日本沼蝦肌抑素前體肽在枯草芽孢桿菌的表達及其對生長效果的影響
2020-03-04 03:15:48章曉棟沈文英
水生生物學報 2020年1期
章曉棟 沈文英 任 崗
(紹興文理學院生命科學學院生物系, 紹興 312000)
肌抑素(Myostatin, MSTN)是骨骼肌生長發育的負調控因子[1], 抑制成肌細胞的增殖和分化[2]。肌抑素前體蛋白由信號肽、編碼前體肽的N-末端區和編碼成熟肽的C-末端區三部分組成, 經兩步蛋白酶切轉化成成熟的肌抑素[3,4]。肌抑素前體肽(Myostatin propetide, MSTNpp)在酶切分泌后通過非共價相互作用, 與成熟肽間形成復合物, 抑制成熟肽與其受體的結合, 阻斷肌抑素的信號轉導通路,從而抑制肌抑素生物活性, 促進肌肉生長[5]。研究表明, 過表達肌抑素前體肽或將肌抑素前體肽注入小鼠均可有效促進肌肉生長, 表現出骨骼肌的“雙肌”現象[6,7]。在斑馬魚胚胎發育中過表達肌抑素前體肽, 骨骼肌肌纖維數量顯著增加, 胚胎體型大、生長加快[8]。相對于脊椎動物中肌抑素及前體肽的深入研究, 無脊椎動物肌抑素的研究較少。目前只在海灣扇貝(Argopecten irradians)[9]、斑節對蝦(Penaeus monodom)[10]、大西洋龍蝦(Homarus americanus)[11]、日本長額蝦(Pandalopsis japonica)[12]、日本沼蝦(Macrobrachium nipponenese)[13]等物種克隆得到肌抑素基因。研究表明, 在蛻皮階段,大西洋龍蝦、日本長額蝦、日本沼蝦等肌抑素基因表達下調, 促進間歇性快速生長[11—13]。而關于無脊椎動物肌抑素前體肽的研究尚未見報道。
日本沼蝦(M. nipponenese)是我國養殖面積最廣、產量最大的淡水經濟蝦類。在日本沼蝦養殖中, 存在生長緩慢、個體小型化、性早熟等現象[14],導致生長速度和生產性能低下, 養殖總產量不高。目前, 主要通過遺傳育種和飼料研發等手段改良日本沼蝦性狀, 提高養殖規格和效益[15,16]。本研究從日本沼蝦肌肉生長調控角度探討促進生長的途徑,在前期日本沼蝦MSTN全基因的克隆和表達特性研究[13]基礎上, 采用枯草芽孢桿菌表達體系表達日本沼蝦肌抑素前體肽, 分析其對生長的影響, 為闡明日本沼蝦肌抑素前體肽對肌肉生長發育的調節機理及其在養殖中的應用提供實驗數據。
1 材料與方法
1.1 材料
大腸桿菌DH5α、枯草芽孢桿菌WB800, 均為本實驗室保存。穿梭分泌性表達載體pGJ105, 為本實驗室構建。
1.2 穿梭分泌性表達載體的構建
根據日本沼蝦MSTNpp原始基因序列特征, 按照枯草芽孢桿菌的偏愛密碼子對MSTNpp基因序列進行優化合成(上海生工, 上海)BsMSTNpp, 并亞克隆到pET28a(NdeⅠ/XhoⅠ), 獲得質粒pET28a-BsMSTNpp。根據優化合成基因全長序列, 設計表達引物, 上游引物為P001CF: 5′-GGCAACCGCCTCT GCAGCGCAGAAAAGCCACGGGAAAAC-3′, 下游引物為3pETR: 5′-TCATTAAGCTTGTCGG GATCCGTTAGCAGCCGGATCTCAGTGG-3′, 下劃線為同源重組片段。以質粒pET28a-BsMSTNpp為模板, 目的基因全長序列951 bp, 反應體系為50.0 μL。PCR反應條件: 98℃, 5min; 98℃,10s;55℃, 10s; 72℃, 60s; 5個循環; 98℃, 10s; 58℃, 30s;72℃, 60s; 30個循環; 72℃延伸10min, 并進行PCR產物切膠回收。
枯草芽孢桿菌的表達載體pGJ105用PstⅠ和BamHⅠ進行雙酶切, 酶切產品切膠回收, 將純化后的PCR產物2.5 μL與載體酶切產物pGJ105(PstⅠ-BamHⅠ)2.5 μL、Gibson Assembly Master Mix(2×,NEB, E5510S)5 μL, 置于PCR儀中50℃反應15min,進行同源組裝, 連接產物轉化DH5α, 篩選并測序獲得重組質粒pGJ105-BsMSTNpp。
1.3 MSTNpp基因在枯草芽孢桿菌中的表達及鑒定
通過二步法[17]制備枯草芽孢桿菌感受態細胞。將構建成功的重組質粒pGJ105-BsMSTNpp轉化進入枯草芽孢桿菌WB800感受態細胞中; 通過PCR和酶切鑒定, 挑選陽性轉化子。將陽性克隆轉接到發酵培養基(SpcR, 100 ng/mL)中, 在30℃, 180 r/min培養至24h, 收集上清液, 以含pGJ105空質粒的WB800菌株作為對照, 通過Western blot檢測目的蛋白的表達。
取不同時間的發酵液上清進行Western blot分析, 檢測重組BsMSTNpp的表達和分泌的時間動態情況。
1.4 MSTNpp重組枯草芽孢桿菌菌劑和飼料的制備方法
重組枯草芽孢桿菌接種于枯草發酵培養基中,30℃, 180 r/min培養24h離心取菌泥, 37℃恒溫干燥至恒重, 與白炭黑、玉米淀粉按1:0.5:4的質量比混合制成干菌粉, 干菌粉中重組枯草芽孢桿菌活菌數為109cfu/g。將枯草芽孢桿菌工程菌按照日本沼蝦餌料的0.5‰、1‰的比例加入日本沼蝦餌料中, 用α淀粉粘合、攪拌均勻, 風干, 再噴灑5‰的橄欖油包被, 風干后4℃保存備用。
1.5 MSTNpp枯草芽孢桿菌投喂日本沼蝦養殖實驗
選擇平均體重為(1.52±0.11) g, 平均體長為(4.55±0.21) cm的健康日本沼蝦, 隨機分為4組, 每組3個重復, 每個重復200尾, 養殖于體積為1 m3的水泥池中, 實驗期間保持養殖水溫為(24±3)℃, 日投喂量為蝦體重的2%, 試驗開始前暫養1周。在日本沼蝦基礎飼料中添加0.5‰和1‰重組枯草芽孢桿菌菌劑, 分別作為實驗組1和實驗組2; 投喂無任何添加的日本沼蝦基礎飼料, 作為對照組1, 在日本沼蝦基礎飼料中添加1‰空白枯草芽孢桿菌生物制劑(不表達MSTNpp的空白枯草芽孢桿菌), 作為對照組2;實驗周期為30d。實驗開始前和結束時測定日本沼蝦體重, 計算特定生長率(Special growth rate,SGR)和增長率。SGR(%)=(LnWt-LnW0)/T×100%; 其中Wt,W0分別表示平均終末體重(g)、平均初始體重(g);T表示飼養時間(d)。增長率(%)=[(實驗組或對照組2SGR-對照組1SGR)/對照組1SGR]×100%。
實驗結束取日本沼蝦肌肉組織, 稱重, 按1﹕9的比例加入PBS, 冰浴條件下勻漿, 制備成10%的組織勻漿, 2500 r/min, 離心10min, 取上清測定肌酸激酶活性, 肌酸激酶(CK)試劑盒購自南京建成生物科技有限公司(南京)。
1.6 數據處理
實驗數據采用平均值±SD表示, 利用DPS軟件處理數據,P<0.05表示存在顯著性差異。
2 結果
2.1 BsMSTNpp表達載體的構建
以質粒pET28a-BsMSTNpp為模板, P001CF(PstⅠ)和3pETR(BamHⅠ)為引物擴增優化好的BsMSTNpp基因, 經瓊脂糖凝膠電泳檢測, 目的條帶大小約1000 bp, 與預期結果(951 bp)相符。采用PstⅠ和BamHⅠ雙酶切pGJ105質粒, 分別回收酶切產物和PCR片段, 通過同源組裝連接后轉化, 雙酶切和PCR鑒定陽性重組子, 送陽性克隆進行測序鑒定, 測序正確得到BsMSTNpp重組表達質粒pGJ105-BsMSTNpp (圖1)。
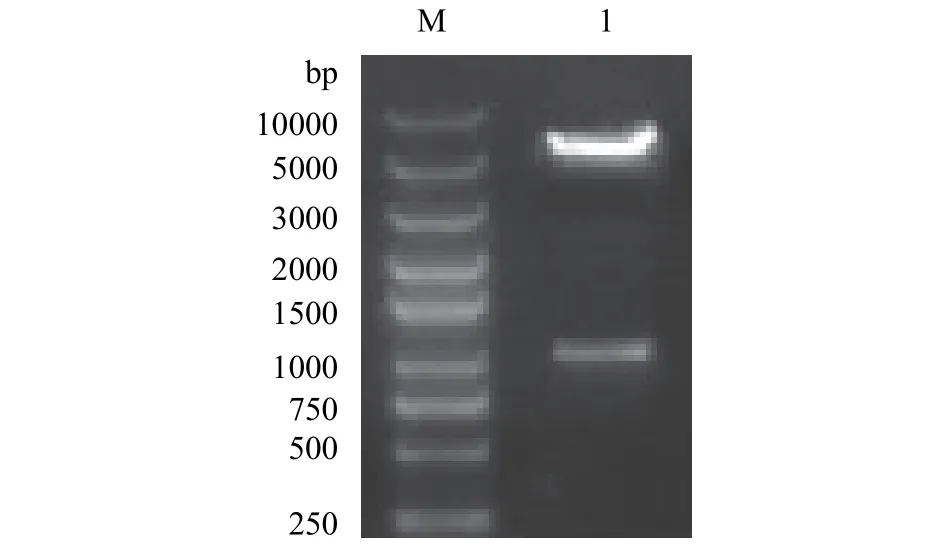
圖1 重組質粒pGJ105-BsMSTNpp和目的基因PCR電泳Fig. 1 PCR of recombined pGJ105-BsMSTNpp and BsMSTNpp gene
2.2 BsMSTNpp基因在枯草芽孢桿菌中的表達
將重組質粒pGJ105-BsMSTNpp轉化到枯草芽孢桿菌宿主菌WB800, 培養24h后, 收集上清液進行Western blot鑒定, 結果顯示陽性菌上清中有與預期大小相近的條帶, 由于融合了His.Tag的一段序列,所以重組蛋白理論分子量約36.0 kD (圖2)。
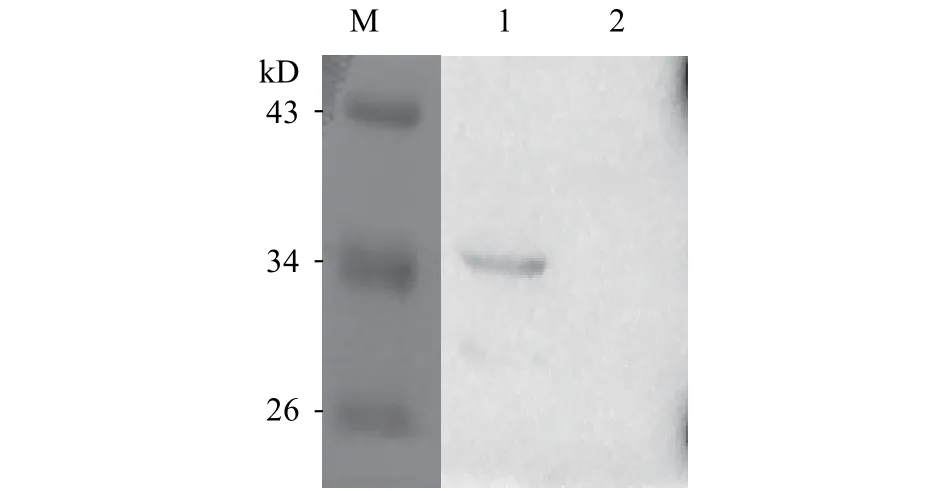
圖2 重組菌發酵上清Western blot鑒定Fig. 2 Western blot analysis of the supernatant samples
2.3 BsMSTNpp基因在枯草芽孢桿菌分泌表達時間動態
取不同時間的發酵液上清進行Western blot鑒定分析, 檢測重組BsMSTNpp的表達和分泌情況(圖3)。在發酵液的上清中, 在培養24h后開始出現重組BsMSTNpp的表達條帶, 從條帶的灰度分析可見, 重組BsMSTNpp隨著時間的延長, 其表達量逐漸在升高。在檢測的時間范圍內, 在分泌上清中100h后達到最高值, 重組BsMSTNpp的表達量相對于24h約提高了10倍。
2.4 肌抑素前體肽對日本沼蝦生長和CK酶活力的影響
養殖實驗結束后測定日本沼蝦特定生長率, 對照組1、對照組2、實驗組1和實驗組2日本沼蝦SGR分別是2.88%、3.00%、3.17%和3.31% (表1)。添加0.5‰和1‰表達MSTNpp的重組枯草芽孢桿菌的實驗組日本沼蝦SGR顯著高于對照組1(P<0.05),分別提高10.1%和14.9%; 實驗組日本沼蝦SGR均高于對照組2, 但不存在顯著性差異。日本沼蝦肌酸激酶活性測定結果表明, 實驗組1和實驗組2的CK酶活性顯著高于對照組(P<0.05), 表明重組表達MSTNpp枯草芽孢桿菌能有效提高日本沼蝦CK酶活性。推測重組表達的MSTNpp可能通過促進肌細胞增殖和分化提高肌肉組織生長, 從而進一步影響日本沼蝦生長速度。

圖3 不同發酵時間WB800 [pGJ105-BsMSTNpp]上清中BsMSTNpp的Western blot分析Fig. 3 Western blot analysis for recombined BsMSTNpp expression
3 討論
枯草芽孢桿菌(B. subtilis)具有較清楚的遺傳背景、良好的分泌性和無致病性等優點, 將其芽孢制備成口服制劑能夠耐受胃腸道環境, 目前已作為重要工業菌種應用于水產、畜牧養殖業, 被批準為食品級安全菌株[18]。和大腸桿菌表達系統相比, 枯草芽孢桿菌胞壁不含內毒素, 簡化了表達產物的純化和回收; 和酵母表達系統相比, 枯草芽孢桿菌培養成本低, 發酵周期短[19,20]。但枯草芽孢桿菌表達系統存在一個限制因素, 即生長到對數末期時會分泌表達多種胞外蛋白酶, 導致目的蛋白降解。Bol-huis等[21]利用基因失活的突變方法, 成功構建一株在野生菌株的基礎上缺失了8個蛋白酶基因的枯草芽孢桿菌WB800, 該蛋白酶缺陷菌株胞外蛋白酶的活力只有野生型的0.32%, 克服了表達蛋白降解及不穩定等問題。近年來, 隨著枯草芽孢桿菌表達系統的完善, 越來越多地應用于飼用酶、飼用代謝產物、抗菌肽、藥用蛋白、抗原蛋白的生產[22—25]。由于大部分枯草芽孢桿菌不含內源性質粒, 體外連接的雙鏈質粒DNA分子直接轉化宿主效率很低, 因此構建大腸桿菌-枯草芽孢桿菌穿梭載體十分必要[26—28]。本研究根據枯草芽孢桿菌表達系統的特點, 對日本沼蝦MSTNpp基因序列進行了優化, 同時為提高轉化效率, 構建了大腸桿菌-枯草芽孢桿菌穿梭載體pGJ105, 構建重組表達載體pGJ105-BsMSTNpp, 成功地在枯草芽孢桿菌WB800中表達了日本沼蝦肌抑素前體肽基因, 經過Western blot檢測到大小為36.0 kD的表達產物。本研究采用了組合型啟動子PrapA, 不需要誘導物, 重組蛋白可以持續表達[27], 通過分析發酵時間對重組BsMSTNpp表達量的影響, 結果表明發酵時間與重組BsMSTNpp表達量成正相關。
為驗證表達產物的生物活性, 本實驗將表達有MSTNpp的枯草芽孢桿菌工程菌添加到日本沼蝦飼料進行投喂。結果表明, 枯草工程菌能夠有效提高日本沼蝦的肌酸激酶活性和特定生長率, 促進日本沼蝦生長。大量研究表明枯草桿菌可以提高草魚、凡納濱對蝦等水產養殖動物的生長性能[28—31]。在本研究中, 實驗組日本沼蝦特定生長率顯著提高,但和對照組2間不存在顯著性差異, 可能與養殖周期不夠長有關。在小鼠和斑馬魚的研究表明, 過表達肌抑素前體肽可以有效促進骨骼肌細胞的增殖和生長[6—8]。CK又叫磷酸肌酸激酶(Creatine phosphokinase, CPK), 催化肌酸轉換成磷酸肌酸, 消耗三磷酸腺甘(Adenosine triphosphate, ATP)生成二磷酸腺苷(Adenosine diphosphate, ADP), 是肌肉組織代謝的關鍵酶, 在成肌細胞增殖和分化過程中能快速合成肌酸激酶[32], CK已成為直接測定肌細胞增殖和分化能力的特異性指標[33,34]。本研究結果表明重組表達MSTNpp枯草芽孢桿菌能有效提高日本沼蝦肌酸激酶活性, 證明日本沼蝦肌細胞的增殖和分化能力增強了。推測重組表達的MSTNpp可能通過促進肌細胞增殖和分化提高肌肉組織生長,從而進一步影響日本沼蝦生長速度。
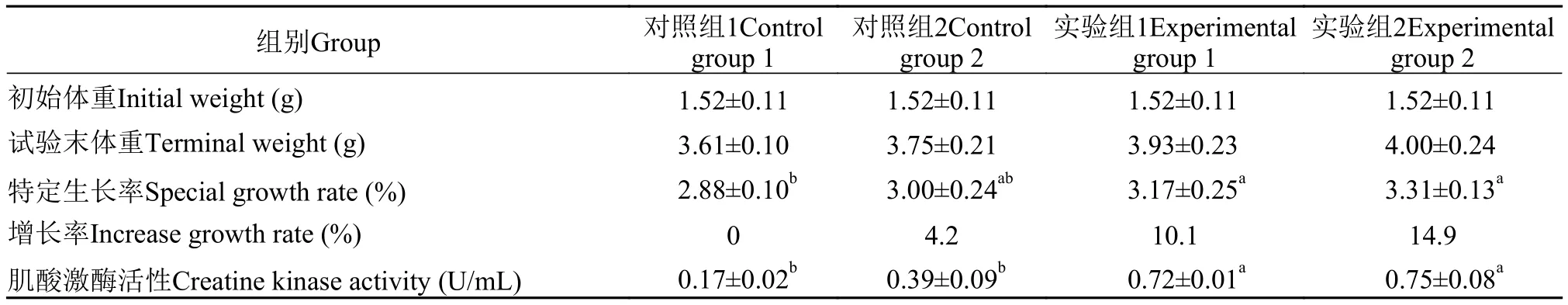
表1 MSTNpp重組蛋白對日本沼蝦生長率和肌酸激酶活性的影響Tab. 1 Effects of recombined MSTNpp on growth rate and creatine kinase activity
4 結論
本實驗采用枯草芽孢桿菌系統對日本沼蝦MSTNpp進行表達, Western blot檢測到重組MSTNpp蛋白分子量約為36.0 kD。為進一步研究MSTNpp對日本沼蝦生長的作用機理提供了基礎。將重組枯草芽孢桿菌投喂日本沼蝦30d, 驗證表達產物的生物活性, 結果證明重組表達MSTNpp枯草芽孢桿菌能提高日本沼蝦CK酶活性, 促進肌細胞增殖和分化, 進一步影響生長速度。為肌抑素前體肽進一步應用于日本沼蝦養殖提供了實驗基礎。
