綏芬河大麻哈魚個體生物學研究
2020-03-04 03:16:06王繼隆王維坤李培倫楊文波
水生生物學報 2020年1期
王繼隆 劉 偉 王維坤 李培倫 楊文波,
(1. 中國水產科學研究院黑龍江水產研究所,哈爾濱 150070; 2. 中國水產科學研究院農業農村部漁業遙感科學觀測實驗站,北京 100141; 3. 黑龍江省東寧市鮭魚孵化放流站 , 東寧 157200; 4. 中國水產科學研究院資源與環境研究中心,北京 100141)
大麻哈魚(Oncorhynchus ketaWalbaum)是典型的溯河洄游性魚類, 其廣泛分布于北太平洋及沿岸國家的淡水河流中。綏芬河大麻哈魚是我國主要的四個大麻哈魚地理種群之一[1], 具有重要的經濟和科學價值。近幾十年來, 由于過度捕撈、棲息地破壞等人為因素的影響, 大麻哈魚資源衰退嚴重[2]。為了保護和恢復大麻哈魚資源, 自20世紀80年代在綏芬河水域開展大麻哈魚增殖放流工作[3], 目前綏芬河大麻哈魚維持在一定的種群數量。作為一種典型洄游性魚類及海洋放牧魚類, 大麻哈魚的研究工作一直備受關注。國外學者對大麻哈魚的研究較為深入, 有關大麻哈魚資源量變動、繁殖活動及海、淡水中的生長情況等都有相應報道[4—6], 近年來國內學者在大麻哈洄游群體數量結構與生物學方面開展了一些研究[7—10], 但是主要針對黑龍江、烏蘇里江大麻哈魚地理種群。董崇智等[3,11,12]學者曾經對綏芬河水域大麻哈魚生物學特征開展過相關研究, 但是近十幾年對該種群的研究未再見報道。
生物學特征的研究是了解魚類生長情況、種群動態和資源變化的基礎, 為漁業資源管理提供數據資料和科學依據。目前綏芬河大麻哈魚正面臨過度捕撈、環境變化等負面因素以及增殖放流工作帶來的多重因素的綜合影響。大麻哈魚種群在長期的生存壓力之下, 其生物學指標等表型特征是否會發生變化,值得我們關注。鑒于此, 本文基于2012—2017年在綏芬河水域開展大麻哈魚回歸群體的調查監測, 對其年齡、生長、繁殖生物學等個體生物學特征進行研究分析, 以期為綏芬河大麻哈魚的增殖、保護工作提供基礎資料。
1 材料與方法
1.1 樣本采集
本研究中大麻哈魚樣本于2012至2017年每年9—11月采捕于綏芬河東寧段(圖1), 采捕方式為定置刺網(網目大小10 cm)。對捕獲的大麻哈魚進行現場測量叉長、稱量體質量、取鱗片, 采集樣本低溫保存帶回實驗室進一步分析, 本研究共采集到大麻哈魚樣本447尾。
1.2 材料處理及數據分析
采用鱗片進行年齡鑒定, 鱗片前處理、年齡鑒定及鱗徑測量方法參照王繼隆等[9]的方法。大麻哈魚的叉長退算公式如下:
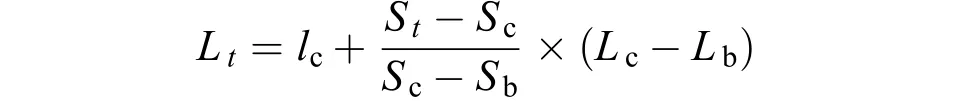
式中,Lc和Sc為大麻哈魚捕獲時的叉長和鱗片鱗徑,Lb和Sb為鱗片形成時的叉長和鱗徑,Lt和St為t齡時叉長和鱗徑。根據以往研究結果[13], 式中Lb=4 cm;Sb=0.114 cm。
參考王繼隆等[9]研究結果, 采用Von Bertalanffy生長方程(VBGF)擬合大麻哈魚的叉長生長, 公式如下:
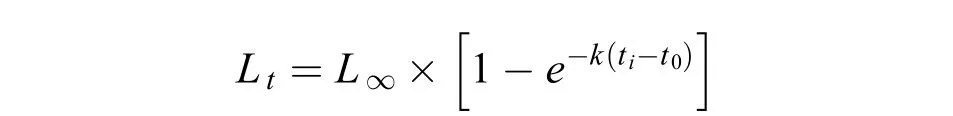
式中,L∞、k、t0為生長方程參數,L∞為漸近叉長,k為生長曲線的平均速率,t0為假設的理論生長起點年齡。生長模型的參數采用最大似然法估計。
采用殘差平方和(ARSS)方法分析判斷雌、雄個體叉長生長差異的顯著性[14]。
采用叉長生長特征指數來衡量大麻哈魚的生長速度, 計算公式如下:
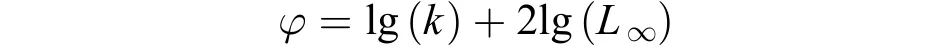
以2 cm為間距, 利用各叉長組的性成熟個體百分比擬合Logistic 曲線, 推算大麻哈魚的初次性成熟叉長概率分布, 計算50%初次性成熟叉長(L50)。根據資料, 將大麻哈魚最后一個年輪形成時期作為其性成熟的劃分依據[15]。
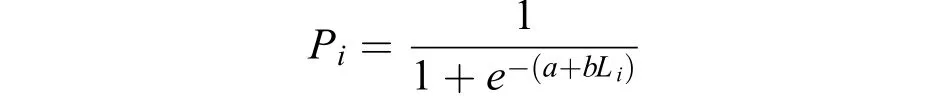
式中,Pi為成熟個體占組內樣本的百分比;Li為各叉長組(cm)。初次性成熟叉長為:L50= -a/b。
由于大麻哈魚溯河洄游期間停食空胃, 性腺成熟指數(Gonad somatic index,GSI)計算公式中分母采用體質量(W)。公式如下:
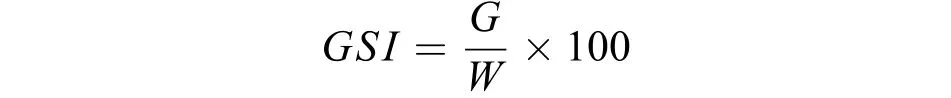
式中,G為性腺重,W為體質量。
絕對繁殖力F(粒)=樣品卵粒數/樣品重量×性腺重; 叉長(L)相對繁殖力(粒/cm)FL=F/L; 體重相對繁殖力(粒/g)FW=F/W。
2 結果
2.1 體長、體重與年齡組成
對447尾大麻哈魚樣本進行生物學分析(表1),雌性大麻哈魚叉長為: 49.2—79.9 cm, 平均為64.7 cm,65—67 cm為優勢叉長組; 體重為: 1250.1—5499.8 g,平均為2943.9 g。雄性大麻哈魚叉長為: 43.5—78.2 cm,平均為55.5 cm, 52—54 cm為優勢叉長組; 體重為:801.5—5348.9 g, 平均為1857.3 g。雌、雄大麻哈魚樣本年齡組均分為1+—5+齡五個年齡組。其中雌性個體以3+齡為主, 占總樣本總數的60.64%, 其次為4+和2+齡, 1+齡數量最少, 平均年齡為4.07齡; 雄性個體以2+齡個體為主, 占總數的46.46%, 其次為3+和1+齡, 5+齡數量最少, 平均年齡為3.15齡。可見,雌性個體叉長、體重、年齡均比雄性個體大。
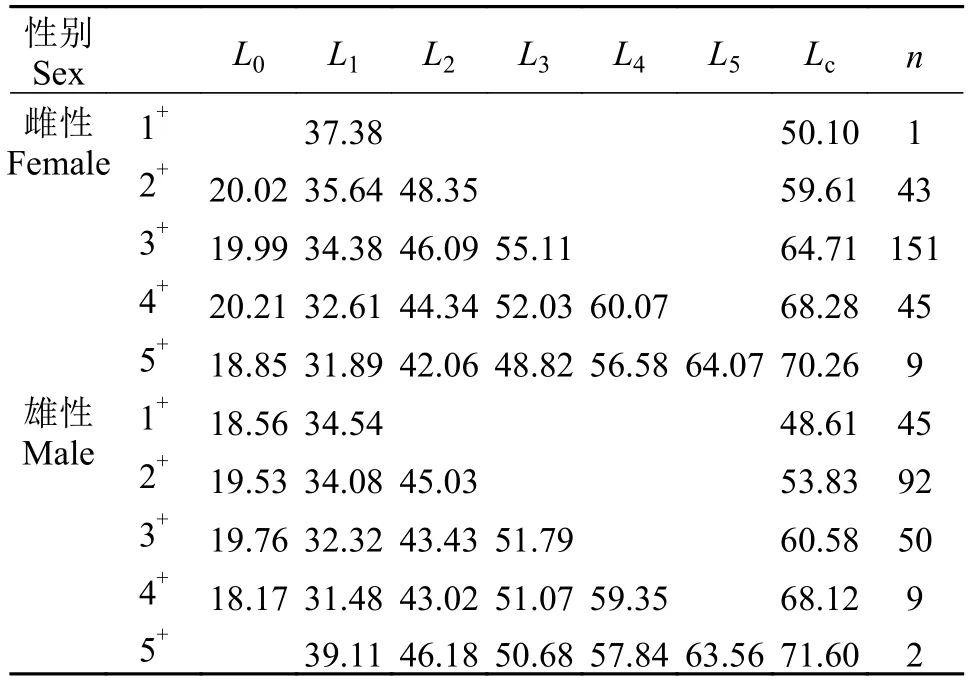
表1 綏芬河大麻哈魚的實測叉長與退算叉長Tab. 1 Actual fork length and back-calculated fork length of chum salmon from the Suifen River
2.2 生長
大麻哈魚雌、雄個體的體長-體重關系式分別為:W=0.0082×L3.0604;W=0.0076×L3.0746(W為體重, g;L為叉長, cm)。t檢驗表明兩關系式中的參數b均與3無顯著差異[t雌= 0.45<t(0.05, 244);t雄= 1.03<t(0.05, 194)], 表明大麻哈魚的生長屬于勻速生長,可采用VBGF進行生長模擬, 生長方程參數見表2。以3+齡大麻哈魚為例, 采用ARSS分析雌、雄大麻哈魚VBGF生長的差異, 結果表明綏芬河3+齡大麻哈魚雌、雄個體間生長差異顯著(F= 13.91 >F0.01,2,5),故將雌、雄大麻哈魚的叉長生長分別進行分析。
由表2可見, 大麻哈魚叉長生長指標隨著年齡的增加而遞減, 即年齡越大生長速度越慢。大麻哈魚2+齡組雌、雄性個體的生長速度相同, 3+齡組雌性個體生長速度高于雄性個體。為分析大麻哈魚各年齡段的生長情況, 分析叉長的生長速度。以3+齡組雌、雄大麻哈魚為例, 在叉長生長方程的基礎上分別對時間t求一階導數, 即得到相應的生長速度方程:
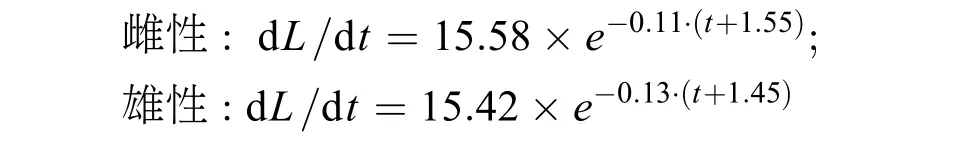
根據生長方程繪制了大麻哈魚叉長生長及生長速度曲線(圖2), 可見, 大麻哈魚雌性個體的生長速度快于雄性個體, 雌、雄個體叉長生長速度隨年齡的增長均呈遞減趨勢。
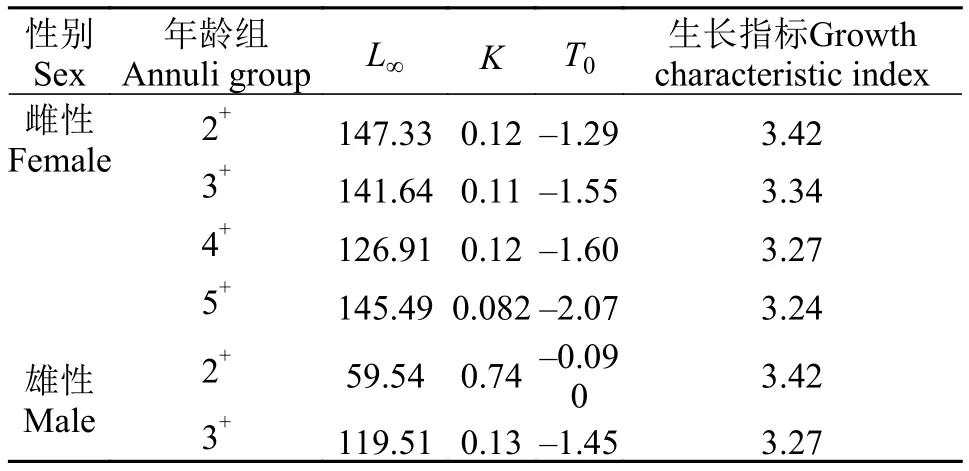
表2 不同年齡組大麻哈魚的生長Tab. 2 Growth of chum salmon from different age groups
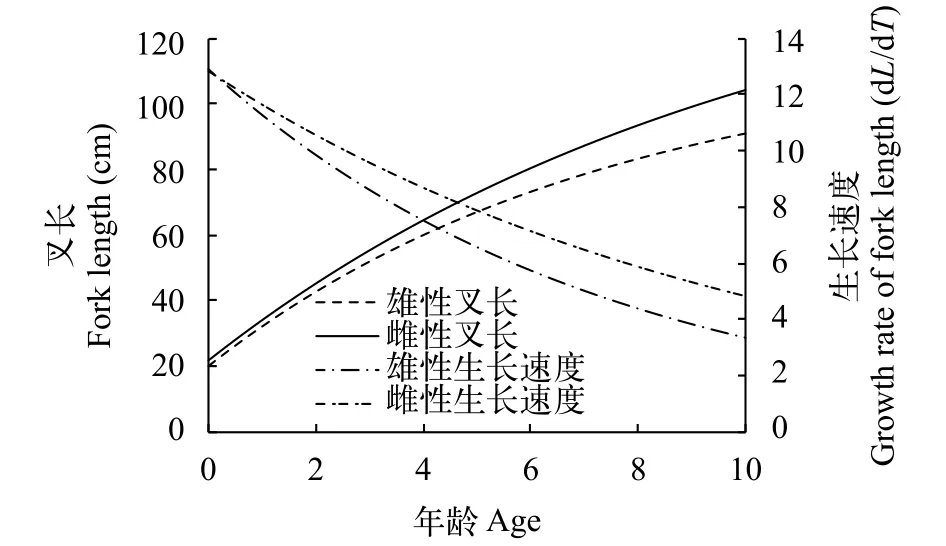
圖2 大麻哈魚的叉長生長曲線和生長速度曲線Fig. 2 The growth curve and the growth rate curve of fork length of chum salmon
2.3 繁殖生物學
50%初次性成熟叉長(L50)根據logistic方程計算綏芬河大麻哈魚雌、雄個體的L50分別為51.53和42.15 cm, 對應的年齡分別為2.60和1.91齡(圖3)。ARSS分析表明雌、雄個體間的性成熟概率差異極顯著(F= 49.43 >F(0.01,25,27)), 雄性個體的性成熟期明顯早于雌性。
GSI和絕對繁殖力用于測定繁殖力的大麻哈魚樣本為142尾, 叉長為: 49.2—79.9 cm, 平均叉長為65.0 cm, 體重為: 1250.1—5498.5 g, 平均體重2966.2 g, 年齡范圍為1+—5+齡, 其中3+、4+齡組樣本較多。表3可見, 3+齡組的繁殖貢獻最大, 懷卵量比例達到52.77%, 其次為4+齡組, 占30.82%。
圖4可見, 大麻哈魚個體絕對繁殖力(F)分布在857—4535粒, 平均為3412粒, 其中主要分布在3800—4300粒, 個體絕對繁殖力隨年齡的增長而增加。叉長相對繁殖力(FL)分布在15—70 eggs/cm, 平均為52.42 eggs/cm, 其中主要分布在55—65 eggs/cm;體重相對繁殖力(FW)分布在0.5—1.8 eggs/g, 平均為1.17 eggs/g, 其中主要分布在1—1.5 eggs/g。可見大麻哈魚F與FW分布較集中, 而與FL分布比較分散。
相關分析表明,F與叉長、體重、性腺重成顯著正相關關系;GSI與叉長、體重、F成顯著負相關關系, 其中F和叉長相關關系系數最大;FW與GSI、F和FL成正相關關系。本文采用線性函數和冪指數函數擬合大麻哈魚F與叉長、體重的關系, 結果顯示冪指數函數效果更好, 所得冪指數方程為:F=0.0311×L2.7745(R2=0.638);F=1.946×W0.9374(R2=0.704)。
3 討論
3.1 綏芬河大麻哈魚的生長特征
2012—2017年綏芬河大麻哈魚的雌、雄性個體平均叉長、體重分別為64.7 cm、2943.9 g和55.5 cm、1857.3 g。雌、雄大麻哈魚樣本年齡組均分為1+—5+齡。雌性個體以3+齡為最多, 占總數的60.64%; 雄性個體以2+齡為主, 占46.46%。董崇智等[12]1986年調查綏芬河大麻哈魚的平均叉長、體重為70.2 cm、4150 g, 雌性個體年齡組為3+和4+齡,其中3+齡占72.3%, 雄性個體年齡組為2+—4+齡, 其中3+齡占62.5%。可見綏芬河大麻哈魚的叉長、體重、年齡和20世紀80年代相比均呈減小趨勢。相似的現象在烏蘇里江大麻哈魚種群中也有發現[8,16]。本研究中大麻哈魚的年齡范圍較1986年調查結果有所擴大。主要因為本研究采用六年的調查數據進行分析, 樣本收集時間長, 故樣本年齡組范圍較廣。本調查發現1+齡組的低齡大麻哈魚占總樣本的10.3%, 以往僅見于董崇智等[11]1990年的調查研究中, 但其比例較小(占1.7%)。可見, 性成熟年齡低齡化、規格小型化是目前綏芬河大麻哈魚生物學特征變化的集中體現。大麻哈魚性成熟年齡除遺傳因素外還與海洋環境條件有關[6]。在優越的環境條件(生物與非生物)下大麻哈魚生長速度快, 性成熟年齡提前[6]。然而海洋環境條件的變化具有一定周期性, 幾十年來綏芬河大麻哈魚性成熟年齡的降低主要原因應是漁業捕撈的選擇性。長期的漁業捕撈使得大規格個體死亡率高、小規格個體種群延續率增加是導致大麻哈魚小型化的一個重要因素[8]。
綏芬河大麻哈魚的叉長生長速度與性成熟年齡成反比, 性成熟年齡越小生長指標較大, 這種現象在烏蘇里江大麻哈魚種群中也有相似的發現[2,16],這也與Morita等[6]的研究結果一致。綏芬河大麻哈魚雌、雄個體的L50分別為51.53和42.15 cm, 而王繼隆等[10]計算2010—2011年烏蘇里江雌、雄大麻哈魚L50分別為: 47.95和46.19 cm。在本研究中綏芬河雌性大麻哈魚L50大于烏蘇里江個體, 雄性L50則小于烏蘇里江個體。魚類性成熟的時間受遺傳因素和生態環境因子的影響, Morita等[6]研究發現大麻哈魚的生長狀況決定了其性成熟的時間。可見, 大麻哈魚不同地理種群、不同年代間的生長環境導致其生長速度、性成熟年齡的差異。
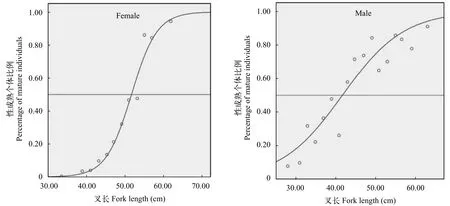
圖3 大麻哈魚雌雄性成熟個體在各體長組所占比例的邏輯斯蒂方程擬合Fig. 3 Logistic functions fitted to percent mature of female and male individuals of chum salmon
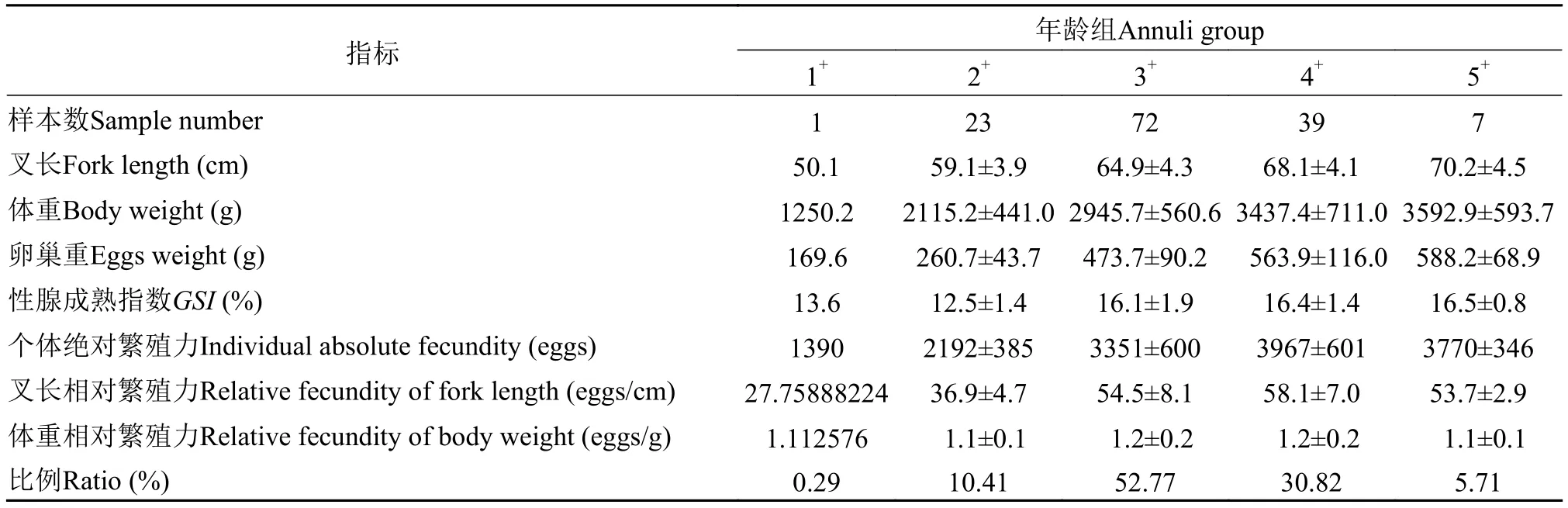
表3 大麻哈魚繁殖生物學指標Tab. 3 Reproductive biological indicators of chum salmon
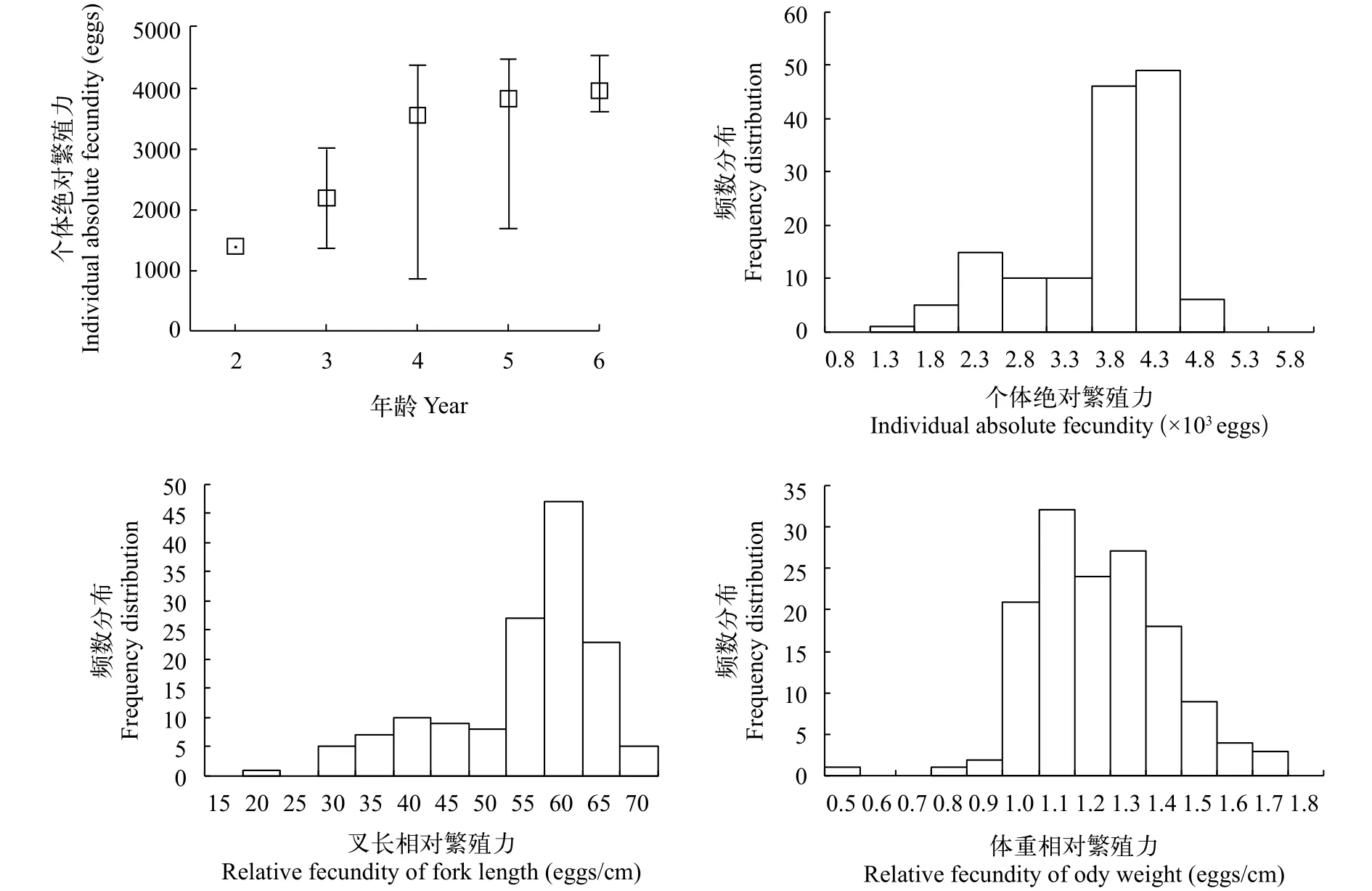
圖4 大麻哈魚個體繁殖力分布Fig. 4 Distribution of individual fecundity for chum salmon
3.2 綏芬河大麻哈魚的繁殖生物學特征
魚類繁殖力體現魚類群體間及種間的特性, 是魚類對周圍環境的適應性表現。2012—2017年綏芬河大麻哈魚平均懷卵量為3412粒, 與董崇智等[12]1986年調查結果相比, 平均懷卵量下降近20%。大麻哈魚絕對繁殖力(F)與年齡、叉長、體重都成正相關關系, 可見, 綏芬河大麻哈魚個體絕對繁殖力的下降主要是由個體低齡化、小型化所導致, 這種現象在大黃魚(Pseudosciaena crocea)、藍點馬鮫(Scomberomorus niphonius)等魚類研究中有相似報道[17,18]。和其他鮭科魚類相比(表4), 大麻哈魚的絕對繁殖力高于細鱗鮭[19]、馬蘇大麻哈魚[21,22]、駝背大麻哈魚[11]、亞東鮭[25]、花羔紅點鮭[26], 低于烏蘇里白鮭[23]、高白鮭[24]、哲羅鮭[20]。體重相對繁殖力是魚類長期自然選擇的結果, 體現了魚類的繁殖策略, 它是判定魚類繁殖力大小和魚類種屬和生態特異性的指標[27,28]。一般來說, 懷卵量高的魚類, 魚卵成活率低, 而懷卵量低的魚類往往成活率高, 魚類通過各自策略來維持物種的穩定[29], 大麻哈魚的FW和馬蘇大麻哈魚、哲羅鮭、細鱗鮭接近,可見這幾種魚類的繁殖策略相似, 體現了該類魚魚卵粒徑大, 懷卵量相對少, 魚卵成活率高的特點。但是和烏蘇里白鮭、高白鮭相比大麻哈魚FW低很多, 表明鮭科魚類間具有不同的繁殖策略, 體現了物種間繁殖策略的多樣性。
不同魚類間的繁殖力與生物學指標的擬合關系不同, 相關關系也存在差異。在本研究中大麻哈魚F與叉長、體重、性腺重成顯著正相關關系, 采用冪函數擬合了F與叉長、體重的關系。這與翹嘴鲌、黑斑刺蓋太陽魚和銀鲇等魚類[30—32]的研究結果相似。綏芬河大麻哈魚GSI與叉長、體重、F成顯著負相關關系, 與烏蘇里江大麻哈魚的研究結果一致[10]。FW與GSI、F和FL成顯著正相關關系, 這在翹嘴鲌、烏蘇里白鮭等魚類的研究中有類似的發現, 但也存在一定的差異[23,32]。叉長、體重、GSI是衡量大麻哈魚繁殖力的重要指標, 實際工作中可利用叉長、體重與繁殖力之間的關系來估算大麻哈魚的種群繁殖力。
綏芬河大麻哈魚群體中出現了低齡化、小型化、繁殖力下降等生物學特征的改變。尤其是低齡個體在群體中占有較高的比例, 導致大麻哈魚的種群結構發生變化。雖然多年的增殖放流工作對維持綏芬河種群數量的穩定起到了重要作用, 但是大麻哈魚生物學特征的變化也應引起足夠的重視。因此, 應長期對綏芬河大麻哈魚生物學特征進行監測, 為大麻哈魚種群資源保護與恢復以及群體多樣性保護工作提供科學依據。
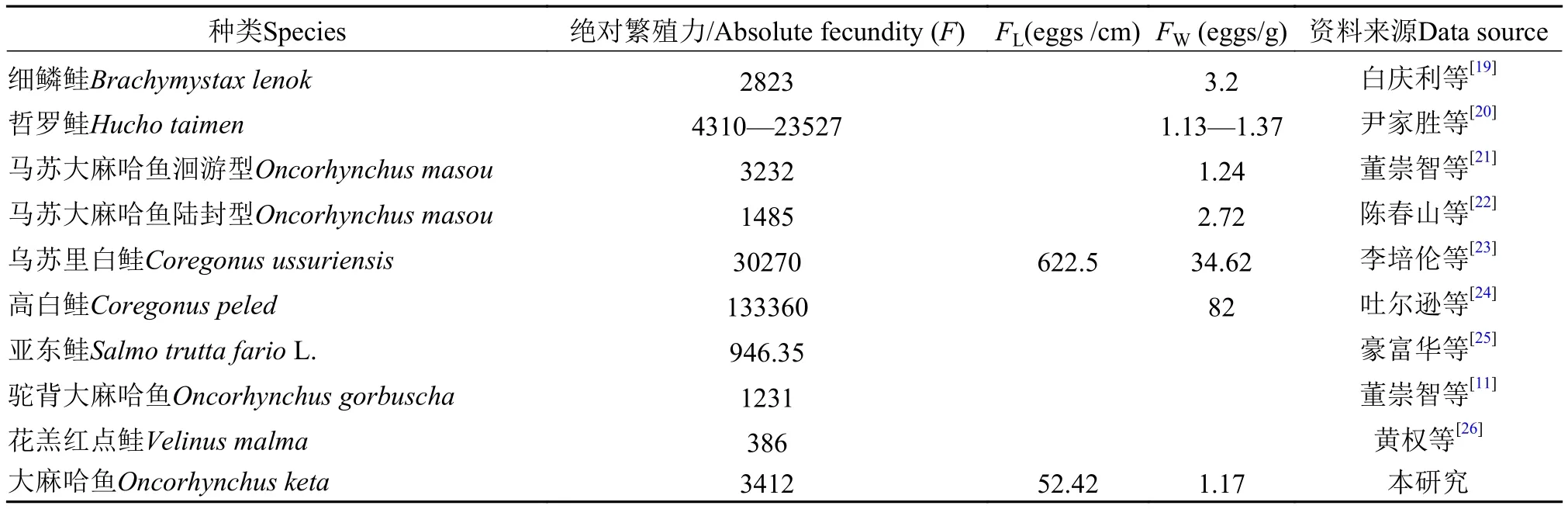
表4 大麻哈魚與其他幾種鮭科魚類繁殖力比較Tab. 4 Comparison of fecundity among chum salmon and other species of the family Salmonidae
猜你喜歡
現代畜牧科技(2021年4期)2021-07-21 06:13:00
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
數學物理學報(2017年2期)2017-06-05 09:12:30
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
