轉基因金葉銀中楊葉色及生長變異分析
2020-03-05 04:04:28李藝迪顧宸瑞冮慧欣劉桂豐
植物研究 2020年6期
關鍵詞:生長
李藝迪 顧宸瑞 冮慧欣 劉桂豐 陳 肅 姜 靜
(林木遺傳育種國家重點實驗室,東北林業大學,哈爾濱 150040)
GLK 轉錄因子又稱為Golden2-Like 和G2-Like,是Myb 類轉錄因子中的GARP 家族成員[1],其功能主要調控植物葉綠體發育、果實品質,同時也參與植物生物脅迫、植物衰老和激素信號轉導等[2~6]。研究證明,擬南芥(Arabidopsis thaliana)中的2 條GLK 功能冗余,雙突變體Atglk1、Atglk2葉色表現出淡綠色表型[7]。在番茄(Solanum lycopersicum)中,SlGLK1 和SlGLK2 的過量表達使番茄未成熟果實更加深綠色,同時促進果實均勻成熟、進而改善成熟果實的品質[3~4]。在C4 植物玉米(Zea mays L.)和高粱(Sorghum bicolor(L.)Moench)中,GLK1 和GLK2 分別在葉鞘(BS)和葉肉(M)細胞中表達[8]。然而,C4 植物白菜花(Cleome gynandra)中,CgGLK1 和CgGLK2 均在葉鞘和葉肉細胞中表達,并且在葉肉細胞中表達量更高[9]。水稻(Oryza sativa)的OsGLK1 參與調控葉綠體發育,并且Os-GLK1 在Chl 的生物合成中發揮重要作用[10]。對擬南芥的研究發現,AtGLK也參與病原微生物的防御作用,例如,該基因的過量表達AtGLK1OX株系對禾谷鐮刀菌(Fusarium graminearum)、葡萄孢菌(Botrytis cinerea)抗性顯著高于野生型擬南芥[11~13]。在林木中有關GLK 基因的研究鮮有報道,對白樺(Betula platyphylla Suk.)研究發現,其基因組中僅有1 個BpGLK1 成員,該基因的缺失突變及干擾表達轉基因白樺均表現葉片葉綠素含量降低,葉片呈現黃色[14~15]。
隨著城市的發展,人們對葉色艷麗的高大喬木需求越來越大,采用基因工程技術改變植物葉色是快速創制植物新品種的有效途徑,例如,過表達PdMYB118 的轉基因山新楊(P.davidiana×P.bolleana)在葉片中產生非常多的花色苷,并且在溫室和田間生長時將其顏色變成紅色[16]。過表達PtrMYB119 毛果楊(Populus trichocarpa)花青素水平顯著提高,葉色呈現紅色[17]。上述MYB 基因在彩色樹種選育中具有潛在的應用價值。除此之外,MdMYB10 過表達蘋果(Malus domestica)中,花青素水平升高,色素沉著,形成紅色果實[18]。轉基因煙草中SoCHS1 的過表達將花色由淺粉紅色變為粉紅色[19]。
銀中楊(Populus alba×P.berolinensis)樹皮灰綠色,樹冠呈圓錐形,樹姿優美,葉片較大,葉面深綠色,葉背面銀白色,密生絨毛,在城市園林綠化中廣泛應用[20]。筆者所在團隊根據白樺BpGLK1 研究思路,采用農桿菌介導的葉盤法獲得了PaGLK基因過表達及抑制表達銀中楊,其中獲得的3 個PaGLK 抑制表達銀中楊葉色也為金黃色,但是亮度及黃色程度不盡相同。為了解轉PaGLK 基因銀中楊葉色的變異規律,以及葉色黃化對轉基因銀中楊生長是否產生影響,試驗以PaGLK 過表達、PaGLK 抑制表達轉基因株系為材料,分析葉綠素相對含量與葉色的時序變異規律,探討葉綠素含量的降低對株高生長的影響。研究結果為后續轉基因銀中楊在城市園林綠化中推廣應用提供參考。
1 材料與方法
1.1 材料
轉PaGLK 基因銀中楊:PaGLK 過表達株系(G1、G2、G3);PaGLK 抑制表達株系(Y1、Y2、Y3)均由研究團隊保存。
1.2 方法
1.2.1 轉基因銀中楊分子檢測
分別提取PaGLK 過表達轉基因銀中楊(G1-G3)葉片DNA 為模板,以pCAMBIA1300-PaGLKGFP 質粒為陽性對照,以非轉基因銀中楊(WT)的總DNA 為陰性對照,同時設水為陰性對照,引物PaGLK-F,PaGLK-R(見表1),PCR 反應體系按相關文獻[21],反應結束后,吸取2μL 進行電泳檢測,觀察目的條帶的位置。
分別提取PaGLK 抑制表達轉基因銀中楊(Y1-Y3)葉片DNA 為模板,以pFGC5941-CIS-F 和pFGC5941-Cis-R,pFGC5941_Anti_F 和pFGC5941-Anti-R 作為引物(見表1)進行PCR 檢測。以pFGC5941-PaGLK 質粒為陽性對照,以非轉基因銀中楊(WT)的總DNA為陰性對照,同時設水為陰性對照,PCR 反應體系按相關文獻[21],反應結束后,吸取2 μL 進行電泳檢測,觀察目的條帶的位置。
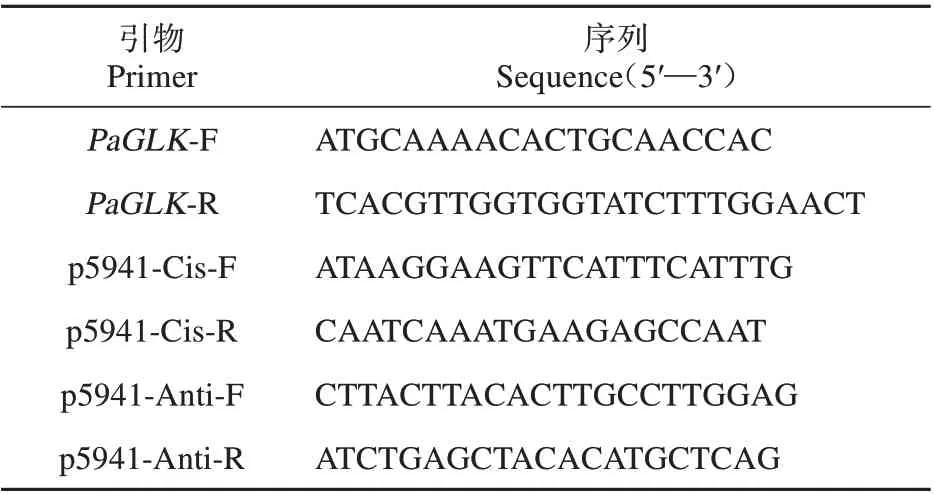
表1 引物序列Table 1 Primer sequences
qRT-PCR 檢測:分別提取參試株系總RNA,反轉錄為cDNA,將其稀釋10 倍后用于定量qRTPCR 的模板,以18S rRNA 為內參基因(見表2),進行對PaGLK 基因的qRT-PCR 分析,qRT-PCR 反應體系及擴增程序[15]。
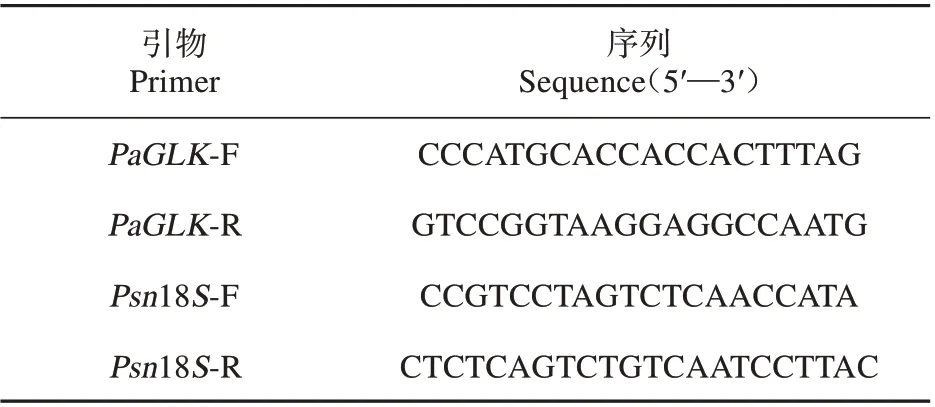
表2 qRT-PCR 引物序列Table 2 qRT-PCR primer sequence
1.2.2 轉基因銀中楊苗木擴繁及葉色調查
采用組培微繁技術分別擴繁野生型銀中楊(WT)及各轉基因株系,4月中旬移栽至育苗盤中,置于溫室中常規管理,5月中旬初選取高度一致的苗木,移栽至18 cm×13 cm 花盆中,每個株系共30株,分3次重復,7個株系,共計210盆苗木,放置在塑料大棚中進行常規水肥管理。
RHS 比色:將葉片近軸面與RHS 標準比色卡(英國皇家園藝學會,2015)進行對比,記錄葉片的顏色。
色差儀比色:于2019 年5 月15 日開始至9 月15 日結束,每隔15 d 分別對參試株系測定1 次,共計9次。采用分光色差儀(CR-400,Japan)測定第4葉的葉色參數,測量結果用CIELab 表色系統進行色度分析[22]。
1.2.3 葉片葉綠素相對含量測定
采用便攜式葉綠素測定儀(SPAD-502 PLUS,KONICA MINOLTA)測定轉基因及WT 株系功能葉的葉綠素相對含量,讀取SPAD 值,測定于5 月15 日開始,9 月15 日結束,每15 天測定1 次,每次每個株系測定10株樹葉片,3次重復。
1.2.4 生長性狀調查
于2019 年5 月中旬開始調查其生長等性狀,每15天測量1次,直至該年9月份整個植物生長周期測量結束。
數據利用MATLAB Compiler Runtime 8.3對苗高的生長規律進行Logistic方程擬合,Logistic曲線方程:

式中:y 為苗木苗高(cm);t 為苗木生長的時間,1 月1 日為1(d);a 為苗木開始生長時的苗高初始值(cm);b 為苗木停止生長時苗高值(cm);t0為苗木苗高日生長速度最快的日期(d);c 為曲線在苗木苗高日生長速度最快處的斜率(cm·d-1)。
在符合“S”型曲線的林木個體生長過程中,速生期是林木生長的關鍵階段,通過擬合方程得出速生期參數:
速生期的起始點t1;
速生期的結束點t2;
速生點t0;
速生期的持續時間RR=t2-t1;
速生期苗高的平均生長量GR=t2點的苗高-t1點的苗高;
速生期內苗高日生長量的平均值GD=GR/RR;
速生期內生長量占總生長量的比值RRA=GR/b-a。
2 結果與分析
2.1 轉基因株系的分子檢測
分別以銀中楊PaGLK 過表達株系的葉片總DNA 為模板,結果顯示:陽性質粒及3 個轉基因株系在1 200 bp 處擴增出單一譜帶,與PaGLK 序列堿基長度吻合(1 269 bp),而WT 株系無擴增譜帶(圖1)。
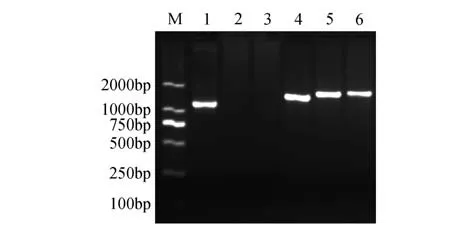
圖1 PaGLK過表達轉基因株系PCR擴增電泳圖譜M.DNA Maker DL2000;1. 陽性質粒;2. 陰性對照(水);3.WT株系;4~6.G1-G3株系Fig. 1 PaGLK overexpression transgenic lines PCR electrophoresis patternM.Marker DL2000;1.Positive plasmid;2.Negative control(ddH2O);3.WT line;4-6.G1-G3 line
分別以銀中楊PaGLK 抑制表達株系的葉片總DNA 為模板,結果顯示:3 個轉基因株系正向和反向均有擴增譜帶,且與預期的200 bp 長度吻合(圖2),實驗結果證明目標基因已經整合到銀中楊基因組中。
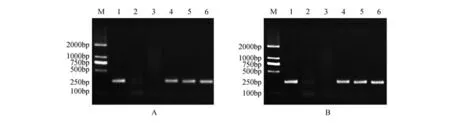
圖2 PaGLK抑制表達轉基因株系PCR擴增電泳圖譜A.PaGLK抑制表達轉基因株系正向PCR;B.PaGLK抑制表達轉基因株系反向PCR M.DNA Maker DL2000;1. 陽性質粒;2. 陰性對照(水);3.WT株系;4~10. 轉PaGLK基因抑制表達株系Y1~Y3Fig.2 PaGLK repression transgenic lines PCR electrophoresis patternA.PCR of forward fragment in PaGLK repression transgenic lines;B.PCR of reverse fragment in PaGLK repression transgenic lines M.DNA Maker DL2000;1.Positive plasmid;2.Negative contro(lddH2O);3.WT line;4-6.Y1-Y3 line
進而對獲得的轉基因銀中楊進行qRT-PCR 檢測(見圖3),結果顯示G1、G2、G3 過表達株系中PaGLK 基因表達量顯著高于WT 株系,分別是WT株系的6.7、3.7、3.8倍,而Y1、Y2、Y3抑制表達株系中PaGLK 基因則顯著下調表達,較WT 株系低65.3%、68.7%、73.70%。進一步證明PaGLK 基因已經整合到銀中楊基因組中。
將銀中楊PaGLK 與白樺BpGLK1 序列利用BioEdit軟件進行多序列比較,比較結果(見圖4)。
2.2 轉基因銀中楊的葉色變異
CIELab 表色系統中L*值是衡量葉色明暗程度的指標,該值越大則葉片亮度越高,b*值表示葉片的藍黃屬性,b*值由大變小,則表示藍色減退黃色增加[22]。

圖3 轉基因株系相對定量分析注:不同小寫字母表示P<0.05顯著水平Fig.3 RelativequantitativeanalysisofthetransgeniclinestNote:Different lowercase letter represents the significance difference P<0.05
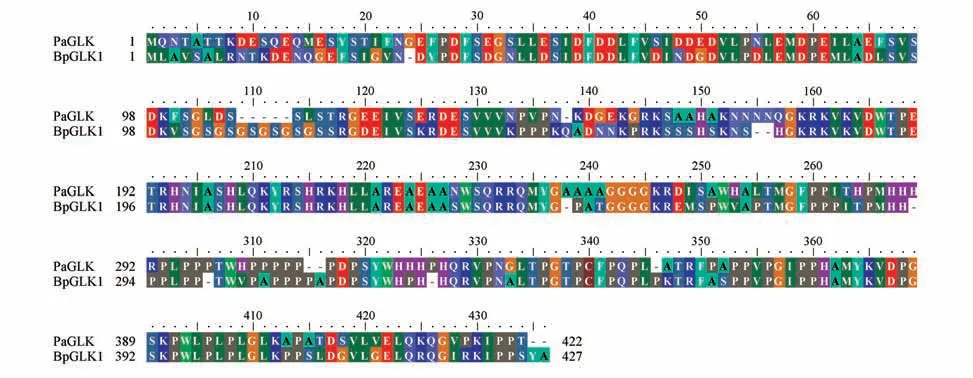
圖4 銀中楊PaGLK與白樺BpGLK1序列的多序列比對Fig.4 Multiple sequence alignment of PaGLK and BpGLK1 in birch
對不同發育期各參試株系的葉色參數L*值分析,結果表明,PaGLK 過表達株系中的G1 株系在整個發育階段都顯著低于WT 株系,而G2、G3 株系的L*值僅在8 月1 日、8 月15 日顯著低于WT 株系,其他發育階段與WT 株系差異不顯著(見圖5)。參試株系的3 個抑制表達株系從6 月初開始到9 月中旬其L*值持續高于WT 株系,并達到顯著水平(P<0.05)。說明,銀中楊PaGLK 基因的低量表達使葉片的亮度其他株系。各株系的葉色參數L*值時序變化均在7月15日該值最大,此時3個抑制表達株系的L*值均顯著高于WT及過表達株系,其均值較WT 株系均值高15.4%、較3 個過表達株系均值高19.9%。
不同發育期各參試株系b*值測定,結果表明,G1過表達株系的b*值在整個發育階段都顯著低于WT株系,而G2、G3株系在5月15日到7月15日之間其葉片b*值與WT 株系差異不顯著,從8 月開始直到調查結束,b*值一直低于WT 株系,并達到顯著水平(P<0.05)(見圖5)。抑制表達株系從7 月中旬到9 月中旬這期間的b*值一直顯著高于WT株系,并達到顯著水平(P<0.05)。在6 月15 日的b*值低于其他時期,并且抑制表達株系的b*值與對照株系差異不顯著。隨著參試株系的生長7 月15日到9月15日之間,3株抑制表達株系的b*值均顯著高于WT株系。
RHS 比色卡測定結果顯示(見表3),WT 株系始終呈現中等黃綠色,抑制表達株系的葉色始終呈現深黃綠色(RHS2015 144B),而過表達株系的葉色在7月中旬為中等橄欖綠色和深黃綠色,隨后漸漸的變為淺橄欖綠。
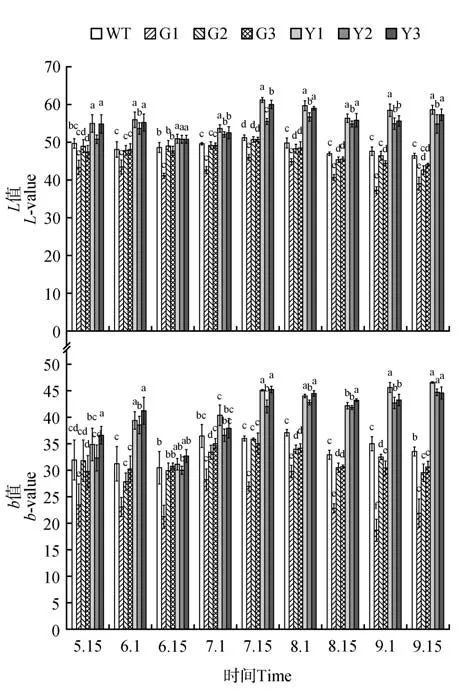
圖5 參試株系葉片L*值、b*值時序變化注:a、b、c、d為同一時間點參試株系L*值、b*值的多重比較Fig.5 Temporary variation of test lines about value b and value LNote:a,b,c and d in figures indicate the multiple comparisons between test lines about value b*and value L*in the same time
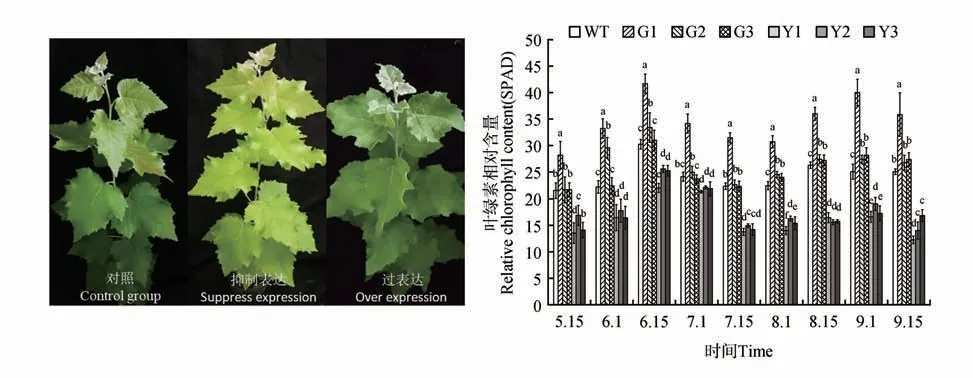
圖6 參試株系的葉色觀察和葉綠素相對含量(SPAD)時序變異注:a、b、c、d為同一時間點參試株系SPAD值的多重比較Fig.6 Observation of leaf color and temporal variation of relative chlorophyll conten(tSPAD)in the test linesNote:a,b,c and d in figures indicate the multiple comparisons between test lines about SPAD value in the same time
2.3 轉基因銀中楊葉綠素含量分析
轉基因銀中楊葉的葉色不同于WT 株系(圖6),對不同發育期的各參試株系進行SPAD 的測定,結果表明(見圖6),參試株系在整個生長過程中,過表達株系G1 的SPAD 始終顯著高于對照株系,G2、G3 在8 月1 日到9 月1 日始終高于WT 株系,并達到顯著水平(P<0.05)抑制表達株系的SPAD 在6 月1 日到9 月15 日 都顯著低于WT 株系和過表達株系,其中在9 月15 日的SPAD 是最低的,其均值低于WT 株系的42.7%,低于過表達株系SPAD均值的52%。
2.4 轉基因銀中楊的生長特性分析
2.4.1 轉基因銀中楊苗高生長模型的建立與擬合
分別對轉基因株系苗高生長過程進行擬合(見圖7)轉基因及WT 株系苗高生長過程為“S”型曲線,用四參數Logistic 方程式分別對轉基因及對照銀中楊苗高的平均值進行擬合,并繪制出“S”形生長曲線(見圖7)。各生長模型方程的擬合系數均高于0.99,達到顯著水平,說明用四參數Logistic曲線方程對轉基因及WT 株系生長節律進行擬合是準確可靠的,可用于轉基因及WT株系高生長分析與預測。
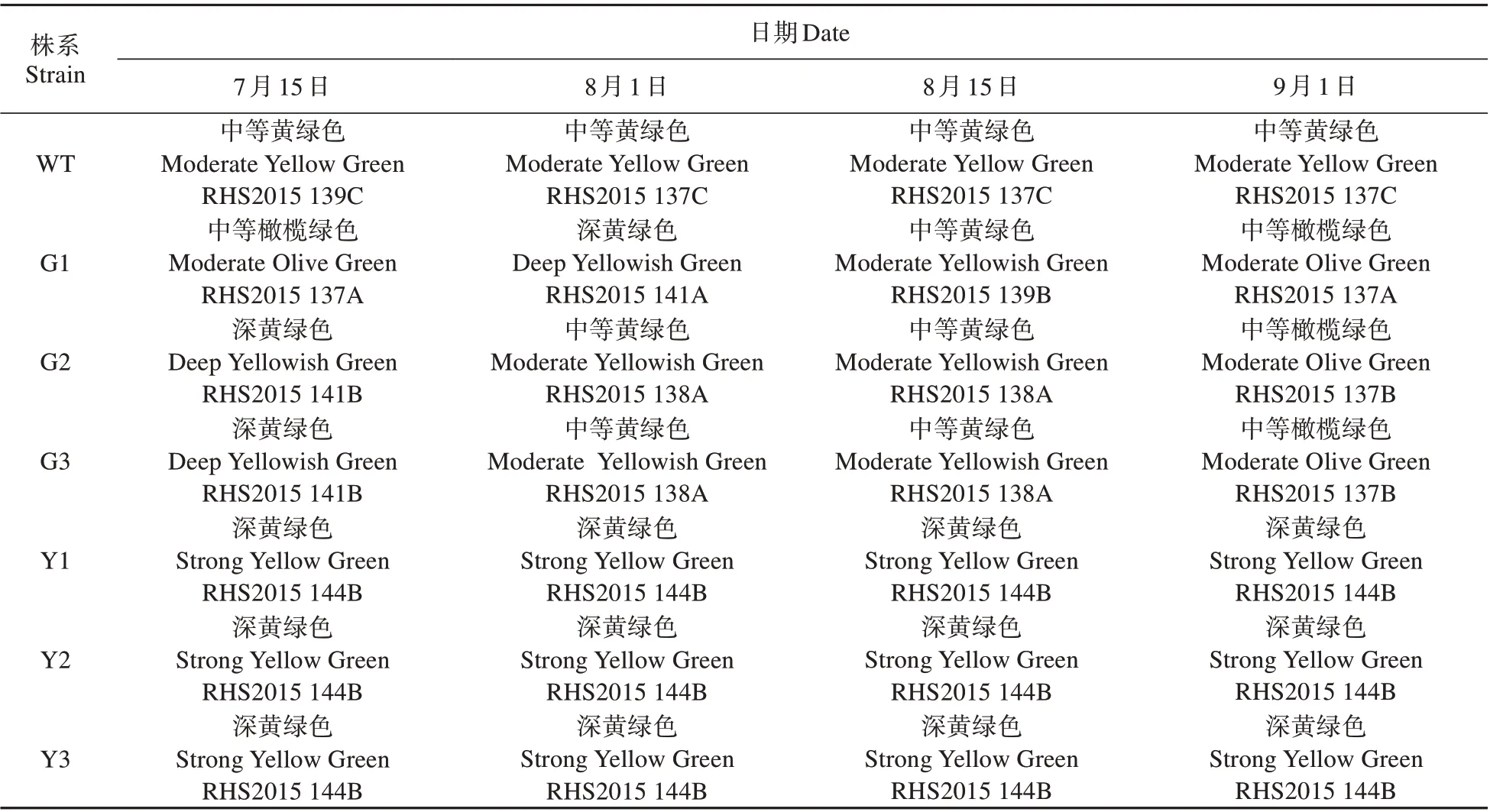
表3 參試株系的葉色時序變化Table3 Time series of leaf color of the tested plants
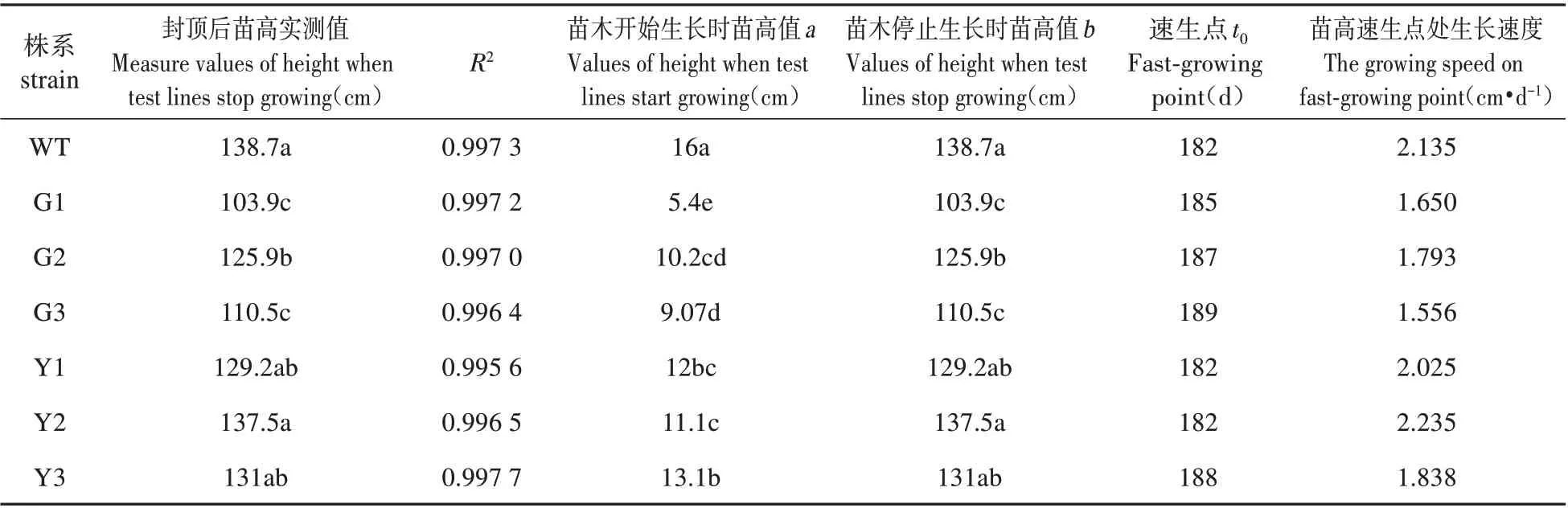
表4 轉基因銀中楊苗高的Logistic模型Table 4 Logistic model of poplar seedling height in transgenic silver

圖7 轉基因株系樹高邏輯斯蒂擬合曲線Fig.7 Logistic fit curve of tree height of transgenic lines
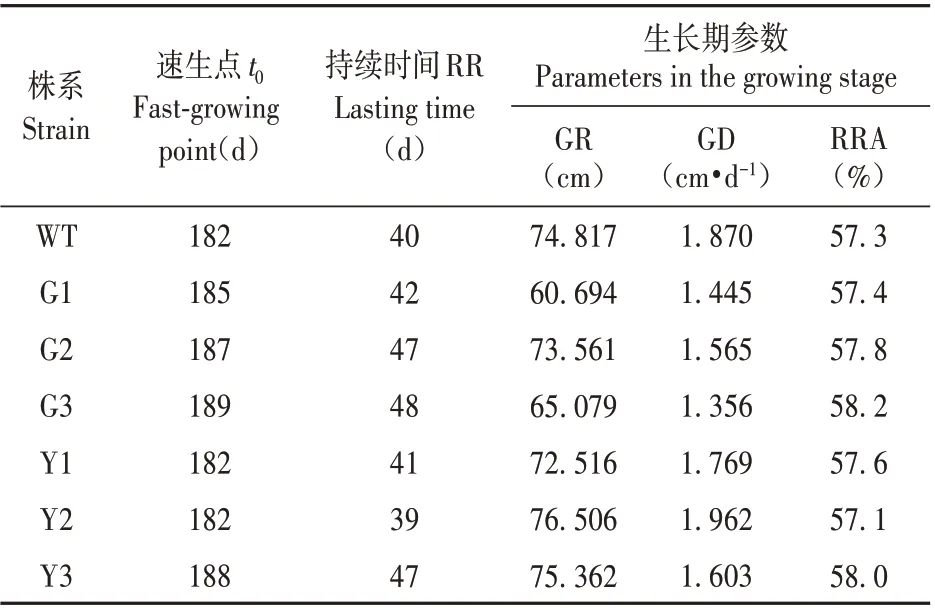
表5 轉基因株系速生期生長參數比較Table 5 Comparison of growth parameters of transgenic strains in the fast growing period
由表4 可知,過表達株系與WT 株系的苗高生長(即停止生長后的實測值)差異顯著(P<0.05),過表達株系當年高生長顯著低于WT株系,其苗高均值低于WT 株系的18.22%,其中G3 苗高速生點最高,為1.793 cm·d-1,低于WT 株系。抑制表達株系停止生長后的苗高與WT株系差異不顯著,其中Y2的苗高速生點最高,為2.235 cm·d-1。
2.4.2 轉基因銀中楊速生期生長參數變異
轉基因銀中楊苗高從5 月中旬開始生長,9 月中旬封頂,生長期約為120 d。轉基因株系與WT株系從開始生長、到進入速生期以及速生期結束的時間基本一致。由表5可知,過表達株系在速生期內苗高的平均生長量(GR)低于對照株系,其速生期內苗高日生長量均值(GD)也低于WT 株系,其均值為WT 株系的22.19%。然而抑制表達株系中的Y2 速生期內苗高日生長量均值(GD)高于WT 株系,另外兩個株系速生期內苗高日生長量(GD)沒有高于WT 株系,但是速生期的持續時間長于WT株系,因此導致最終停止生長時的高生長與WT差異不顯著。
3 討論
銀中楊是銀白楊與中東楊雜交選育出的三倍體雄性無性系,該無性系不僅具有樹干通直,樹皮灰綠、樹姿優美的特點,還具有雄性敗育、無散粉的特點,在城市園林綠化中深受人們青睞[20]。以其為受體獲得的轉基因金葉銀中楊,可以免去外源基因通過花粉向非轉基因品種或野生近緣物種漂移給人們帶來的后顧之憂,可以說通過基因工程技術創制的金葉銀中楊在城市園林綠化中發展前景廣闊。
Golden2-like(GLK)基因家族的成員可以協調光合作用的表達并控制不同植物物種中葉綠體的發育[7,23~26]。根據報道,擬南芥和苔蘚中,GLK1 和GLK2 同時缺失的突變體的葉片表現為葉綠素含量降低,葉片變黃。番茄中SlGLK1 的缺失也能引起葉片顏色變淺[3,7]。研究證明,白樺BpGLK1 抑制表達株系的葉綠素含量明顯降低,葉片呈現深黃綠色,過表達株系的葉綠素含量比對照高[15],植物葉色變異也往往與葉綠體中的色素含量的變化有關[27~28],本試驗中轉PaGLK 銀中楊苗期試驗顯示,過表達株系葉色為綠色,9 月時葉色為中等橄欖綠色,株高生長較WT 緩慢;而3 個抑制表達株系的葉色參數代表葉片亮度的L*值均顯著高于WT 株系,尤其是Y1 和Y3 株系葉片亮度更高,代表葉片黃色的b*值也顯著高于WT 株系,在整個生長季,Y1和Y3株系一直保持鮮艷的黃色。從葉片亮度及色彩方面分析,3 個抑制表達株系中Y1 和Y3 株系更鮮艷,應用推廣潛力更大,后續還需繼續追蹤調查。
光合作用可以為植物提供能量,從而促進植物生長。葉綠體是植物細胞內最重要、最普遍的質體,它是進行光合作用的重要場所。所以,葉綠體的發育與植物生長發育有密切的關系。在水稻低葉綠素突變體中發現,雖然葉綠素含量降低,但是葉綠體的發育沒有受到影響,在高光照條件下具有較高的光合速率[29]。適當降低葉綠素含量,不僅有利于減少光抑制,而且還有利于氮素在光合系統內的分配,從而提高光合氮素利用效率[30~31]。本試驗中,轉PaGLK 銀中楊苗期試驗顯示,PaGLK 抑制表達并未影響銀中楊苗期株高生長,移栽當年停止生長時抑制表達株高均值為132.58 cm,與WT 株系差異不顯著,但是,PaGLK過表達株系的株高卻顯著低于WT株系,可能葉綠素含量顯著增高,葉片吸收了過量的光能,導致光抑制現象所致。總之,光合作用是一系列復雜的代謝反應總和,僅維持葉片高葉綠素含量并不是提高有效光合速率的必要條件[32]。研究表明,在擬南芥和白樺的研究中,GLK 主要通過調控捕光天線蛋白、光系統復合體蛋白以及葉綠素合成等相關基因的表達,影響植物的光合色素的合成及葉綠體的發育[14~15,23],GLK 基因的缺失及抑制表達均可使植物葉片褪綠黃化,并且該性狀在大田中穩定表現[14~15,22]。因此,在其他植物中GLK 基因可以作為黃色彩葉樹育種的候選基因。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
