33種魚類微型反向重復轉座元件鑒定
2020-03-04 03:15:44胡競文趙連鵬韓民錦彭作剛
水生生物學報 2020年1期
關鍵詞:物種
胡競文 邵 峰 趙連鵬 韓民錦 彭作剛
(1. 西南大學生命科學學院, 淡水魚類資源與生殖發育教育部重點實驗室, 重慶 400715;2. 西南大學家蠶基因組生物學國家重點實驗室, 重慶 400715)
轉座元件(Transposable elements, TEs)是廣泛存在于基因組中的重復序列, 也稱為轉座子(Transposons), 它可以在基因組中移動并插入到新的位點[1]。轉座子由美國遺傳學家Barbara McClintock在玉米中發現[2,3], 根據轉座子不同的轉座機制, 將轉座子分為兩大類[4]。第一類是RNA類轉座子, 也被稱為逆轉錄轉座子(Retrotransposons), 這類轉座子通過“復制-粘貼”機制, 以RNA為媒介進行轉座; 第二類是DNA類轉座子(DNA transposons), 這類轉座子通過“剪切-粘貼”機制, 以DNA為媒介進行轉座。
根據能否實現自主轉座, 轉座子可以分為自主型轉座子和非自主型轉座子。非自主型轉座子最早在玉米(Zea mays)中被發現[5], 如今較受關注的非自主型轉座子是微型反向重復轉座元件(Miniature inverted-repeat transposable elements, MITEs), 它們由于缺少編碼轉座酶的序列而不能進行自主轉座。這類轉座子有一個明顯的結構特征, 序列的兩端存在反向重復序列(Terminal inverted repeat, TIR)和靶位點重復序列(Target site duplication, TSD)[5]。MITEs的序列長度通常小于800 bp, 序列中AT含量較高, 在基因組有較多的拷貝數, 可以形成穩定的二級結構, 并且能夠插入到基因內部或附近的位置[6],對基因的結構[7]、基因組的大小[8,9]和基因的表達調控[10]都有影響。目前, 植物[8,9,11]、昆蟲[12]、病毒[13,14]等基因組的MITEs預測和數據庫構建已有相關文章發表, 例如在41種植物中鑒定出3527個MITEs家族, 大多數高等植物比低等植物含有更多的MITEs,植物基因組中MITEs序列的數量與基因組大小呈正相關[10]。在98種昆蟲基因組中鑒定出6012個MITEs家族, 其中有601個新MITEs, TSD為TA的MITEs序列在昆蟲基因組中大量存在, 并且昆蟲基因組大小也與MITEs含量呈正相關[12]。
魚類是最古老的脊椎動物, 在脊椎動物進化過程中具有獨特的地位。魚類的物種數量占脊椎動物物種總量的一半以上, FishBase數據庫中記錄的魚類已達到34200種(2019年3月), 并且數量還在不斷增加。根據形態特征和棲息環境等差異, 魚類被分成多個類群[15—18], 如無頜類、軟骨魚綱、肉鰭魚綱和輻鰭魚綱。對于魚類轉座子的研究工作也在開展, 例如發現古代魚類中的DNA轉座子含量低于逆轉錄轉座子含量[19], 較大基因組魚類的轉座子多樣性更高[20], 魚類基因組中轉座子含量可能與物種棲息地等因素有關[21]等。目前, 更多研究關注轉座子對“宿主”的作用, 比如轉座子對魚類性別分化有一定影響[22], 魚類基因組加倍與轉座子擴增有關[23]。雖然魚類轉座子的研究工作不斷完善, 但是, 魚類基因組中MITEs的系統研究尚未開展。
目前, 很多魚類物種的基因組測序工作已經完成, 魚類的基因組數據、蛋白質序列可以從NCBI(National Center of Biotechnology Information)中下載, 這為魚類基因組中的MITEs鑒定提供了良好的契機, 使我們能夠運用生物信息學方法對MITEs進行鑒定, 豐富脊椎動物的轉座子數據。本研究從NCBI數據庫搜集了具有代表性的33種魚類基因組數據(包括無頜類3種、軟骨魚綱2種、肉鰭魚綱1種、輻鰭魚綱27種)并對它們進行鑒定和比較學分析以揭示MITEs在魚類基因組中的分布規律、擴增模式以及對基因組進化的影響。
1 材料與方法
1.1 魚類基因組數據的搜集
如表1所示, 從NCBI數據庫中下載33種魚類的基因組相關數據信息, 涵蓋無頜類、軟骨魚綱、肉鰭魚綱和輻鰭魚綱, 包括小丑魚(Amphiprion ocellaris)、歐洲鰻鱺(Anguilla anguilla)、日本鰻鱺(Anguilla japonica)、美洲鰻鱺(Anguilla rostrate)、墨西哥麗脂鯉(Astyanax mexicanus)、象鯊(Callorhinchus milii)、大西洋鯡(Clupea harengus)、半滑舌鰨(Cynoglossus semilaevis)、斑馬魚(Danio rerio)、蒲氏盲鰻(Eptatretus burger)、伯氏樸麗魚(Haplochromis burtoni)、貝氏隆頭魚(Labrus bergylta)、大黃魚(Larimichthys crocea)、矛尾魚(Latimeria chalumnae)、猬鰩(Leucoraja erinacea)、斑點雀鱔(Lepisosteus oculatus)、日本七鰓鰻(Lethenteron camtschaticum)、斑點叉尾鮰(Ictalurus punctatus)、斑馬宮麗魚(Maylandia zebra)、黃鱔(Monopterus albus)、布氏新亮麗鯛(Neolamprologus brichardi)、弗氏假鰓鳉(Nothobranchius furzeri)、羅非魚(Oreochromis niloticus)、日本青鳉(Oryzias latipes)、象鼻魚(Paramormyrops kingsleyae)、海七鰓鰻(Petromyzon marinus)、紅麗魚(Pundamilia nyererei)、大西洋鮭魚(Salmo salar)、亞洲龍魚(Scleropages formosus)、紅縛平鲉(Sebastes rubrivinctus)、犀角金線鲃(Sinocyclocheilus rhinocerous)、紅鰭東方鲀(Takifugu rubripes)以及花斑劍尾魚(Xiphophorus maculatus)。
1.2 基因組中MITEs的預測
使用MITE-Hunter[24](設定默認參數)對33種魚類基因組的MITEs進行預測, 將得到的MITEs作為問詢序列與基因組數據進行blastn比對(identity>80%、coverage>80%、e值<1e-5、長度<800 bp),保留在基因組中拷貝數多于3條的MITEs一致序列。然后提取MITEs在基因組的拷貝, 使用MUSCLE進行序列比對[25], Bioedit確定MITEs拷貝序列的TSD和TIR[26], 并在DAMBE中生成校正邊界后的MITEs一致序列[27]。在同一個基因組中的MITEs遵從“80-80-80”規則去冗余[4], 記錄每條MITEs的序列長度。再次使用blastn將得到的一致序列結果作為問詢序列與基因組進行比對, 利用perl腳本統計MITEs在基因組中的拷貝數和序列長度等信息。若與MITEs一致序列有相同TIR和TSD, 稱之為全長拷貝(Full-length copies); 若沒有完整TIR的拷貝序列, 稱為片段拷貝(Fragment copies)。得到MITEs在各個基因組中的含量信息后, 在SPSS中進行MITEs含量和基因組大小的Pearson相關性分析。
1.3 MITEs在魚類基因組中的擴增和分布
MITEs可以在基因組中通過轉座進行擴增, 因此我們分析了MITEs的插入時間和分布情況。通過K2P模型(Kimura 2 parameter distances)[28]計算全長拷貝序列在基因組中的插入時間, 首先使用DAMBE得到全長拷貝和MITEs一致序列之間的分化度, 進而利用公式T=K/2r[29]計算插入時間(取核苷酸位點替換速率r= 2.5×10-8[30,31],K值為分化度)。為了解MITEs在基因組中的分布, 下載所選魚類物種的基因組注釋文件, 其中的8個物種(蒲氏盲鰻、日本七鰓鰻、海七鰓鰻、亞洲龍魚、日本鰻鱺、美洲鰻鱺、歐洲鰻鱺和紅縛平鲉)沒有內含子、外顯子信息注釋文件, 不能計算插入位置, 猬鰩的注釋文件信息很少, 不能得到有效位置信息。對剩余的24種魚類, 根據MITEs全長拷貝在基因組上的位置, 使用perl腳本統計全長拷貝插入到基因內、基因外顯子和內含子中的數量。
2 結果
2.1 魚類基因組中MITEs的含量
通過MITE-Hunter對33個魚類基因組MITEs進行初步預測, 后續校準、去冗余后確定了所選物種基因組中MITEs一致序列和拷貝數, 共得到2433個MITEs一致序列, 487594個全長拷貝。不同魚類基因組的MITEs拷貝數和含量差異較大(表1)。MITEs(全長拷貝和片段拷貝)含量最高的5個物種分別是
斑馬魚(237.76 Mb, 16.66%)、海七鰓鰻(213.53 Mb,21.18%)、大西洋鮭魚(189.29 Mb, 6.38%)、墨西哥麗脂鯉(99.05 Mb, 7.42%)和日本七鰓鰻(83.17 Mb,8.07%)。也有部分物種的MITEs含量較低, 例如大西洋鯡MITEs含量為3.94 Mb(0.49%), 象鯊為3.83 Mb(0.39%), 歐洲鰻鱺為2.70 Mb(0.27%), 猬鰩的含量為1.73 Mb(0.11%), 紅鰭東方鲀基因組中MITEs含量最低, 僅為0.61 Mb(0.16%)。從分類階元來看, 無頜類的MITEs含量較高, 如海七鰓鰻; 軟骨魚綱和肉鰭魚綱魚類的MITEs含量較低, 如猬鰩和矛尾魚; 輻鰭魚綱的魚類MITEs含量最為豐富,如斑馬魚。

表1 33種魚類基因組MITEs存在情況Tab. 1 The presence of MITEs in 33 fish genomes
相關性分析發現基因組大小和MITEs含量之間呈正相關(r=0.124,P=0.045, 圖1), 比如海七鰓鰻的基因組為1007.98 Mb, 含有213.53 Mb的MITEs序列; 斑馬魚的基因組為1427.29 Mb, 含有237.76 Mb的MITEs序列。同時, MITEs序列的AT含量較高,魚類基因組中的MITEs轉座子也是如此, 它們的AT含量在51.75%(猬鰩)到61.68%(斑馬魚)之間, 均多于GC核苷酸在基因組的占比。而MITEs在魚類基因組中存在豐富的拷貝[32,33], 其中MITEs拷貝數最多的是斑馬魚(88337個全長拷貝), 最少的是紅鰭東方鲀(500個全長拷貝)。
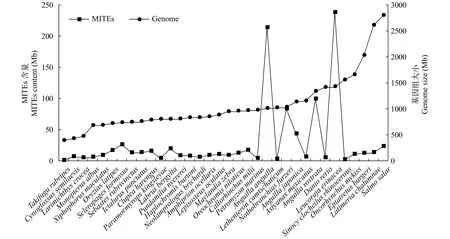
圖1 33種魚類基因組大小和MITEs含量的關系Fig. 1 The relationship between genome size and MITEs content of the 33 fish species
2.2 魚類基因組中的MITEs超家族
將得到的MITEs一致序列根據各自的TSD和TIR序列特征進行分類[12], 進一步計算各個超家族在基因組的占比。分析結果表明, 魚類基因組中MITEs超家族有: TC1-Mariner、PHIS、P、Kobolok、PiggyBac、hAT、Ginger、CMC、Merlin、Sola2, 不同魚類基因組中的MITEs超家族的分布存在較大差異(圖2)。TC1-Mariner超家族(TSD:TA)存在于本研究所有的魚類基因組中, 其中斑馬魚的TC1-Mariner全長拷貝數最多, 有42501個(占基因組中MITEs序列的48.11%), 紅鰭東方鲀的TC1-Mariner全長拷貝序列最少, 只有19條(3.8%)。PHIS超家族(TSD: TWA, CTWAG, CWG)存在29個物種中, 海七鰓鰻的PHIS全長拷貝數最多, 為12868個(20.44%)。P超家族(TSD: 7—8 bp, TIR:CANRG)存在于30個魚類物種中, 斑馬魚P超家族的全長拷貝最多, 為22414個(25.37%)。Kobolok超家族(TSD: TTAA, TIR: RG)存在于23個魚類物種中, 墨西哥麗脂鯉的Kobolok超家族全長拷貝最多,為4647個(8.69%)。PiggyBac超家族(TSD: TTAA,TIR: CMY)存在于27個物種基因組, 日本青鳉全長拷貝最多, 有16951個(70.74%)。hAT超家族(TSD:5—8 bp, TIR: YARNG)除了象鯊和斑點叉尾鮰以外, 存在于31個物種基因組中, 全長拷貝數最多的是斑馬魚, 有6586個(7.46%)。Ginger超家族(TSD:4—5 bp, TIR: TGT)存在于除了半滑舌鰨以外的32個物種基因組中, 大西洋鮭的Ginger拷貝數最多,為4320個(6.39%)。其他的MITEs超家族, 比如Merlin, 只存在于犀角金線鲃的基因組, 拷貝數為547個(7.51%)。Sola2超家族只存在于海七鰓鰻、蒲氏盲鰻、猬鰩三個物種基因組中, 拷貝數最多的是海七鰓鰻, 有3084個(4.90%)。除了上述的MITEs超家族, 還有部分MITEs拷貝由于TSD和TIR無任何規律, 無法鑒定出家族, 全部歸為unknown。總而言之, 魚類基因組中TC1-Mariner超家族占比最大, 與之前的研究結果相符合[16]。
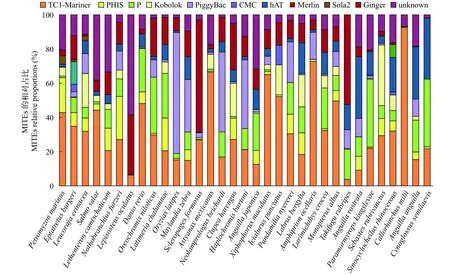
圖2 33種魚類基因組中MITEs超家族分布情況Fig. 2 Distributions of MITEs superfamily in the genome of the 33 fish species
2.3 MITEs插入時間估計
根據K2P模型[26]和公式T=K/2r[27], 使用軟件DAMBE計算不同全長拷貝與一致序列之間的分化度(K), 并用perl腳本統計各物種中MITEs的插入時間。結果表明, 大多數魚類的MITEs全長拷貝(79.58%)插入基因組的時間約為4百萬年前, 也有少數的MITEs全長拷貝插入時間更為古老(20.42%,圖3)。
在魚類進化過程中, 基因組的轉座子數量也在發現改變, 從得到的MITEs插入時間結果分析,7.5百萬年前大部分魚類基因組中的MITEs拷貝較少, 隨著時間的推移, 部分物種出現了MITEs數量激增的現象, 魚類基因組的MITEs插入時間相對更集中在2百萬—0.5百萬年前, 如日本七鰓鰻、海七鰓鰻、猬鰩、矛尾魚、斑點雀鱔、斑馬魚、小丑魚、貝氏隆頭魚在此期間都有一個MITEs集中擴增的表現。其中斑馬魚的MITE拷貝數就曾在7.5百萬—6.5百萬年前有短暫的數量上升, 到6.5百萬—6百萬年前出現MITEs數量減少(僅有1889個全長拷貝), 而后的4百萬年間, 斑馬魚的MITEs數量又緩慢的開始增加, 直到2百萬—1.5百萬年前達到一個擴增的峰值(11596個全長拷貝), 到近期MITEs數量又逐步降低。同樣, 大西洋鮭魚的MITEs數量也是緩慢的上升, 從7.5百萬—7百萬年前的435個MITEs全長拷貝, 經過5百萬年, 到2.5百萬—2百萬年前時達到數量峰值(10841個全長拷貝), 然后MITEs序列又逐漸減少。不同的是, 有些物種在進化過程中,MITEs會經歷兩次擴增“爆發”期(拷貝數量達到峰值, 而后下降), 比如墨西哥麗脂鯉、斑馬魚、羅非魚、象鼻魚和犀角金線鲃。除此之外的其他物種都僅有一次MITEs擴增“爆發”。
2.4 MITEs在基因組中的分布
魚類基因組中的MITEs轉座子具有較高的豐度, 為了保證結果的準確性, 僅選取有完整TIR的MITEs全長拷貝進行插入位置分析。結果顯示(表2),魚類基因組的MITEs全長拷貝插入到基因內部的數量十分豐富(計算插入時, 以轉座子所處位置為準, 較長的轉座子可能出現重復計數的現象, 對結果無影響), 例如斑馬魚插入到基因的有12885個。其中矛尾魚、斑點雀鱔、象鼻魚、大西洋鯡、斑馬魚、犀角金線鲃、墨西哥麗脂鯉、斑點叉尾鮰、大西洋鮭魚、紅鰭東方鲀、黃鱔、半滑舌鰨、貝氏隆頭魚、大黃魚、羅非魚、布氏新亮麗鯛、斑馬宮麗魚、紅麗魚、伯氏樸麗魚、小丑魚、花斑劍尾魚、弗氏假鰓鳉和日本青鳉基因組中的MITEs都有插入到外顯子的情況。對這部分基因進行注釋, 發現象鼻魚、斑馬魚、墨西哥麗脂鯉、弗氏假鰓鳉、黃鱔、大黃魚、小丑魚、羅非魚的MITEs都插入到能夠編碼具有鋅指結構的蛋白(Zinc finger)的序列中, 這些序列編碼的蛋白大多與基因表達相關; 墨西哥麗脂鯉有一條MITEs插入到能編碼視蛋白對應的序列中; 斑點雀鱔的有一條MITEs插入到編碼脂肪包被蛋白(與肥胖相關)的序列中等等。但后續還需要更詳細的分析和實驗驗證, 才能明確MITEs是否對基因表達有影響。
3 討論
魚類占現有脊椎動物總數的50%以上, 它豐富的物種多樣性, 廣闊的物種分布以及古老的進化地位都使它成為生物進化歷史上不可或缺的一部分。隨著測序技術的發展, 越來越多的魚類基因組數據被公布, 基因組中的轉座子序列也得到了生物學家的關注, 目前已完成52種魚類的轉座子鑒定工作[21], 魚類特有的轉座子數據庫也已經發布[34]。MITEs屬于非自主型的DNA轉座子, 已有研究表明MITEs起源于自主DNA轉座子, 并且其在基因組中存在豐富的拷貝[35], MITEs的插入對“宿主”的基因功能也會造成影響[36—38]。我們使用de novo預測和結構預測相結合的方法對33種魚類基因組進行MITEs轉座子預測, 然后用Bioedit手工鑒定, 有效降低了預測結果的假陽性, 為后續插入時間和插入位置分析提供保障。
魚類基因組中MITEs含量差異較大(0.11%—21.18%), 比植物基因組(0.01%—9.98%)[10]和昆蟲基因組(0.00%—6.24%)[12]中的MITEs含量更高。MITEs在基因組中的拷貝數較多, 斑馬魚中全長拷貝可達88337個, 其原因可能有兩個, 一方面, 輻鰭魚綱的魚類比軟骨魚綱的含有更多DNA轉座子; 另一方面, 斑馬魚是模式物種, 基因組質量高(基因組組裝到染色體, Contig N50為854399 bp), 預測結果也更好。在本研究中, 猬鰩基因組鑒定的MITEs含量很低(0.11%), 這可能與軟骨魚基因組中DNA類轉座子總體含量偏低及基因組組裝質量不高(基因組Contig N50為665 bp)有關。同時, 發現魚類基因組大小和基因組中MITEs含量呈正相關(r=0.124,P<0.05), 與植物和昆蟲的情況類似[10,12], 因此可知MITEs的高拷貝數、高含量對基因組大小的進化有一定貢獻。而魚類基因組中的MITEs超家族主要是TC1-Mariner (TSD: TA), 它在植物[10,30]、昆蟲[12]、病毒[14]等基因組中都含有, 也是最初發現的MITEs之一[30], 在MITEs含量中占主導地位。

圖3 33種魚類基因組中MITEs插入時間估計Fig. 3 MITEs insertion time estimation in the 33 fish genomes
MITEs與其他轉座子不同, 它在基因組中傾向插入到基因內部或附近(基因側翼5 kb區域內)[39],本研究結果顯示, 大部分MITEs全長拷貝都插入到“宿主”的基因內部, 符合MITEs的插入特征。但MITEs的插入可能會使基因表達發生變化, 例如水稻的mPing轉座子插入Rurm1基因的內含子中, 使水稻穎葉更加細長[40]。在魚類基因組中, 很多MITEs序列插入到基因的內含子, 甚至有23種魚類的MITEs序列插入到基因的外顯子, 這在昆蟲和植物中很少見。所以, 還需要對插入到基因的MITEs序列進行深入研究, 確定它們對“宿主”的基因表達和功能是否有影響。
MITEs能夠以“剪切-粘貼”模式高效率轉座[35],依靠對應的自主轉座子編碼的轉座酶, 在短時間內增加拷貝數。MITEs插入時間分析結果顯示, 魚類基因組的MITEs大多在2百萬—0.5百萬年前出現了一次擴增“爆發”期, 這期間MITEs的數量快速增加并達到峰值, 之后數量又逐漸減少。轉座子“生命周期(Lifecycle)”假說[41]認為, 轉座子進入“宿主”后有一個快速擴增時期, 這期間轉座子拷貝數急劇增加, 同時產生突變導致部分轉座子失活。接著進入成熟期, 轉座子拷貝數增加和丟失的速率達到平衡。最后進入衰老期, 自主型和非自主型轉座子丟失, 不再擴增, 衰老期持續的時間最長可達幾百萬年。我們得到的MITEs擴增結果與這個假說相符合, 這種情況也說明MITEs雖然能通過轉座增加拷貝, 但并不能一直擴增下去, 若這種擴張給“宿主”帶來負面影響, “宿主”會有一些沉默機制阻止MITEs擴增, 如piRNA(非編碼小RNA)通過誘導特殊位點的組蛋白修飾能對轉座子轉座進行有效抑制[42]。若這類MITEs對轉座子有利, 則會繼續留存在基因組中, 例如斑馬魚基因組中MITEs序列有兩次擴增的跡象。因此在魚類基因組中, MITEs的擴增會影響基因組大小, 同時“宿主”也有相應的機制控制MITEs的數量, 而MITEs在基因組中的插入會對物種的進化、基因的結構和功能產生影響, 因此鑒定魚類的MITEs是研究魚類基因組中轉座子功能的基礎。

表2 MITEs在魚類基因組中的分布Tab. 2 Distributions of MITEs in the fish genomes
4 結論
對33種魚類基因組的MITEs進行預測、鑒定和分類, 發現魚類的MITEs含量差異顯著, MITEs含量與魚類基因組大小呈正相關。在33種魚類中, 均含有大量TSD為TA的MITEs序列, 與昆蟲、植物類似。魚類基因組中的MITEs插入時間大多在4百萬年前, 更傾向于插入在基因內部, 這可能會對基因表達調控產生影響。我們闡述了魚類基因組中MITEs的分布情況, 為后續魚類基因組中MITEs功能的研究奠定了基礎。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
