土壤含水量對白芨與菌根真菌共生關系的影響
2020-03-03 14:37:13刁海欣黃清俊苗利媛曾歆花黃衛昌
江蘇農業科學 2020年22期
刁海欣 黃清俊 苗利媛 曾歆花 黃衛昌
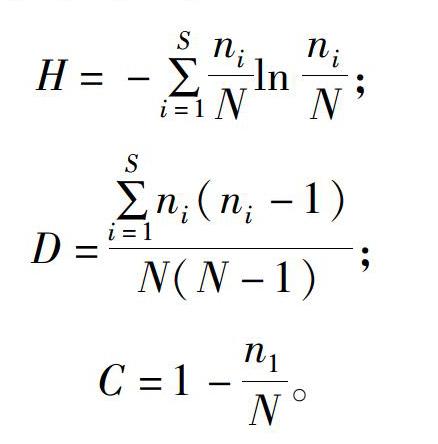
摘要:采用石蠟切片技術、DNA分子鑒定技術和穩定同位素技術相結合的手段,研究不同土壤碳氮含量對白芨菌根真菌多樣性、菌根結構特征及碳氮穩定同位素特征和光合特性的影響。結果表明,不同土壤含水量條件下,白芨菌根真菌共鑒定出305個OTU,8門99屬。菌根真菌總物種數和α多樣性指數隨著土壤含水量的減少而增加(土壤含水量為15%的真菌總數比土壤含水量為45%時多了18個OTU);同時隨著土壤含水量的降低,優勢菌根真菌逐漸轉變為傘菌綱Agaricomycetes、Serendipita和肉座菌目Hypocreales真菌;基于門水平,子囊菌門Ascomycota真菌豐度隨著土壤含水量減少而明顯減少,而擔子菌門Basidiomycota真菌豐度波動不大;菌根真菌侵入方式不受土壤含水量影響,均是通過通道細胞進入皮層組織,但菌絲團數量和侵染率隨著土壤含水量的減少而降低;此外,白芨葉片的碳氮同位素豐度隨著土壤含水量的減少而逐漸增加,凈光合速率卻隨之明顯下降,土壤含水量為45%時是白芨與菌根共生的最佳水平。研究結果明確了白芨與菌根真菌共生關系對土壤含水量變化的響應以及對土壤含水量的耐受范圍,為白芨屬植物野生種群恢復提供參考。
關鍵詞:白芨;菌根真菌多樣性;高通量測序;顯微結構;碳氮穩定同位素;土壤含水率
中圖分類號:S682.310.1文獻標志碼:A
文章編號:1002-1302(2020)22-0122-09
作者簡介:刁海欣(1994—),女,河南商丘人,碩士研究生,主要從事蘭科植物菌根生態學研究。E-mail:dhxtx1216@163.com。
通信作者:曾歆花,博士,助理研究員,主要從事蘭科植物與菌根真菌共生關系研究,E-mail:zengxinhualuxi@163.com;黃衛昌,博士,教授級高級工程師,主要從事蘭科植物保育研究,E-mail:hwc_zx@126.com。
大多數蘭科植物都與菌根真菌形成密切的共生關系[1]。近年來大部分蘭科植物的野生物種和生境生態都遭到嚴重破壞,已被列為瀕危物種[2]。研究表明,土壤水分含量是影響蘭科植物與菌根真菌共生關系的關鍵因素[3-5]。魏明等研究發現,在干旱脅迫下菌根真菌可以提高石斛的相對含水量和體內抗氧化酶活性,增強植株對干旱的耐受性[6]。Diez指出,蘭科植物的種子萌發需要適宜的共生真菌,并且受土壤濕度、有機物和酸堿度的影響[7]。McCormick等研究發現,蘭科植物與膠膜菌屬真菌在正常環境條件下形成互惠共生體,但在干旱條件下,共生真菌會由膠膜菌屬真菌轉換為其他真菌物種[8]。DNA分子鑒定技術是當前研究菌根真菌多樣性的有效手段,可以明確菌根真菌物種組成及豐度變化等情況,進而了解二者共生關系對干旱的響應機制,為蘭科植物保育工作提供參考。
白芨(Bletillastriata)是地生蘭,其藥用和觀賞價值很高[9]。當前菌根生態學逐步得到重視,但土壤因子對白芨與菌根真菌共生關系的影響研究不足。本研究對白芨進行分子鑒定、顯微切片觀察及碳氮穩定同位素和光合參數測定,比較不同土壤含水量條件下白芨菌根真菌多樣性、真菌侵染過程和生態適應性的差異,探討白芨與菌根真菌共生關系對土壤含水量變化的響應機制,明確適宜干旱逆境的優勢菌根真菌及量化最佳的土壤濕度條件。
1材料與方法
1.1材料
白芨采自上海辰山植物園竹林的2年生人工栽培種,種源地為江西崇義。于2019年4月挖取60株無病蟲害、長勢較好且規格一致的幼苗,同時修剪干枯、有蟲害的營養根。
火山泥與黃沙(體積比1.5∶1)混合基質,使用前在121℃的高壓滅菌鍋內滅菌30min,風干2d后備用。
1.2方法
1.2.1試驗設計
設置4個土壤含水量水平,分別為15%(W1)、25%(W2)、35%(W3)、45%(W4),以W4(土壤最適含水量)為控制對照組,每個水平10個重復。每盆(口徑高度為10.20.95cm)加入相同的混合基質505g,每盆種植1株幼苗。將所有盆栽放置于室外,并定期澆充足的雨水,保證幼苗健壯生長。白芨種植1個月后,水分脅迫處理3個月,每隔1d澆1次水。土壤含水量通過稱質量法(HJ613—2011《土壤干物質和水分的測定重量法》)計算獲得:首先在鋁盒中放入20g風干土樣并稱質量,然后105℃烘干6h,直至恒質量,計算出土壤含水量,進而換算出每個土壤含水量水平下每盆需要的澆水量。
1.2.2DNA分子技術
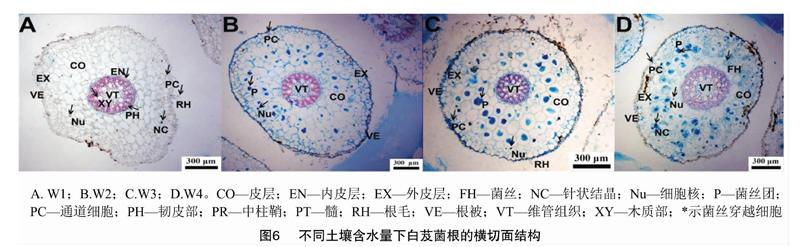
2019年7月采集白芨營養根,每個土壤含水量水平選擇6個植株,2株為1個重復,共3個重復。每個植株取3~5個長勢良好的營養根,將其放入已編號的塑封袋中,并立即裝入冷藏保溫袋中帶回實驗室處理。首先用流水沖洗根表面泥沙等雜質,然后依次按下面步驟消毒滅菌:無菌水30s,70%乙醇2min,2.5%次氯酸鈉溶液5min,70%乙醇30s,無菌水沖洗3次。一部分根放入無菌塑封袋中于-80℃保存用于后續分子生物學鑒定;另一部分放入FAA固定液中(1d 利用FastDNASpinKitforSoil(MPBiomedicals,美國)提取菌根真菌總DNA,并用1%的瓊脂糖凝膠電泳檢測DNA質量,DNA濃度和純度測定采用NanoDrop2000法。對白芨菌根真菌ITS序列進行PCR擴增(特異引物為ITS1F:5′-CTTGGTCATTTAGAGGAAGTAA-3′和ITS4R:5′-TCCTCCGCTTATTGATATGC-3′),擴增流程:95℃預變性3min;95℃變性30s,55℃退30s,72℃延伸45s,37個循環;72℃穩定延伸10min,最終10℃保存。其中PCR反應體系(20μL)為:5×TransStartFastPfu緩沖液4μL,2.5mmol/LdNTPs2μL,5μmol/L上游引物0.8μL,5μmol/L下游引物0.8μL,TransStartFastPfuDNA聚合酶0.4μL,BSA0.2μL,模板DNA10ng,補足ddH2O。每個樣本3個重復。在Illumina公司的MiseqPE300平臺上測序(上海美吉生物醫藥科技有限公司),并將原始數據上傳至NCBISRA數據庫(SequenceReadArchive,http://www.ncbi.nlm.nih.gov/Traces/sra)(序列號:SRP256191)。 使用上海美吉生物醫藥科技有限公司云平臺進行交互式生物信息云分析(http://www.i-sanger.com)。其中原始序列使用Trimmomatic軟件進行質控,使用FLASH軟件拼接序列,使用UPARSE軟件(version7.1http://drive5.com/uparse/)OTU聚類分析序列(相似度:97%),利用RDPclassifier(http://rdp.cme.msu.edu/)以及比對Unite(Release6.0http://unite.ut.ee/index.php)真菌數據庫(比對閾值:70%)注釋序列;使用mothur(versionv.1.30.1http://www.mothur.org/wiki/Schloss_SOP#Alpha_diversity)分析菌根真菌多樣性指數,包括豐度(S)、香農指數(H)、辛普森指數(D)和覆蓋率(C),同時可操作分類單元(OTU)相似水平為97%。計算公式如下: 1為僅有1條序列的OTU數量,個。 1.2.3石蠟切片技術 用無菌刀片把營養根切成2~3mm的根段。常規石蠟切片制作,即脫水、透明、浸蠟(石蠟熔點:56~60℃)、包埋、切片(厚度:8μm)、粘片、脫蠟、染色(番紅固綠對染)、封片(中性樹膠)。隨機選取不同土壤含水量水平的切片于光學顯微鏡下觀察。菌絲條條清晰、明顯疏松的菌絲團是絲狀菌絲團,而菌絲結構難辨的致密菌絲團是團塊狀菌絲團[3]。 參照Biermann等提出的根段侵染率加權法[10]計算真菌侵染率:每個水平隨機選擇300個視野,并將根段侵染程度劃分為4個等級,即C0、C1、C2和C3,其中C0表示皮層細胞中無菌絲團,C1、C2和C3分別表示1%~30%、31%~60%和>60%的皮層細胞中有菌絲團。計算公式如下: 1.2.4穩定同位素技術 于2019年7月底采集白芨新鮮葉片,每個土壤含水量水平選擇6株健壯植株,2株為1個重復,共3個重復。每株取3張長勢良好的葉片。同時采集自養非蘭科植物作為對照的葉片,即花葉滇苦菜、龍葵、小飛蓬、蓮子草,每個物種3次重復,每次6張葉片。流水沖洗干凈葉片,置于烘箱120℃殺青20min,65℃烘干48h至恒質量,最后粉碎烘干的葉片。用錫箔杯包裹2~4mg葉樣后,在元素分析儀內燃燒并被還原為純凈的CO2和N2氣體,CO2稀釋后進入穩定同位素質譜儀檢測。用相對豐度(δ值)表示穩定同位素的自然豐度變化,公式如下: 1.2.5葉綠素含量測量和光合參數測定 1.2.5.1葉綠素含量測定 于2019年7月底采用丙酮-乙醇(體積比1∶1)混合液法測量白芨葉片的葉綠素含量[11]。每個水平選擇6株健壯植株,2株1個重復,共3個重復。用孔徑R=6mm的打孔器在相同位置(避開主脈)的新鮮葉片上打孔4片,約1.13cm2;切成細絲放入5mL丙酮-乙醇混合液中,并用錫紙包裹試管,浸提24h,期間搖晃3~4次,使綠色盡可能褪完全;吸取1.5mL浸提液于比色皿中,用紫外可見分光光度計分別測量波長645nm和663nm處的吸光度。計算公式如下: 1.2.5.2光合參數測定 于2019年7月底,09:00—11:00(晴朗天氣),使用Li-600XT光合儀測定白芨葉片的凈光合速率、氣孔導度、胞間CO2濃度和蒸騰速率。每個水平選擇6株長勢良好的植株,2株為1個重復,共3個重復。測量上部完全開展的第1、第2張功能葉(從上往下數第2、第3張)的中部,同時光合儀設置為:LED2×3光源,光照度為500μmol/(m2·s),CO2濃度與環境濃度相近,流速設置為500μmol/s。 1.3數據處理 利用Excel2010和SPSS25.0軟件進行數據分析,其中使用單因素方差分析(one-wayANOVA)比較白芨在不同土壤含水量條件下菌根真菌多樣性指數、菌絲團數量、碳氮穩定同位素豐度值和凈光合速率的差異,并用Duncans多重比較分析顯著性(P<0.05);白芨葉片碳氮穩定同位素豐度與光合參數的相關分析采用Pearson相關性分析;利用OriginPro9.1軟件作柱形圖。 2結果與分析 2.1土壤含水量對菌根真菌多樣性的影響 2.1.1白芨菌根真菌Alpha多樣性分析 由表1可知,基于OTU水平,土壤含水量對白芨菌根真菌多樣性產生一定的影響。白芨菌根真菌群落豐度隨著土壤含水量的減少而增加,其中土壤含水量為15%時達到頂峰,為79;土壤含水量為25%時次之,為77;土壤含水量為45%和35%的真菌豐度較低,分別為45和41。不同土壤含水量水平的菌根真菌香農指數、辛普森指數和覆蓋率無顯著差異。 2.1.2白芨菌根真菌的Venn圖分析 分別在OTU水平和屬水平對白芨不同土壤含水量水平的菌根真菌多樣性進行比較。由圖1-A可知,4個土壤含水量水平的菌根真菌總物種數為305個OTU。在不同土壤含水量條件下,菌根真菌總數存在差異。土壤含水量為15%時最高(143個),其次是土壤含水量25%(140個)、土壤含水量45%(125個)、土壤含水量35%(91個)。白芨特有菌根真菌數隨著土壤含水量的減少而發生一定變化,土壤含水量45%(62個)明顯高于土壤含水量15%(54個)、25%(43個)和35%(20個)。此外,各土壤含水量水平之間都有數量不一的重疊真菌,其中4個水平的共有真菌有14個OTU。 圖1-B呈現出在土壤含水量條件下菌根真菌共99屬。白芨菌根真菌總屬數在不同土壤含水量水平下存在差異,其中土壤含水量45%和15%的屬總數較高,分別是55屬和47屬;土壤含水量35%次之,為39屬;土壤含水量25%最少(36屬)。在不同土壤含水量條件下,特有菌屬數目情況與屬總數有一定類似,土壤含水量45%(24屬)>15%(15屬)>25%(9屬)>35%(7屬)。除此之外,不同土壤含水量之間存在不同數量的重疊菌屬。 2.1.3白芨菌根真菌群落組成 在不同土壤含水量條件下,白芨菌根真菌物種組成存在顯著差異。從圖2可以看出,子囊菌門豐度隨著土壤含水量的減少而降低,土壤含水量為45%時最高(74.91%),土壤含水量為15%時僅為16.83%;而擔子菌門豐度在不同土壤含水量水平下波動不大。此外,鑒定出壺菌門Chytridiomycota分布于土壤含水量15%(2.76%)和25%(10.10%)中,少量的被孢霉門Mortierellomycota真菌出現在土壤含水量15%和35%中。 在屬水平上,不同土壤含水量水平下的菌根真菌群落組成也存在一定差異(圖3)。本研究將某一真菌占菌群總物種數的百分比≥10%判斷為優勢真菌,1%~10%的為常見真菌,≤1%的為稀有真菌[12]。隨著土壤含水量的減少,白芨的優勢菌根菌群種類發生改變。其中土壤含水量為45%(W4)中的雙足囊菌屬Dipodascus(OTU83、OTU832、OTU107等5個OTUs)、籃狀菌屬Talaromyces(OTU877、OTU103、OTU80等5個OTUs)和Cutaneotrichosporon(OTU73、OTU71)占優勢地位;土壤含水量為35%(W3)時,新赤殼屬Neocosmospora(OTU878、OTU874、OTU697)、籃狀菌屬(OTU877、OTU103、OTU554)和鐮孢屬Fusarium(OTU79、OTU727)占主導地位;土壤含水量為25%(W2)時菌根真菌中以傘菌綱(OTU360、OTU198、OTU783等9個OTUs)和肉座菌目(OTU801)為優勢菌屬;土壤含水量為15%(W1)中競爭力較高的菌根真菌是傘菌綱(OTU398、OTU287和OTU246)和Serendipita(OTU289、OTU356、OTU203等6個OTUs)真菌。受土壤含水量影響,共有真菌的豐度在含水量較少的處理中明顯下降,如青霉屬Penicillium(從W4中的5.71%下降到W1中的0.52%,轉變為稀有真菌)。常見真菌數量隨著土壤含水量的減少顯著下降,種類也會存在差異。如土壤含水量為45%的常見菌根真菌有單胞菌屬Kazachstania(7.29%)、新赤殼屬(6.99%)、青霉屬(5.71%)等13屬,而土壤含水量為15%時只鑒定出Cutaneotrichosporon(5.46%)、短梗霉屬Aureobasidium(4.95%)、絲核菌屬Rhizoctonia(2.63%)等10屬。 2.1.4白芨菌根真菌Lefse多級物種差異判別分析 由圖4-A可見,土壤含水量為15%和45%時存在有顯著差異的真菌。其中土壤含水量為15%時存在1組有顯著豐度差異的真菌類群,為散囊菌綱Eurotiomycetes(從綱到目);土壤含水量45%有2組顯著豐度差異的真菌,即子囊菌門(從綱到屬)和被孢霉屬Mortierella(從門到屬)。此外,土壤含水量15%和45%中占主要優勢的真菌分別是散囊菌綱Eurotiomycetes[JP3]和被孢霉科Mortierellaceae(圖4-B)。 2.1.5白芨菌根真菌Beta多樣性分析通過Beta多樣性分析不同土壤含水量水平之間菌根真菌多樣性的差異。由圖5可知,白芨菌根真菌在不同土壤含水量條件下存在差異,同時造成該差異的最大主成分(PC1)可解釋其中23.87%的差異。 2.2土壤含水量對菌根真菌侵染過程的影響 2.2.1白芨根部結構 在不同土壤含水量條件下,白芨根部結構一致,從外至內可分為表皮(根被)、皮層、中柱3部分(圖6)。但根徑、根被厚度和中皮層細胞層數等隨著土壤含水量減少無顯著差異。在光學顯微鏡下可觀察到根最外圍的是若干層扁平磚狀表皮細胞(即根被),且土壤含水量為15%時,根被最厚(56.5μm)。根被向內是皮層組織,在根中占最大的比例,又分為外皮層、中皮層和內皮層。外皮層是緊挨著根被的1層長短表皮細胞間隔排列的多角形細胞,其中短的表皮細胞體積較小、近方形且細胞核膨大,又稱通道細胞。外皮層向內為中皮層,細胞體積較大,近似等徑球形,排列疏松,土壤含水量為15%時細胞層數最多,為(7±1)層。部分皮層細胞中出現針狀結晶。中皮層向內是由單層細胞組成的內皮層,細胞體積較小,排列緊密整齊。內皮層向內是由中柱鞘、維管束和髓組成的中柱,經番紅固綠對染后,木質部被染成紅色,韌皮部被染成藍色;當土壤含水量為45%時維管束數量最多(11個)。 2.2.2白芨在不同土壤含水量下的菌根真菌侵染方式和菌絲形態特征 由圖6可知,在不同土壤含水量條件下,白芨菌根真菌侵染方式無差異,均通過通道細胞進入根的皮層細胞,菌絲穿過細胞壁后繼續向鄰近細胞擴展并集中分布于皮層細胞的某一區域;染菌細胞的細胞核常膨大,同時可觀察到形狀不一、呈藍綠色的菌絲或菌絲團,且菌絲常向細胞核靠近。 由表2可知,白芨的真菌侵染率和菌絲團數量等菌絲特征受土壤含水量的影響存在一定差異。當土壤含水量為45%時的真菌侵染率最高,為25%,共有63個菌絲團;皮層細胞中存在2種不同的菌絲形態,即絲狀菌絲團和團塊狀菌絲團,且絲狀菌絲團居多;鄰近染菌細胞的未侵染細胞的細胞核膨大,番紅固綠對染后呈紅色。當土壤含水量為25%和35%時,真菌侵染率均下降為15%,并以團塊狀菌絲團為主,但菌絲團數量存在差異,土壤含水量25%(37個)高于土壤含水量35%(32個)。當土壤含水量為15%時菌根真菌侵染率最低(5%),僅存在少量菌絲殘結。 2.3土壤含水量對白芨碳氮同位素特征和光合特性的影響 2.3.1土壤含水量對白芨碳氮同位素特征的影響 在不同土壤含水量水平下,白芨葉片的δ13C值均明顯高于對照植物,不同土壤含水量水平之間白芨[CM(22]δ13C值變化很小,δ13C值介于-26.99‰~-28.19‰之間,變幅為1.2‰。由圖7可知,不同土壤含水量水平之間Δδ13C值無差異。白芨葉片的δ15N在不同土壤含水量水平下低于對照植物,δ15N值范圍為1.29‰~2.32‰。同時Δδ15N隨著土壤含水量的減少而增加,其中當土壤含水量為15%時Δδ15N值最高,為-1.14‰;土壤含水量為25%和35%次之;土壤含水量為45%時Δδ15N值最低,-2.18‰(圖7)。對白芨葉片δ13C、δ15N和土壤含水量進行相關分析,發現δ13C與土壤含水量呈極強負相關(相關系數為-0.824),δ15N與土壤含水量呈顯著負相關(相關系數為-0.974,P<0.05),同時δ13C與δ15N呈極強正相關(相關系數為0.933)(表3)。 2.3.2土壤含水量對白芨光合特性的影響 不同土壤含水量條件下,菌根真菌對白芨光合特性產生影響。白芨葉片的葉綠素總量在不同土壤含水量條件下無差異,但葉片凈光合速率、氣孔導度、蒸騰速率和胞間CO2濃度存在顯著差異(圖8)。凈光合速率、氣孔導度和蒸騰速率隨土壤含水量的減少而明顯降低。當土壤含水量為45%(W4)時,葉片凈光合速率6.65μmol/(m2·s)明顯高于其余3個土壤含水量水平;W4的氣孔導度0.10mol/(m2·s)也是明顯高于其余3個土壤含水量水平;同時蒸騰速率隨土壤含水量的減少而逐漸降低,即W4[JP3][2.36mmol/(m2·s)]>W3[1.55mmol/(m2·s)]>[JP4]W2[0.63mmol/(m2·s)]>W1[0.17mmol/(m2·s)]。隨土壤含水量的減少,胞間CO2濃度在土壤含水量為15%(W1)時達到頂峰(282.02μmol/mol),而土壤含水量25%(W2)最低(157.93μmol/mol)。此外,δ13C與凈光合速率呈極強負相關,與氣孔導度呈強負相關;δ15N與凈光合速率呈極強負相關(P<0.05),與氣孔導度呈極強負相關(表3)。 3討論與結論 本研究從細胞水平與分子水平、形態及生理等方面研究土壤含水量對白芨與菌根真菌共生關系的影響。結果表明,土壤含水量為45%時是白芨與菌根共生的最佳水平;干旱脅迫條件下,菌根真菌會通過增加物種多樣性和改變菌根真菌群落組成來增加對蘭科植物的養分供給,從而彌補干旱對植物所造成的生理傷害。 對于地生蘭植物來講,影響植株生長的土壤因子和影響蘭科植物分布的因子是不可分割的[8]。土壤因子對蘭科植物的重要性以及它是如何改變蘭科植物與菌根真菌共生關系是生態學的一個重要領域,而這方面的知識在很大程度上仍知之甚少。干旱常被證明是造成地生蘭植物種群分布的重要因素[13],同時土壤濕度和透氣性會直接影響菌根真菌的生長發育和菌根共生體的形成[1]。我們觀察到白芨菌根真菌總數和優勢菌屬數隨土壤含水量的減少而增加,最有可能的解釋是土壤含水量缺乏時,菌根真菌的多樣性增加是為了提高白芨的抗旱性,緩解干旱脅迫[14]。本研究發現隨著土壤含水量的減少,白芨優勢菌屬最終被傘菌綱(OTU398、OTU287、OTU246)和蠟殼耳目(OTU289、OTU356、OTU764)真菌所替代,可見這些真菌更適宜在干旱環境中生存并促進白芨菌根共生體的形成,對白芨應對干旱脅迫有重要的意義。這也進一步驗證了McCormick等的研究結果:干旱條件會抑制斑葉蘭接觸菌根真菌,迫使植物轉換共生真菌[8]。水分脅迫會抑制光合作用,降低蒸騰速率,阻礙植物生長[15]。本研究結果與以往研究結果一致,即白芨葉片的凈光合速率、氣孔導度和蒸騰速率均隨著土壤含水量的減少而下降。結合菌根真菌的種類隨著土壤含水量減少有所增加的結果,可以推測干旱對白芨所造成的生理傷害更嚴重,而菌根真菌只能緩解干旱脅迫程度,并無法完全彌補這一傷害。鑒于土壤含水量、菌根真菌和白芨三者之間的相互關系,未來在白芨等其他蘭科植物的栽培管理上一定要注意保證土壤水分充足,同時篩選出抗旱菌根真菌,針對性制備菌劑,為蘭科植物保育提供參考依據。 白芨菌根真菌的侵入方式不受土壤含水量的影響,均是通過通道細胞進入皮層細胞,且菌絲穿過細胞壁侵染鄰近細胞繼續擴展,最終在皮層細胞中定殖。這與郭順星的研究結果[16]類似,指出白芨菌根真菌是通過根毛和通道細胞侵入菌根內。白芨菌根真菌的侵染率和菌絲團數量隨土壤含水量的減少而下降,但白芨菌根真菌總數隨土壤含水量的減少而增加,可見菌根真菌侵染率和菌絲團數量只能作為不同土壤含水量條件下菌根差異比較的指標,但不可作為判斷真菌多樣性的標準。有研究表明菌絲團形態代表某一菌根真菌種類的一種表現形態[17]。本研究發現當土壤含水量為45%時存在2種形態的菌絲團(絲狀和團塊狀),而土壤含水量減少后僅存在團塊狀菌絲團。由此推測,這是白芨在不同土壤含水量條件下需要依賴不同的菌根真菌造成的。 總之,白芨與菌根真菌共生關系對土壤含水量缺乏的響應是由土壤水分缺乏程度、白芨、菌根真菌等多種因素影響的復雜反應過程,有待于進一步深入研究二者共生關系響應干旱脅迫的分子機制。 參考文獻: [1]劉潤進,陳應龍.菌根學[M].北京:科學出版社,2007:35-36,221-222. [2]梁宇,郭良棟,馬克平.菌根真菌在生態系統中的作用[J].植物生態學報,2002,26(6):739-745. [3]McCormickMK,WhighamDF,ONeillJP,etal.AbundanceanddistributionofCorallorhizaodontorhizareflectvariationsinclimateandectomycorrhizae[J].EcologicalMonographs,2009,79(4):619-635. [4]GaleSW,YamazaiJ,HutchingsMJ,etal.Constraintsonestablishmentinanendangeredterrestrialorchid:acomparativestudyofinvitroandinsituseedgerminabilityandseedingdevelopmentinNervilianipponica[J].BotanicalJournaloftheLinnaeanSociety,2010,163(2):166-180. [5]PhillipsRD,BrownAP,DixonKW,etal.Orchidbiogeographyandfactorsassociatedwithrarityinabiodiversityhotspot,theSouthwestAustralianFloristicRegion[J].JournalofBiogeography,2011,38(3):487-501. [6]魏明,童秦怡,柴瑞娟,等.蘭科菌根真菌對干旱脅迫下鐵皮石斛生長和抗氧化能力及相關基因表達的影響[J].西北植物學報,2018,38(10):1905-1912. [7]DiezJM.Hierarchicalpatternsofsymbioticorchidgerminationlinkedtoadultproximityandenvironmentalgradients[J].JournalofEcology,2007,95(1):159-170. [8]McCormickMK,WhighamDF,SloanD,etal.Orchid-fungusfidelity:amarriagemeanttolast?[J].Ecology,2006,87(4):903-911. [9]QianCD,JiangFS,YuHS,etal.AntibacterialbiphenanthrenesfromthefibrousrootsofBletillastriata[J].JournalofNaturalProducts,2015,78(4):939-943. [10]BiermannB,LindermanRG.Quantifyingvesicular-arbuscularmycorrhizas:aproposedmethodtowardsstandardization[J].NewPhytologist,1981,87(1):63-67. [11]舒展,張曉素,陳娟,等.葉綠素含量測定的簡化[J].植物生理學通訊,2010,46(4):399-402. [12]龐雄飛,尤民生.昆蟲群落生態學[M].北京:農業出版社,1996:77-103. [13]ReddochJM.PopulationdynamicsandfloweringsynchronyofGoodyerapubescens(Orchidaceae)insouthwesternQuebec,Canada[J].JournaloftheTorreyBotanicalSociety,2007,134(3):379-388. [14]McCormickMK,WhighamDF,ONeillJP.Mycorrhizaldiversityinphotosyntheticterrestrialorchids[J].NewPhytologist,2004,163(2):425-438. [15]向丹,徐天樂,李歡,等.叢枝菌根真菌的生態分布及其影響因子研究進展[J].生態學報,2017,37(11):3597-3606. [16]郭順星.藥用植物內生真菌生物學[M].北京:科學出版社,2016:632. [17]KristiansenKA,TaylorDL,KllerR,etal.IdentificationofmycorrhizalfungifromsinglepelotonsofDactylorhizamajalis(Orchidaceae)usingsingle-strandconformationpolymorphismandmitochondrialribosomallargesubunitDNAsequences[J].MolecularEcology,2001,10(8):2089-2093.
