參的食物消化有貢獻。采用分類器對海參消化道中微生物進行分類,得到10個細菌門,23綱44目83科127屬[32,35]。但是,沒有相關文獻報道這些外源微生物是否參與海參自溶。Bakunina[36]等從海洋檸檬假單胞菌(Pseudoalteromonascitrea)中分離得到三種可以產生巖藻聚糖酶的枸櫞酸菌株。Chang[37]等分離出了一種新的黃桿菌科菌株-CZ1127,該菌株可以產生多種巖藻聚糖降解酶來利用海參巖藻多糖。由于海參體壁自溶發生于體壁內部,且自溶速度極快,因此,很難將海參體壁外源微生物與海參體壁自溶聯系起來。
3 自溶的控制技術
3.1 熱力滅酶
熱力滅酶包括高溫熱力滅酶和真空熱力滅酶。在高溫熱處理(121 ℃ 15 min)過程中,海參中的微生物和內源蛋白酶會完全失活,膠原纖維束的結構被破壞和斷裂,膠原蛋白的三螺旋結構也逐漸解旋和降解。在貯藏過程中,膠原纖維束和膠原蛋白的結構被進一步破壞和降解,且被破壞的程度與貯藏溫度呈正相關,其中,硬度和咀嚼性下降極為明顯,彈性、凝聚性和恢復性均有所下降,膠原纖維最初緊密交錯排列,之后結構斷裂,排列混亂[38-39]。這種變化有可能來源于海參體壁中的多糖酶,高溫處理后,該類酶活力不降反升[23-24]。采用真空熱力滅酶法時,設置加熱溫度為95 ℃,真空鍋真空壓力為0.04 MPa,加熱時間為3 h,雖然得到的海參有較高含量的蛋白質、脂肪、多糖、礦物質、氨基酸和微量元素,但仍會造成部分營養流失[40-41]。采用熱力滅酶,雖然抑制了酶對海參的自溶,但也明顯破壞海參的組織結構并使其外部結構大幅度縮小,而影響其商業價值。
3.2 超高壓技術
為了進一步提高自溶酶的滅酶力度,超高壓(200 MPa以上)技術應運而生。該條件會使微生物細胞壁和細胞膜的結構遭到破壞,蛋白質降解,酶活性降低,幾乎會殺死所有細菌、霉菌和酵母,但不會像高溫滅菌一樣造成營養流失和風味變化[42]。
超高壓處理海參后,能夠有效鈍化海參體內自溶酶活性,其中蛋白酶和淀粉酶活力被明顯抑制。如在600 MPa、保壓時間30 min、處理溫度15 ℃條件下制作的超高壓海參軟罐頭,其室溫貯藏期為60 d,冷藏貯藏期為90 d[25];在300 MPa、保壓時間10 min、處理溫度60 ℃的條件下,處理得到的刺參持水力大,且該工藝能夠很好的改善刺參的質構,并有效地保留了刺參中的功能成分[43]。把該滅酶技術應用于海參加工工藝中,如果在海參的風味方面有進一步的改善,將有望獲得消費者樂于接受的即食海參產品。
3.3 超聲波輔助電解水
電解水是將食鹽、鹽酸等含有氯離子的溶液電解,生成含有次氯酸的溶液,其中,微酸性電解水的pH維持在5.0~6.5。在微酸性電解水中主要起殺菌作用的物質為次氯酸。次氯酸能損害細胞膜,致使細胞內的蛋白酶、RNA和DNA無法發揮正常的生化活性,導致微生物死亡,且與微生物及有機物接觸后立即還原為水。然而,電解水只能殺滅海參表面的微生物,因此,需聯合超聲波技術抑制海參自溶酶活性[44]。
3.4 高壓電場干燥
高壓電場可增加細胞膜的跨膜電壓,使其變薄并形成微孔甚至崩解,導致細胞死亡,其胞內酶的二級和三級結構也相應遭到破壞[45],從而達到抑制海參體壁自溶的目的。利用該法干燥海參,其復水率、蛋白質和酸性粘多糖含量、質地、顏色、外觀和風味雖然略遜于真空冷凍干燥法[46],但解決了海參體壁自溶問題,且使干燥成本明顯降低。
3.5 酶抑制劑
目前產業中對捕撈海參的加工前都進行滅酶處理,大部分采用酶抑制劑。金屬蛋白酶抑制劑EDTA Na2和1,10-菲羅啉可以抑制基質金屬蛋白酶的活力,在EDTA Na2或1,10-菲羅啉存在下,蛋白降解幾乎完全被抑制[27-28]。半胱氨酸蛋白酶抑制劑包括反環氧琥珀酰-l-l-亮氨酸-氨基(4-胍基)丁烷、碘乙酸、乳清蛋白濃縮物,對肌動蛋白和肌動蛋白的水解均有部分抑制作用,對組織蛋白酶K的抑制率均可達到90%以上[47-48]。使用食品級金屬蛋白酶抑制劑和半胱氨酸蛋白酶抑制劑,有利于減少海參在儲運過程中的自溶,具有成本低、操作簡單等特點。
3.6 交聯
為增強海參體壁的穩定性,可以采用交聯方法延長海參的保質期。交聯方法可以分為化學交聯、生物交聯和固化交聯。化學交聯可利用五倍子提取物和訶子提取物作為交聯劑,它們含有多元酚的結構,能與蛋白質、多糖和生物堿發生化學反應。該交聯劑與膠原蛋白發生的共價結合作用使膠原蛋白結構穩定不容易降解,能夠保持海參的微觀結構,沒有難以接受的異味,海參表面能保持刺的完整性和海參完整的形狀和外觀[49]。生物交聯可以采用轉谷氨酰胺酶交聯海參,其制作方法為:將海參去內臟,清洗,預煮;在中性水溶性殼聚糖溶液中真空浸漬,取出再轉入轉谷氨酰胺酶液中固化;調味;真空充氮包裝;高壓蒸汽滅菌等。采用該方法后,膠原蛋白穩定,海參具有較長的保藏期,0~4 ℃條件下為一年[50]。固化交聯可以采用海藻酸鈉與海參交聯,該方法可以保持海參的完整性和彈性,將海參進行原料處理、海藻酸鈉固化處理、灌湯封口、殺菌加工成即食罐頭產品,更好地解決了海參在貯藏過程中參體彈性差,體壁破裂,湯汁渾濁,保質期短的問題[51]。生物交聯法具有無毒、無異物、不影響海參品質等特點,可有效保持海參的營養成分和物理外觀。
4 展望
海參體壁組成成分復雜,相互關聯緊密,由此所構成的組織結構錯綜而有致,但極易發生自溶,其自溶機制和控制自溶的技術一直是研究的熱點。從目前研究進展來看,海參自溶主要與海參內源酶有關,目前控制海參自溶的技術,如真空熱力滅酶、超高壓技術、超聲波輔助電解水、高壓電場干燥、酶抑制劑和交聯技術,主要是基于控制海參內源酶而建立的。由于海參體壁自溶的發生極其迅速,因此,很難將海參體壁自溶與其內臟和體表及體內寄生的微生物聯系起來。有關鮮活海參自溶機制的問題仍有待于進一步研究,如自溶啟動機制、參與自溶的酶系、酶攻擊的關鍵靶點、靶點對于海參體壁構架穩定性的關系以及膠原蛋白與糖類分子及其它分子之間的鍵合方式等。另外,控制海參自溶的技術也有待不斷優化和更新,既要避免海參結構破壞和營養成分流失,又能夠保證安全有效地抑制海參自溶,得到高質量海參。總之,自溶的研究方法以及海參自溶的控制技術仍是當前解決自溶問題的關鍵。
