2種海水臂尾輪蟲品系生活史特征
2020-03-03 14:37:13張凱雷馬慧娟李茹夢楊家新
江蘇農業科學 2020年22期
張凱雷 馬慧娟 李茹夢 楊家新

摘要:應用單個體和種群培養方法,研究在一定溫度(20℃)下,以濃度為0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL的海水小球藻(Chlorellasp.)為食,比較射陽褶皺臂尾輪蟲(Brachionusplicatilis,SY)和瑪雅臂尾輪蟲(Brachionusmanjavacas,BM)的生活史特征。研究表明,SY品系和BM品系的種群增長率均保持先增長后緩慢下降的趨勢。BM品系和SY品系在初期食物濃度為4×106cells/mL時的種群增長率始終高于其他濃度處理;在后期,食物濃度為8×106cells/mL的種群增長率才最高。BM品系的種群數量與種群增長率變化趨勢一樣;而SY品系在食物濃度為8×106cells/mL時的種群數量一直高于其他濃度。2個品系輪蟲的生殖前期都隨食物濃度升高而降低。在(0.25×106、0.5×106cells/mL)時,BM品系的生殖前期比SY品系的生殖前期要長;在(4×106、8×106cells/mL)時,SY品系的生殖前期比BM品系長;在(1×106、2×106cells/mL)時,2個品系輪蟲生殖前期無顯著差異。2個品系褶皺臂尾輪蟲的子代數均隨食物濃度增大而變多,在低濃度時,SY品系子代數高于BM品系;在中高濃度時,BM品系子代數高于SY品系。2個品系輪蟲的壽命隨著食物濃度升高先增長后縮短,都在食物濃度為4×106cells/mL或8×106cells/mL時的壽命最長。
關鍵詞:褶皺臂尾輪蟲;品系;食物濃度;生殖前期;子代數;壽命
中圖分類號:S963.21+4文獻標志碼:A
文章編號:1002-1302(2020)22-0169-05
通信作者:楊家新,男,教授,博士生導師,從事輪蟲繁殖生物學研究。E-mail:1063859395@qq.com。
褶皺臂尾輪蟲(Brachionusplicatilis)作為海水人工育苗的一種輪蟲,其繁殖狀況的不同都會在生產上帶來不同的效益。當然,生殖前期越短,輪蟲在前期就會繁殖越快;壽命越長,維持種群高密度的時間就越長;子代數越高,密度繁殖越快。在不同食物濃度條件下,種群增長率、種群數量的變化以及生殖前期、壽命、子代數也會不同。已有研究表明,溫度和食物濃度的不同會對輪蟲生殖期產生影響[1-2],品系的不同也會對萼花臂尾輪蟲的生活史產生影響[3]。也有研究發現。萼花臂尾輪蟲的姐妹種和它們的地理位置并沒有顯著差異[4]。Rehberg-Haas等也研究了不同巴芙藻對褶皺臂尾輪蟲的影響[5]。Cheng等研究了鹽度和食物濃度對圓形臂尾輪蟲種群動態的影響[6]。除了藻種,還有輪蟲品系、溫度、鹽度、光照和食物濃度等都對褶皺臂尾輪蟲生長都具有影響,由于環境因子的多樣性,褶皺臂尾輪蟲的生殖發育也會發生不同改變。
本試驗以褶皺臂尾輪蟲(SY)和瑪雅臂尾輪蟲(Brachionusplicatilis,BM)這2個品系輪蟲為研究對象,探討在不同食物濃度下2個品系輪蟲的生長狀況及生態學參數(生殖前期、壽命、子代數)的差異。為尋找2個品系輪蟲在不同生長階段最適的食物濃度范圍,確認不同的食物濃度對輪蟲的影響及是否與品系有關,本研究對中國射陽品系和美國品系的輪蟲進行研究比較。美國BM品系之所以能夠成為廣泛的研究對象,是因為其具有穩定的特點,本研究想通過試驗來確認中國SY品系輪蟲是否也像BM品系一樣具有穩定的特點,能否成為中國本土特有的品系,并能夠成為一種廣泛的試驗對象。
1材料與方法
1.1輪蟲的來源和預培養
1.1.1輪蟲來源SY和BM品系都是從冷凍休眠卵中孵化而來的,SY品系休眠卵底泥是從射陽池塘中獲取,BM品系休眠卵是美國Snell教授所贈。輪蟲休眠卵采用馮大慶的人工海水進行孵化,孵化溫度為(20±1)℃,光照度為4000lx,光∶暗(L∶D)=1∶1[7]。在實驗室中對這2個品系輪蟲進行單個體克隆培養。輪蟲培養所需餌料由F2培養基培養:將處于指數增長期的海水小球藻離心濃縮,再加海水培養基配制成懸液喂養輪蟲。F2培養基(1L海水中所加的量):1mL7.5g/100mLNaNO3、[JP3]1mL0.5g/100mLNaH2PO4·H2O、1mL3g/100mLNaSiO3·9H2O、1mL微量元素溶液、1mL維生素溶液。[JP3]其中,100mL微量元素溶液含有0.315gFeCl3·6H2O、0.436gNa2EDTA·2H2O、0.98mgCuSO4·5H2O、0.63mgNa2MoO4·2H2O、2.2mgZnSO4·7H2O、1.0mgCoCl2·6H2O、18.0mgMnCl2·4H2O。50mL維生素溶液含有0.05mg維生素B12、0.05mg維生素H、0.01g維生素B1。
1.1.2預培養將SY和BM品系輪蟲分別放置于6種食物濃度(0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL)中進行預培養,所用的容器為50mL燒杯,環境溫度為(20±1)℃,光照度為4000lx,L∶D=1∶1。預培養期間,使輪蟲始終處于指數增長期,培養時間為1個星期以上。挑齡期小于4h且活力充足的幼體進行試驗。
1.2試驗方法
1.2.1種群數量的測定采用群體累積培養方法:通過預培養獲得處于指數增長期的輪蟲,按試驗濃度將輪蟲接種到6個不同濃度的六孔板中。試驗變量為小球藻的濃度,分別為0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL,依次記為處理1、處理2、處理3、處理4、處理5、處理6。每組設6個平行,接種密度為1ind/mL,每孔容積為5mL,各試驗條件與預培養條件一樣。試驗期間,每24h換1次輪蟲培養液,直至輪蟲種群密度開始下降,試驗結束。記錄每天的種群數量。
1.2.2生殖前期、子代數及壽命的測定采用單個體培養方法:通過預培養獲得處于指數增長期的輪蟲,按試驗濃度將輪蟲接種到6個不同濃度的24孔板中,小球藻濃度分別為0.25×106、0.5×106、1×106、2×106、4×106、8×106cells/mL,依次記為處理1、處理2、處理3、處理4、處理5、處理6。每組設12個平行,接種密度為1ind/mL,每孔容積為1mL,其他試驗條件均與預培養條件一樣。試驗期間,每24h換1次輪蟲培養液,記錄第1次產卵的時間,每天把新生幼體挑除,直至母體死亡,試驗結束。記錄總共所產的后代數及一共所經歷的時間。
1.3種群增長指標及測定方法
培養結束后,通過所得數據,可以計算出種群增長率(populationgrowthrate,r)。
[JZ]種群增長率(r)=(lnNt-lnN1)/t。
式中:t代表試驗時間,d;N1和Nt分別表示1d和td雌體的數目。
生殖前期指幼體到產生第1枚卵所經歷的時間。后代數指一個輪蟲一生所產的后代數。壽命表示從出生到死亡所經歷的時間,h。
1.4數據處理
應用軟件sigmaplot12.5作圖,SPSS22對試驗數據進行t檢驗。
2結果與分析
2.12個品系褶皺臂尾輪蟲種群動態變化
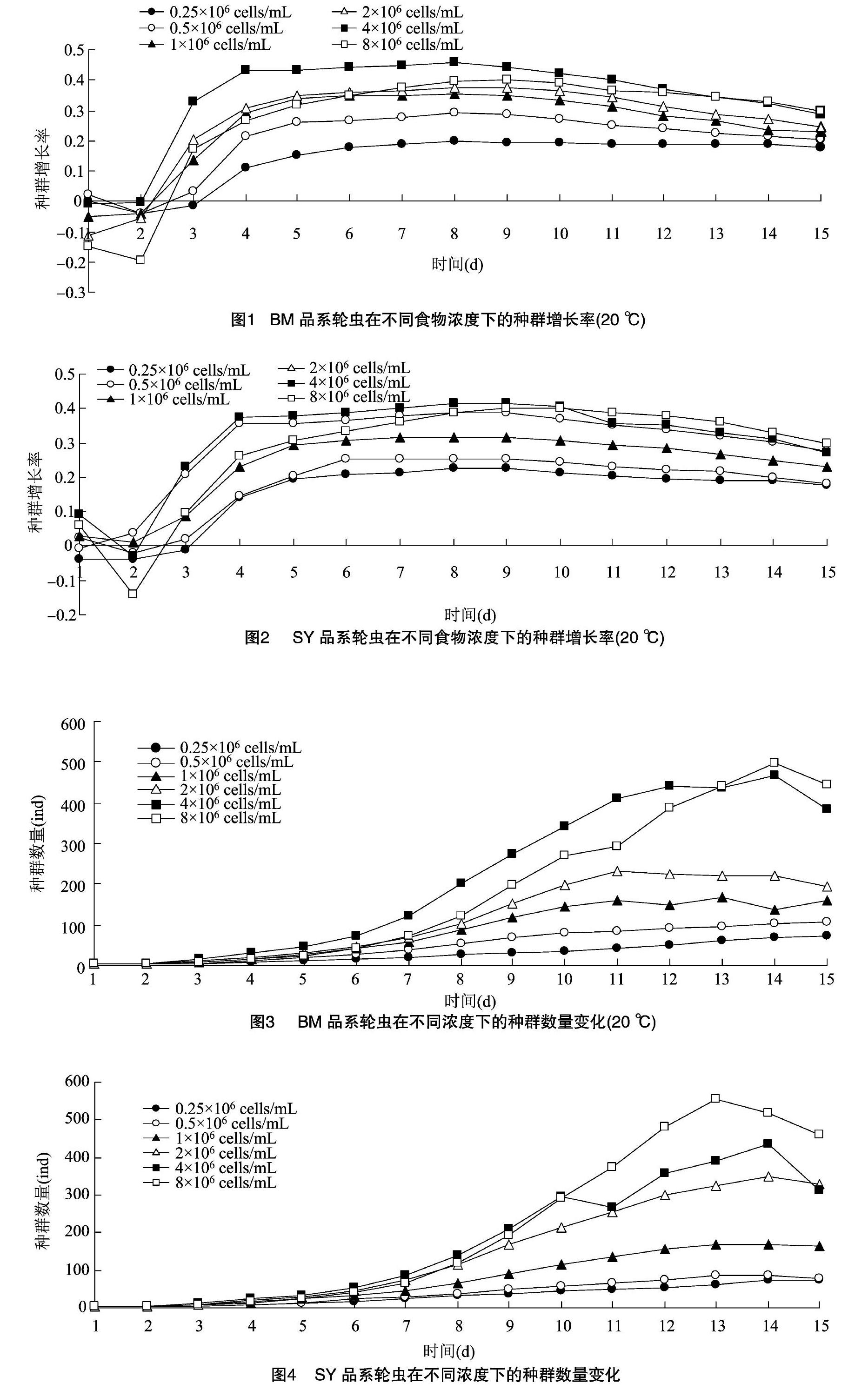
由圖1和圖2可知,2個品系褶皺臂尾輪蟲在不同濃度下,種群增長率均隨時間延長呈先快速增長后緩慢下降的趨勢。單因素方差分析表明,6d之前,BM品系處理6的種群增長率低于處理4,且顯著低于處理5的種群增長率(P<0.05,差異顯著性分析未列出,下同);13d后,處理6的種群增長率才超過處理5。8d以前,SY品系處理6的種群增長率低于處理5,10d后,處理6的種群增長率才超過處理5。除了處理6的食物濃度下,BM和SY品系的種群增長率差異不顯著外(P>0.05),其他食物濃度下SY品系的種群增長率顯著高于SM品系。
2.22個品系褶皺臂尾輪蟲種群數量變化
圖3和圖4為2個品系輪蟲在不同濃度下的種群動態變化。在5mL容量的孔板中,2個品系輪蟲的種群變化在不同濃度下均先增長后下降。在不同食物濃度下,褶皺臂尾輪蟲所到達種群高峰所需要的時間不一樣,BM品系到達峰值所需時間隨食
物濃度的升高先減少后增加,處理4所需時間最短,只需11d。各食物濃度下,SY品系到達峰值所需時間差不多。2個品系種群增長率的最高值均出現在處理5。單因素方差分析表明,6~13dBM品系處理5的種群數量要顯著高于其他濃度,到14d后處理6的種群數量才超過處理5,與處理5的種群數量差異不顯著。SY品系在3~10d時,處理5的種群數量要高于其他濃度的種群數量,與處理6不顯著;11d后,處理6要顯著高于其他濃度。
2.32個品系褶皺臂尾輪蟲生活史特征的變化
由表1可知,在20℃時,BM品系的生殖前期隨食物濃度的升高呈縮短的趨勢,在5mL容量的孔板中,處理1、處理2、處理3之間沒有顯著差異,處理5和處理6之間差異顯著。SY品系的生殖前期隨食物濃度的升高呈波浪變化,總體呈縮短趨勢,2個品系都是處理1所需的生殖前期最長,BM品系在處理6所需生殖前期最短,而SY品系生殖前期最短所需濃度為處理5。BM品系的總子代數隨食物濃度的升高呈上升的趨勢,SY品系的總子代數呈先下降后上升的趨勢。2個品系分別在處理5和處理6時的總子代數最多,BM品系在0.25×106個/mL時總子代數最少,而SY品系子代數最少時所需濃度為1×106cells/mL。BM品系輪蟲的壽命隨食物濃度的升高先延長后縮短,在處理5處壽命最長;SY品系壽命不斷延長,在處理5和處理6處壽命最長。在中低濃度時(處理1、處理2、處理3和處理4),SY品系的壽命要低于BM品系,在高濃度時(處理5、處理6)SY品系的壽命要高于BM品系。
3討論與結論
3.1BM品系和SY品系種群增長率和種群數量的變化
已有多數研究顯示,食物濃度、品系和溫度等因素都對萼花臂尾輪蟲、壺狀臂尾輪蟲有顯著影響。席貽龍等對壺狀臂尾輪蟲的研究說明了高溫會對壺狀臂尾輪蟲的發育產生負面影響,縮短輪蟲壽命[8]。本研究發現,在不同食物濃度下,2種輪蟲的種群增長率均呈先快速增長后緩慢下降的趨勢,種群變化也是先增長后下降,這與耿紅等在紅臂尾輪蟲的研究所得的結果[9]一致。胡好遠等對不同品系的角突臂尾輪蟲進行了研究,發現不同品系角突臂尾輪蟲之間的生活史特征存在著顯著差異[10]。本試驗通過研究2種國內外的輪蟲,發現2種輪蟲的種群增長率并沒有顯著差異,這與之前的研究有所不同。BM品系在處理6濃度處,13d后的種群增長率要大于處理5,這有可能是因為BM品系在13d前,處理5的濃度最適合BM品系生長;到了13d后,由于種群密度變大,處理6更能適合高密度BM品系的生長。而在SY品系中,10d前處理5的種群增長率要高于處理6,說明在初期高食物濃度對SY品系輪蟲產生了抑制作用;10d后,處理6處的種群增長率才開始慢慢超過處理5。2種品系在前期都受到高食物濃度的抑制,從2個品系種群變化可以看出,SY品系在前期高濃度處理的種群數量要高于其他濃度,BM品系在前期其他濃度處理的種群數量要低于處理5,說明了高食物濃度對BM品系的抑制要大于SY品系。姚久祥等認為,在前期低濃度會促進輪蟲的生長,食物濃度過高,會產生毒素,抑制輪蟲生長[11],本研究所得到的結論與之基本一致。
3.2BM品系和SY品系生殖前期、總子代數和壽命的比較
BM品系的生殖前期隨食物濃度的升高而縮短,食物濃度低于處理4時,生殖前期沒有顯著差異,這與席貽龍等在壺狀臂尾輪蟲所得到的食物濃度低于1.5×106cells/mL對輪蟲的生殖前期無顯著影響的結論[8]相一致;SY品系雖然也是呈縮短的趨勢,但是變化并沒有BM品系那么大,只有處理1與處理5的生殖前期存在顯著差異,其他濃度并無顯著差異。
SY的子代數隨濃度的升高呈上升的趨勢;BM的總子代數隨濃度的升高先上升后下降,在處理5處達到了最多,這說明BM的最適生長濃度有可能在處理5處,SY品系要比BM品系更能適應高濃度。
2個品系在處理5處的壽命要顯著長于低濃度,SY在處理6處與處理5處壽命相比基本保持不變,BM品系相比明顯下降,表明BM品系在處理5處的生長發育要更好,更適合BM生長。董麗麗等對3個品系的萼花臂尾輪蟲進行了研究,發現平均壽命隨食物濃度升高先增加后減小,在3×106cells/mL處,平均壽命最大[12],與本研究BM品系褶皺臂尾輪蟲所得到的結論相一致。但與SY品系的結論有所差別,有可能由于不同地理差異所產生不同的影響。BM品系和SY品系在生活史特征上有諸多異同,說明地理位置對褶皺臂尾輪蟲的生活史有一定影響,這與呂林蘭等所得到的不同品系萼花臂尾輪蟲的發育和種群增長存在顯著差異[13]相一致。
BM品系在處理6處的生殖前期和壽命要比處理5處短,子代數要少于處理5;SY品系在處理6處的生殖前期要比處理5長,BM品系種群數量最高值出現在處理5。綜合本研究可知,在20℃下,國內SY品系要比美國BM品系對高濃度海水小球藻的耐受性強,在低于處理5的食物濃度下,BM品系適合大規模培養,高于處理5濃度時,SY品系更適合大規模培養。本研究只對海水小球藻不同濃度進行了試驗,李磊等研究了不同的微藻對褶皺臂尾輪蟲的影響,認為輪蟲的攝食率與餌料類型有關[14]。有人認為,輪蟲依靠纖毛濾食,所以食物細胞的大小和密度影響輪蟲的攝食情況[15]。也有研究說明,由于藻類細胞大小不一,通過幾種藻類的換用能夠更好地促進輪蟲的生長[16]。輪蟲食物濃度低,繁殖率降低,壽命縮短,這與Yoshinaga等得到的結論[17]相一致。
參考文獻:
[1]葛雅麗,席貽龍,陳芳,等.食物濃度和品系對萼花臂尾輪蟲生殖期歷時和混交雌體形成的影響[J].生態學報,2005,25(8):3-9.
[2]Pérez-legaspiIA,Rico-martínezR.Effectoftemperatureandfoodconcentrationintwospeciesoflittoralrotifers[J].Hydrobiologia,1998,387/388:341-348.
[3]馮粒克,席貽龍.三個地理品系萼花臂尾輪蟲生活史特征的比較[J].動物學雜志,2004,39(1):12-15.
[4]XiangX,XiY,WenX,etal.PatternsandprocessesinthegeneticdifferentiationoftheBrachionuscalycifloruscomplex,apassivelydispersingfreshwaterzooplankton[J].MolecularPhylogeneticsandEvolution,2011,59(2):386-398.
[5]Rehberg-HaasS,MeyerS,LippemeierS,etal.AcomparisonamongdifferentPavlovasp.productsforcultivationofBrachionusplicatilis[J].Aquaculture,2015,435:424-430.
[6]ChengSH,KaS,KumarR,etal.Effectsofsalinity,foodlevel,andthepresenceofmicrocrustaceanzooplanktersonthepopulationdynamicsofrotiferBrachionusrotundiformis[J].Hydrobiologia,2011,666(1):289-299.
[7]馮大慶.人工海水的簡易制法[J].生物學通報,1986(10):37-39.[HJ2.14mm]
[8]席貽龍,黃祥飛.溫度對壺狀臂尾輪蟲實驗種群動態的影響[J].海洋與湖沼,2000,31(1):23-28.
[9]耿紅,席貽龍,胡好遠.食物種類和濃度對紅臂尾輪蟲種群增長、個體大小及卵大小的影響[J].應用生態學報,2003,14(5):753-756.
[10]胡好遠,席貽龍,耿紅.三個品系角突臂尾輪蟲生長和發育的比較研究[J].應用生態學報,2003,14(4):565-568.
[11]姚久祥,彭敏.藻類食物對輪蟲群體增殖的影響[J].廣西水產科技,2001(4):21-25.
[12]董麗麗,席貽龍,劉桂云,等.溫度和食物濃度對三品系萼花臂尾輪蟲實驗種群動態的影響[J].應用生態學報,2004,15(11):2165-2169.
[13]呂林蘭,董學興,董斯明,等.兩品系萼花臂尾輪蟲的生活史特征和受精率研究[J].大連海洋大學學報,2012,27(5):401-410.
[14]李磊,朱小明,吳洪喜.營養強化時褶皺臂尾輪蟲對餌料微藻的攝食[J].生態學雜志,2011(9):2019-2025.
[15]RothhauptKO.Differencesinparticlesize-dependentfeedingefficienciesofcloselyrelatedrotiferspecies[J].LimnologyandOceanography,1990,35(1):16-23.
[16]ViayehRM,MohammadiH,ShafieiAB.PopulationgrowthofsixIranianBrachionusrotiferstrainsinresponsetosalinityandfoodtype[J].InternationaleRevuederGesamtenHydrobiologieundHydrographie,2010,95(6):461-470.
[17]YoshinagaT,HagiwaraA,TsukamotoK.Lifehistoryresponseandage-specifictolerancetostarvationinBrachionusplicatilisofMuller(Rotifera)[J].JExpMarBiolEcol,2003,287(2):261-271.
