漓江大型底棲動物功能攝食類群時空分布及生態效應
2020-03-13 03:15:48朱晨曦莫康樂林育青陳求穩
生態學報 2020年1期
關鍵詞:功能
朱晨曦,莫康樂,唐 磊,吳 焱,李 婷,林育青,陳求穩
南京水利科學研究院, 南京 210029
河流生態系統由生物和生境兩部分組成,兩者一直處于動態變化的過程中。其中,生物是河流的生命系統,生境是河流生物的生命支持系統[1]。河流大型底棲動物是河流生態系統食物鏈結構中的重要環節,對河流生態系統的物質循環、能量流動有著積極作用[2]。同時,由于大型底棲動物的活動能力弱、活動范圍小,對于外來污染具有較小的規避能力,并且對外界脅迫較為敏感,因此能有效的指示河流生態系統的健康,其種類組成及時空分布更能體現生境變化對于生物的影響[3-4]。
大型底棲動物依據取食的不同劃分為5個功能攝食類群(Functional feeding groups, FFGs):捕食者、撕食者、刮食者、收集者和濾食者[5- 8]。大型底棲動物功能攝食類群與傳統大型底棲動物形態分類方法不同,它主要依據底棲動物的食物資源類型及在獲取食物過程中形態學的適應機制,來反映生境變化對大型底棲動物群落的影響,揭示出大型底棲動物的群落結構特征和生境適應性特征[9]。大型底棲動物功能攝食類群能較好的反映人類活動對河流生態系統的影響及河流生態系統的受損情況,因此近年來受到了研究者們的廣泛關注[9- 11]。
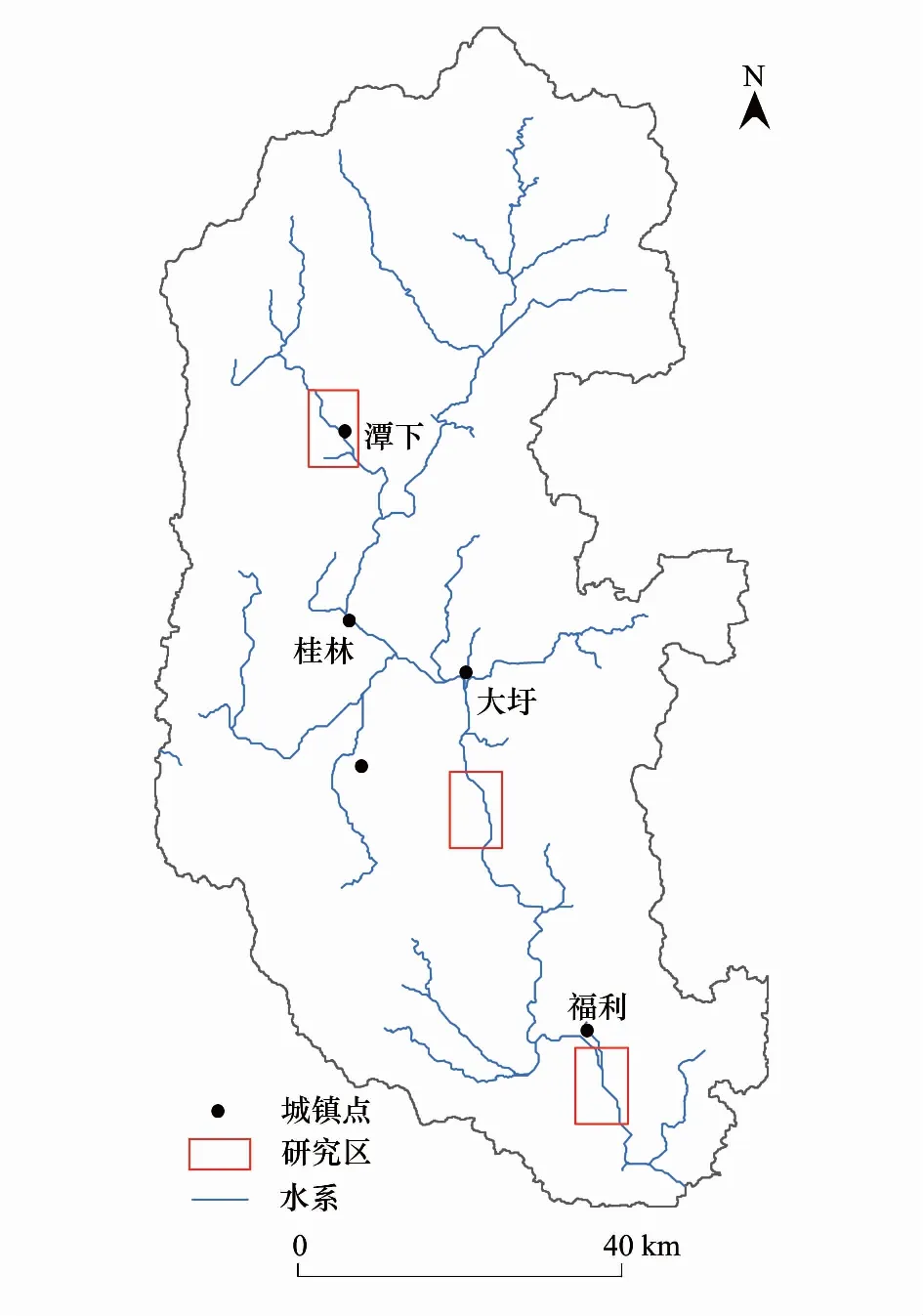
圖1 漓江研究區域分布Fig.1 The study reas in Lijiang River
漓江作為中國唯一一條入選的全球最美河流,是桂林山水的靈魂,保護漓江河流生態系統健康在維持漓江山水自然風景方面有著重要的意義[12]。以往應用大型底棲動物對漓江進行評價的研究,大多基于傳統的形態分類方法,考慮其與水環境、底質類型、沉水植物之間的關系[12-14],鮮有從功能攝食類群角度去評價漓江河流生態系統。本文以漓江為對象,研究大型底棲動物功能攝食類群的時空分布特征,并分析其與環境因子的相關性,采用基于功能攝食類群的生態參數評價漓江河流生態系統健康狀態,為漓江生態環境保護提供依據。
1 材料與方法
1.1 區域概況及采樣區設置
漓江屬于珠江流域西江水系,是支流桂江上游河段的通稱,位于中國的南部(110°18′—111°18′E,23°23′—25°59′N),發源于桂江源頭越城嶺貓兒山。漓江上游為從貓兒山到桂林河段、中游為從桂林到陽朔河段、下游為從陽朔到平樂河段,其中桂林到陽朔河段為著名的桂林漓江風景區。漓江流域總面積為12285 km2,主河道長214 km,主要包括甘棠江、黃柏江、川江、小溶江等幾條支流。
本研究分別在漓江上、中、下游選取典型江段進行研究(圖1)。其中,上游甘棠江研究江段長7 km,底質以細沙和淤泥為主;中游大圩研究江段長6 km,底質以卵石為主;下游福利研究江段長7 km,兩岸植被稀少,主要為人工栽植的鳳尾竹,底質以礫石為主。本研究在不同季節對長約20 km的研究江段進行大型底棲動物的采樣調查,調查時段分別為2009年秋季(9月)、2010年春季(4月)、2012年冬季(1月)和夏季(8月)。4次調查的樣本數分別為150個(上游54個,中游48個,下游48個)、150個(上游54個,中游41個,下游55個)、209個(上游55個,中游78個,下游76個)和165個(上游26個,中游75個,下游64個),并同步對部分水體理化指標及河流生境狀況進行了測定和記錄。
1.2 大型底棲無脊椎動物的采集與鑒定
本研究大型底棲動物的采集一共使用了兩種采樣工具,包括直徑30 cm的40目D型網和面積為1/20 m2的Peterson改良式采泥器。在現場采集時,于每個采樣點斷面的兩岸可涉水區域使用直徑30 cm的40目D型網采集底棲動物樣品共6個樣方,采集面積1.8 m2;斷面中間不可涉水區域使用面積為1/20 m2的Peterson改良式采泥器采集底棲動物樣品共4個樣方,采集面積0.2 m2。將每個斷面采集的10個樣方混合成一個樣品,采集面積共計2 m2。在現場用40目分樣篩篩選樣品,挑出大型底棲無脊椎動物后放入100 mL標本瓶中,然后用10%的甲醛溶液保存并帶回實驗室進行種類鑒定、個體計數、稱重。在鑒定時,所有樣品盡量鑒定到最低分類單元[15-17]。
1.3 水體理化指標測定
在采集現場使用便攜式多參數水質監測分析儀(YSI6600,美國金泉儀器)測定水溫(Temp)、pH、溶解氧(DO)、電導率(EC)、濁度(Tur)、鹽度(Salt),并在每個樣點采集2瓶550 mL水樣,其中一瓶現場加硫酸調整pH值至小于2,低溫保存運回實驗室后分別采用GB3838—2002[18]中規定的鉬酸銨分光光度法、堿性過硫酸鉀消解紫外分光光度法和高錳酸鹽法測定總磷(TP)、總氮(TN)、化學需氧量(COD)。
1.4 大型底棲無脊椎動物功能攝食類群及生態系統屬性劃分
根據河流大型底棲動物的食性類型,參照Cummins等、Barbour等和Bode等的劃分規則[5-8],將漓江大型底棲動物分為以下5類功能群(FFGs):(1)收集者(GC),主要以河底的各種有機物顆粒為食;(2)濾食者(FC),主要以水流中的細有機顆粒物為食(0.45 mm<粒徑<1 mm);(3)捕食者(PR),主要以捕食其他水生動物為食;(4)撕食者(SH),主要以各種凋落物和粗有機質顆粒為食(粒徑>1 mm);(5)刮食者(SC),主要以各種營固著生活的生物類群為食。
參考Yoshimura等及王博涵等的研究[9,19],本文根據大型底棲動物功能攝食類群的參數,從物質循環、物質縱向輸送能力、沿岸物質輸入等方面對漓江生態系統進行評價(表1)。

表1 大型底棲動物功能攝食類群所表征的生態系統屬性
1.5 數據處理與分析
采用物種優勢度指數(Y)來表示大型底棲動物各功能攝食類群中某一物種在其中所占優勢的程度,公式如下:
Y=(ni/N)fi
式中,N為各功能攝食類群大型底棲動物的總密度,ni為第i種的密度,fi為第i種出現的頻率。優勢度指數Y>0.02的種類確定為本次調查各功能攝食類群的優勢種[20]。
基礎數據統計均在Excel軟件中完成,前向篩選和冗余分析(RDA)在R軟件中完成(version 3.3.2,R development Core Team, http://www.r-project.org/)。
2 結果
2.1 大型底棲無脊椎動物物種組成及功能攝食類群組成
本研究4次采樣共采集到64個大型底棲無脊椎動物分類單元,隸屬于3門8綱16目38科。其中昆蟲綱28種、腹足綱25種、甲殼綱3種、瓣鰓綱3種、蛭綱2種、多毛綱1種、寡毛綱1種、蛛形綱1種。對漓江大型底棲無脊椎動物功能攝食類群進行分析,結果顯示,刮食者在漓江占顯著優勢,相對豐度為73.26%;其次為濾食者和收集者,相對豐度分別為15.12%和8.85%;捕食者和撕食者相對豐度較小,分別為2.72%和0.06%。由表2可知收集者、濾食者、捕食者、撕食者、刮食者的優勢物種,濾食者中的河蜆優勢度最大為0.367,除撕食者外其他功能攝食類群均存在兩種優勢物種。
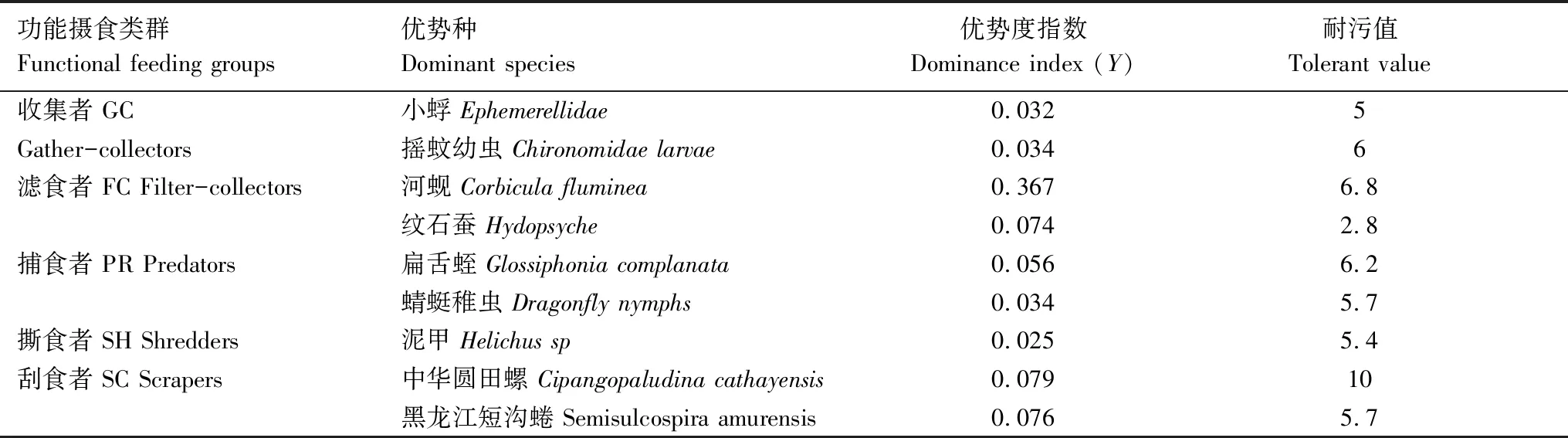
表2 漓江大型底棲無脊椎動物各功能攝食類群優勢種
2.2 大型底棲無脊椎動物功能攝食類群空間分布特征
2.2.1相對豐度空間分布特征
漓江大型底棲無脊椎動物功能攝食類群的相對豐度空間分布見圖2。優勢類群刮食者的最大相對豐度出現在夏季中游江段,為93.48%,最小出現在夏季上游江段,為25.27%;捕食者的最大相對豐度出現在冬季上游江段,為8.88%,最小出現在夏季中游江段,為0.75%;濾食者的最大相對豐度出現在夏季上游江段,為55.05%,最小出現在冬季中游江段,為3.79%;收集者的最大相對豐度出現在春季上游江段,為29.36%,最小出現在秋季下游江段,為0.62%;撕食者相對豐度在各江段都很小,只在夏季上游江段和秋季上游江段相對較大,分別為0.53%、0.42%。
2.2.2密度的空間分布特征
漓江大型底棲無脊椎動物功能攝食類群的密度空間分布如圖3所示。優勢類群刮食者的最大密度出現在夏季中游江段,為2354個/m2,最小出現在夏季上游江段,為73個/m2;捕食者的最大密度出現在春季下游江段,為84個/m2,最小出現在秋季上游江段,為4個/m2;濾食者的最大密度出現在秋季中游江段,為436個/m2,最小出現在冬季中游江段,為49個/m2;收集者的最大密度出現在春季中游江段,為208個/m2,最小出現在秋季下游江段,為5個/m2;撕食者密度在各江段都很小,只在春季、夏季、秋季的上游江段以及秋季的中游江段相對較大,分別為1、2、2、2個/m2。
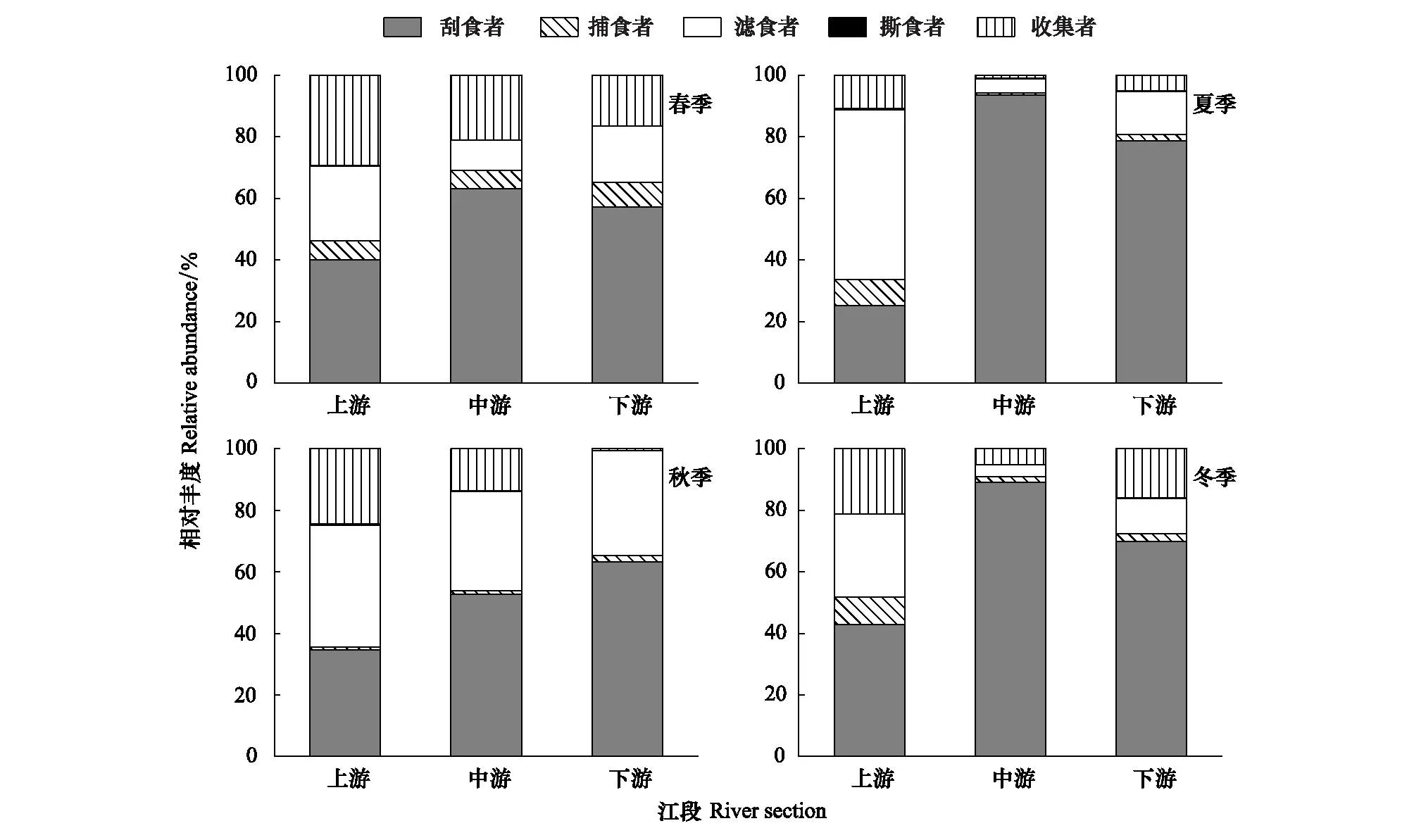
圖2 各季節漓江上中下游大型底棲動物功能攝食類群相對豐度Fig.2 The relative abundance of macroinvertebrate FFGs in Lijiang River among different river section in different seasonsFFGs: 功能攝食類群Functional feeding groups; SC:刮食者 Scrapers;PR:捕食者 Predators;FC:濾食者 Filter-collectors;SH:撕食者 Shredders;GC:收集者 Gather-collectors
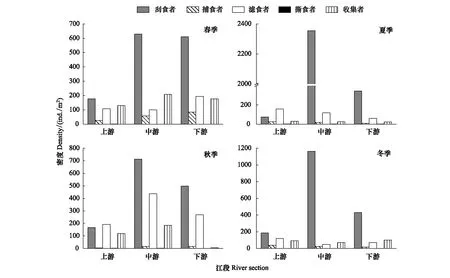
圖3 各季節漓江上中下游大型底棲動物功能攝食類群密度Fig.3 The density of macroinvertebrate FFGs in Lijiang River among different river section in different seasons
2.3 大型底棲無脊椎物功能攝食類群季節變化特征
2.3.1相對豐度季節變化特征
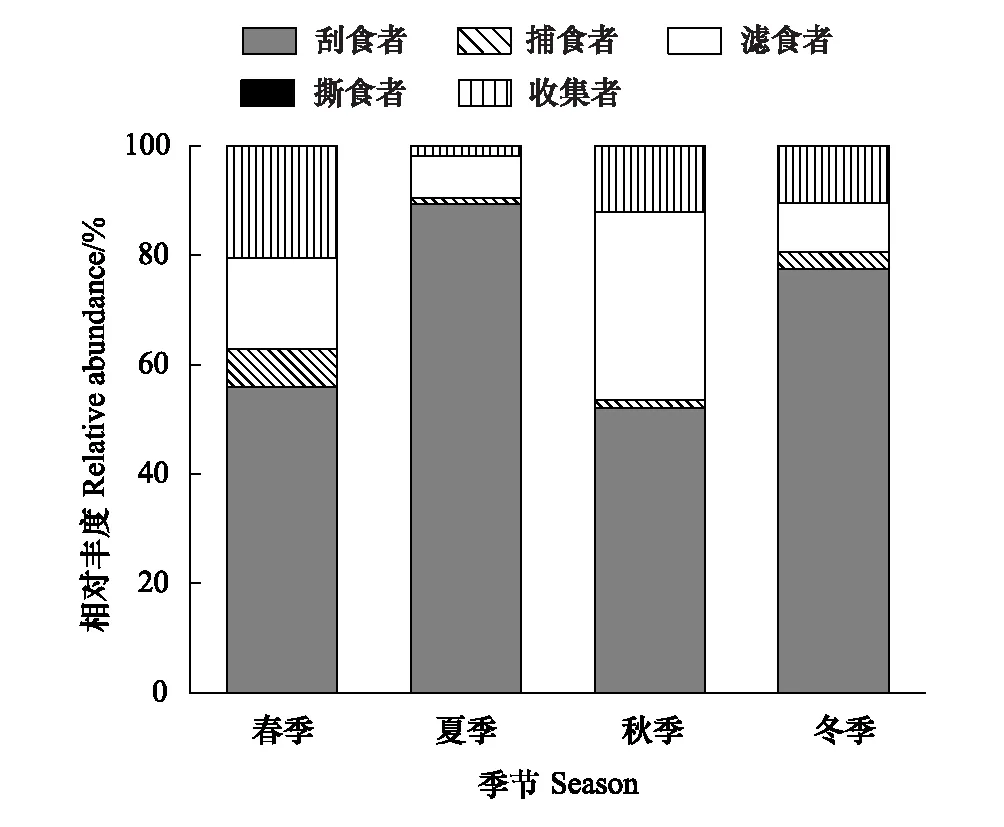
圖4 漓江大型底棲動物各功能攝食類群相對豐度的季節動態 Fig.4 The relative abundance of macroinvertebrate FFGs in different seasons in Lijiang River
圖4顯示了功能攝食類群相對豐度的季節變化。各季節刮食者相對豐度明顯高于其他功能攝食類群,一直處于優勢地位,最大值出現在夏季,為89.37%,最小值出現在秋季,為52.12%;撕食者相對豐度明顯低于其他功能攝食類群,一直處于劣勢地位,最大值出現在秋季,為0.17%,最小值出現在冬季,為0.01%;濾食者相對豐度最大值出現在秋季,為34.31%,最小值出現在夏季,為7.54%;收集者相對豐度最大值出現在春季,為20.46%,最小值出現在夏季,為1.86%;捕食者相對豐度最大值出現在春季,為6.86%,最小值出現在夏季,為1.16%。
2.3.2密度季節變化特征
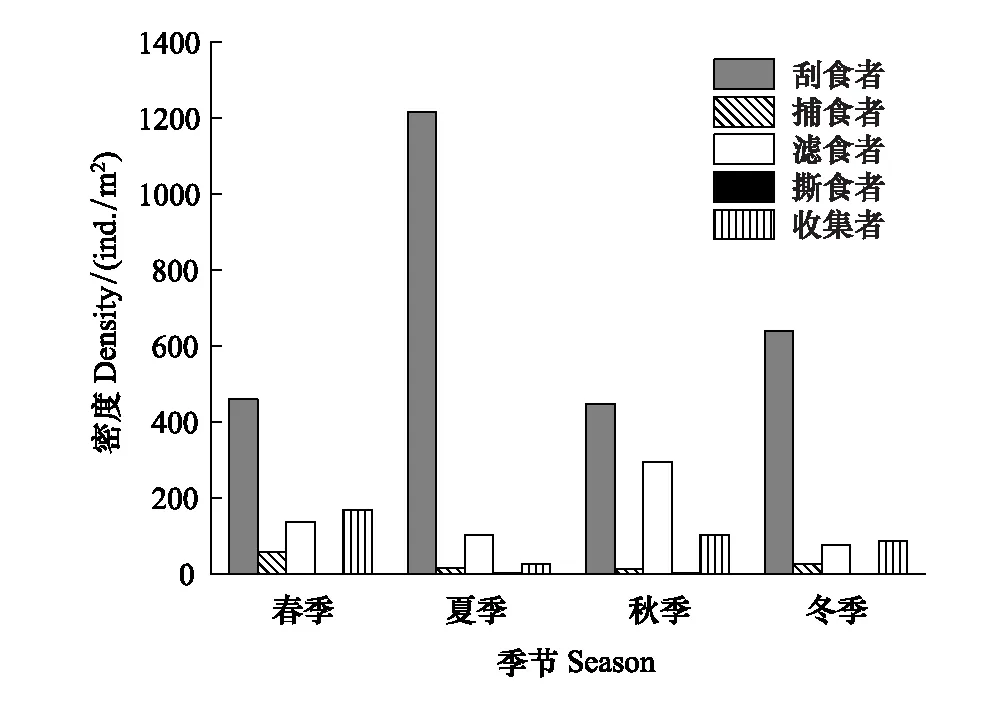
圖5 漓江大型底棲動物功能攝食類群密度的季節動態 Fig.5 The density of macroinvertebrate FFGs in different seasons in Lijiang River
功能攝食類群密度的季節變化見圖5。各季節刮食者密度明顯高于其他功能攝食類群,優勢地位明顯,最大值出現在夏季,為1215個/m2,最小值出現在秋季,為447個/m2;撕食者密度明顯低于其他功能攝食類群,劣勢地位明顯,最大值出現在秋季,為2個/m2,最小值出現在冬季,為0個/m2;濾食者密度最大值出現在秋季,為294個/m2,最小值出現在冬季,為75個/m2;收集者密度最大值出現在春季,為168個/m2,最小值出現在夏季,為25個/m2;捕食者密度最大值出現在春季,為56個/m2,最小值出現在秋季,為12個/m2。
2.4 基于大型底棲無脊椎動物功能攝食類群參數的漓江生態系統評價
基于功能攝食類群的各項參數進行時空差異性分析表明(表3):在物質循環方面,底棲動物初級生產力(F1)在漓江中游大于其余區域,夏季明顯大于其他季節;自養/異養(F2)在中游大于其余區域,夏季明顯大于其他季節;底棲動物分解能力(F3)在中游大于其余區域、夏季小于其他季節;底棲動物次級生產力(F4)從上游到下游逐漸增高。在物質縱向輸送能力方面,縱向輸送能力(F5)和相對縱向輸送能力(F6)中下游較高、夏季明顯較大。在沿岸物質輸入方面,沿岸物質輸入量(F7)和相對物質輸入量(F8)均為上游高于中下游,秋季高于其他季節。在其他方面,捕食者的下行效應(F9)在河流空間分布上差異性不大、四季變化也不大,但參數水平普遍較低;生境穩定性(F10)表現為從河流上游到下游逐漸增強。
綜合上述分析,對漓江生態系統進行評價。結果表明:漓江生態系統在空間上從上游到下游大型無脊椎底棲動物的生物量逐漸增加且生境的穩定性越來越強;在時間上表現為夏季漓江生態系統各項參數水平均較高,生境穩定性也最強。但從參數F9可以看出漓江生態系統內底棲動物的高營養級對低營養級控制力不強[21]。
2.5 大型底棲無脊椎動物功能攝食類群與環境因子的相關性分析
為了探究采樣期間不同環境因子對漓江大型底棲動物功能攝食類群的影響,對功能攝食類群與環境因子DO、pH、EC、Temp、Tur、Salt、TN、TP和COD進行冗余分析(RDA)(圖6),通過前向選擇(forward selection)排除掉貢獻較小的環境因子(P>0.05)。分析結果顯示,漓江四季對大型底棲動物功能攝食類群影響較大的環境因子有所不同,春季COD(P=0.001)和DO(P=0.005)兩個環境因子與漓江大型底棲動物功能攝食類群相關性較強;夏季EC(P=0.001)、pH(P=0.002)和DO(P=0.002)等3個環境因子與功能攝食類群相關性較強;秋季EC(P=0.001)和TN(P=0.024)與功能攝食類群相關性較強;冬季Temp(P=0.001)和Tur(P=0.001)與功能攝食類群相關性較強。
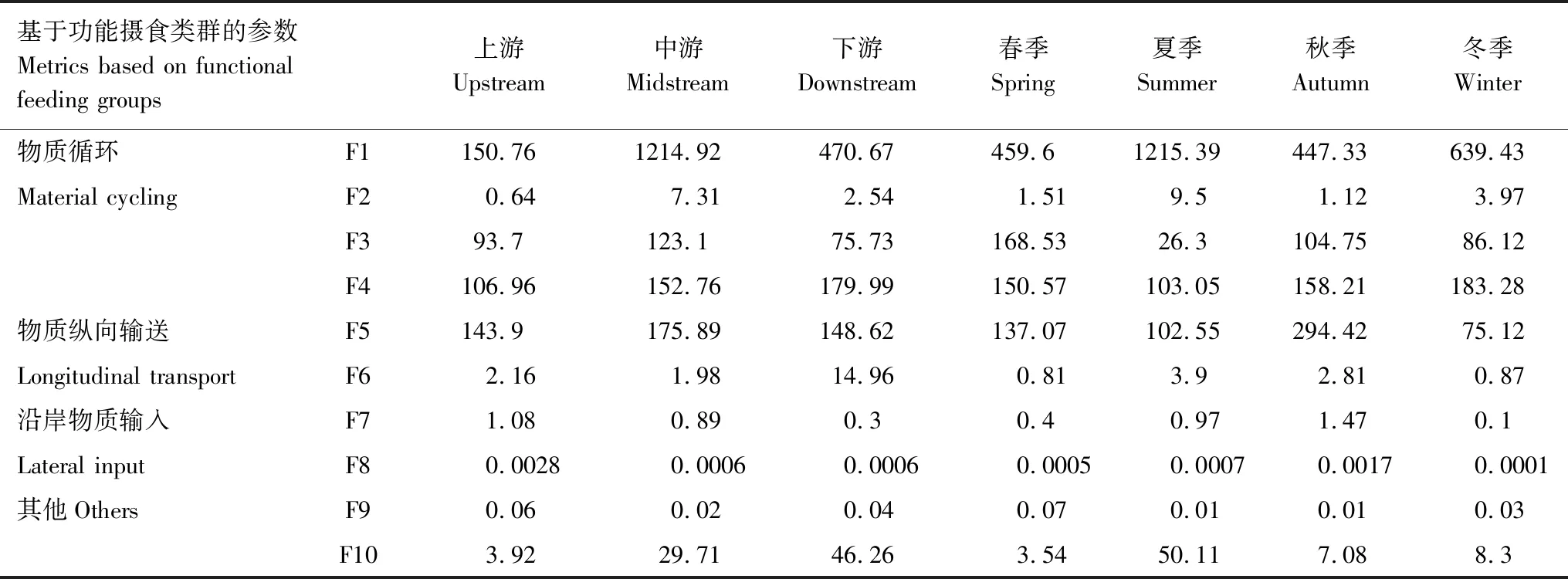
表3 漓江大型底棲無脊椎動物功能攝食類群參數的時空變化
F1:刮食者密度 Density of scrapers;F2:刮食者密度與濾食者、收集者之和的比率 Ratio of scrapers to filterers and gathering-collectors;F3:撕食者密度和收集者密度之和 Density of shredders and gathering-collectors;F4:生物量 Biomass;F5:濾食者密度 Density of filterers;F6:濾食者與撕食者和收集者之和的比率 Ratio of filterers to shredders and gather-collectors;F7:撕食者密度 Density of shredders;F8:撕食者密度與總密度之比 Ratio of shredders to total density;F9:捕食者密度與總密度之比 Ratio of predators to total density;F10:刮食者和濾食者之和與撕食者和收集者之和的比率 Ratio of scrapers and filterers to total shredders and gather-collectors
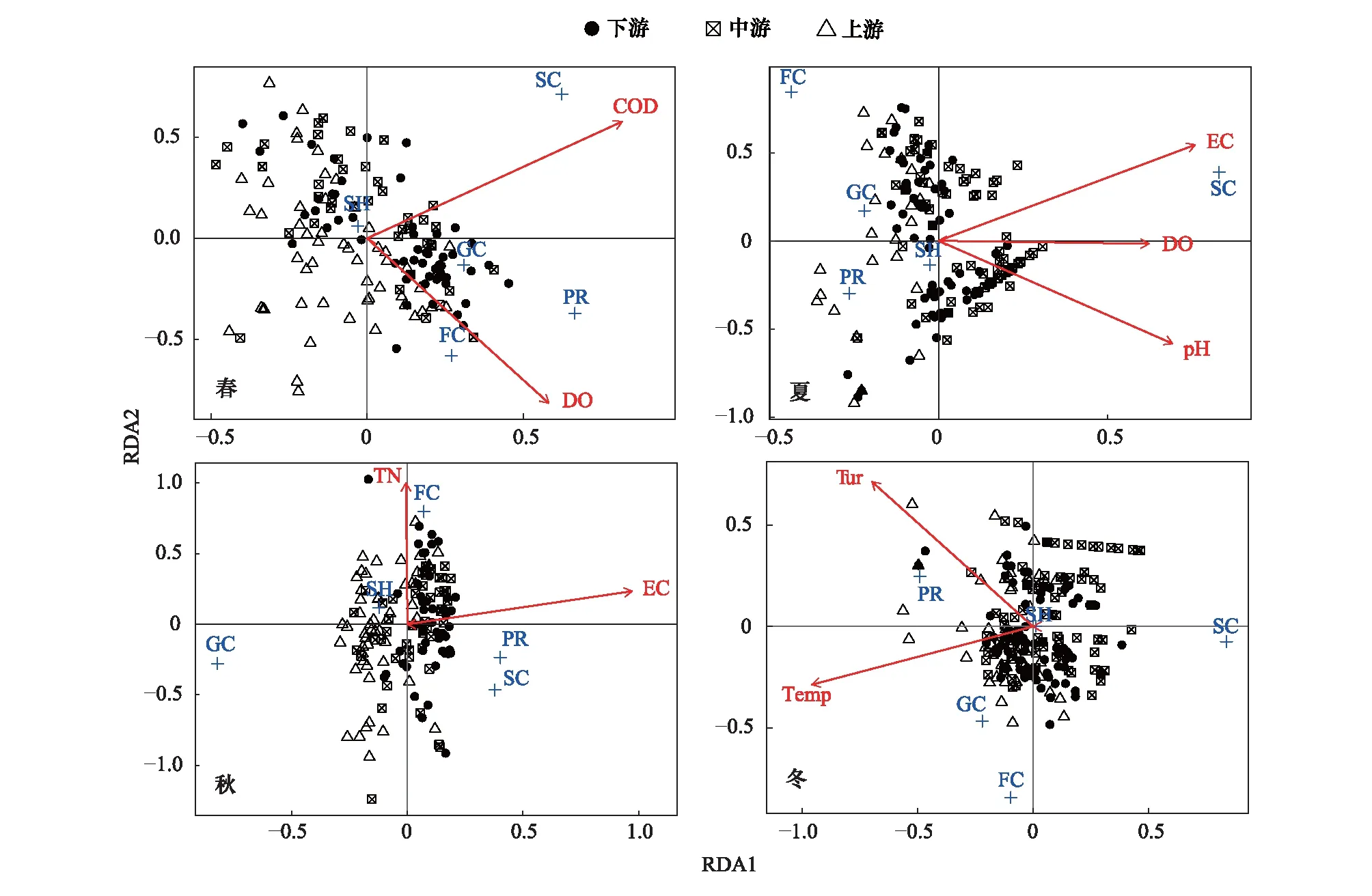
圖6 漓江各季節大型底棲動物功能攝食類群與環境因子的冗余分析(RDA)Fig.6 Relationships of macroinvertebrate FFGs and environmental factors obtained by Redundancy analysis (RDA) in different seasons in Lijiang RiverRDA1:冗余分析第一軸 Redundancy analysis first axis;RDA2:冗余分析第二軸 Redundancy analysis second axis;COD:化學需氧量 Chemical oxygen demand;DO:溶解氧 Dissolved oxygen;EC:電導率 Electrical conductivity;TN:總氮 Ttotal nitrogen;Tur:濁度 Turbidity;Temp:溫度 Temperature
3 討論
3.1 漓江大型底棲無脊椎動物功能攝食類群的組成與時空分布
大型底棲動物功能攝食類群的組成與生境密切相關。漓江大型底棲動物以刮食者占據絕對優勢,撕食者處于劣勢,這一結果與國內其他研究區域如香溪河[22]、昌江[23]等地區的研究結果存在差異。這是由于漓江流域屬于喀斯特地貌,河道底質多為沙卵石加細沙,水淺流急,四季清澈見底,適宜沉水植物生長。漓江茂盛的沉水植物[24]為螺類等刮食者提供了適宜的生境[25],使得刮食者在組成上占據優勢。撕食者數量少的原因是漓江兩岸多為河漫灘與深切河谷,灌喬木稀少,無法給撕食者提供枯枝落葉和蔭蔽的條件,進而撕食者難以生存[26]。
河流連續性概念認為功能攝食類群的空間分布與自然生境中有機物沿河流的運輸、利用和存儲有密切的關系[27],但人類活動對其空間上的分布也產生了較大的影響。按照經典的河流連續性概念預測,對溪流來講,上游河流中撕食者的相對豐度較大[28],捕食者在河流上中下游空間上變化不大[26],刮食者在中游河段的相對豐度最大[29]。本研究觀測結果與上述情況基本吻合。在自然河流中,細顆粒有機物在很大程度上是由粗顆粒有機物分解形成的,其比例自上游至下游逐漸增加,按河流連續性概念預測濾食者和收集者的相對豐度也應自上游至下游逐漸增加[30]。但在本研究中,這兩者的相對豐度表現為上游最大、中下游符合上述規律,這與預測情況偏差較大,原因是本研究所觀測到的濾食者和收集者的優勢種搖蚊幼蟲、小蜉和河蜆均為耐污種,而上游研究區位于漓江支流甘棠江的下游河段,該河段流經村鎮,大量使用的化肥、村莊的生活污水這些有機物輸入到河流中,為一些濾食者和收集者的生長和繁榮創造了條件[29],并致使耐污種比例增高;中下游研究區域均為旅游觀光區,兩岸基本無污染排放。這也與功能攝食類群參數F10相對應,可推斷村鎮居民生活對漓江生態壞境的影響大于旅游觀光的影響。
底棲動物功能攝食類群的季節性分布主要是由于季節變化改變了岸邊植被狀態及河流水文條件,進而使底棲動物的生境發生了微變化,最終導致其時間分布上的差異性。撕食者的分布能夠反映樹葉在河道底部的堆積情況[31]。在本研究中,撕食者的相對豐度和密度在秋季明顯高于其他3個季節,主要是因為樹木的落葉期主要集中在秋季。捕食者的相對豐度和密度在春季高于其他季節,是因為春季其他類群密度較大,為捕食者提供了豐富的食物[9]。刮食者的相對豐度和密度在夏季明顯高于其他季節,主要是因為在夏季繁殖旺盛的中華圓田螺[32]為該功能類群的優勢種,夏季沉水植物的生長也為其提供了適宜的生境。收集者主要以沉積的有機顆粒為食,在流速低的區域有機顆粒較易沉積,因而收集者偏愛流速較低的生境[33]。本研究中收集者的相對豐度和密度在春、秋和冬季無太大差異,在夏季卻很低,這主要是因為夏季是汛期,河流流速增大,不適宜收集者進食。濾食者以懸浮的細有機顆粒物為食,一般都存在一個最適濾食流速[22],在本研究中濾食者相對豐度和密度在秋季明顯高于其他季節,是因為濾食者密度主要受到水流流速及食物影響,秋季處于洪水剛過的時期,水面趨于平穩、流速緩慢,而且撕食者的優勢季節也處在這一時期,其取食粗顆粒有機質后為濾食者提供了更多的細顆粒有機質[26]。
3.2 大型底棲動物無脊椎功能攝食類群與水環境因子之間的關系
在河流生態學研究中確定影響大型底棲動物功能攝食類群的關鍵環境因子對分析河流生態健康起著重要作用[34]。研究指出,環境因子對底棲動物的影響在不同區域不同季節各有差異,較為復雜[34-35]。本研究中RDA分析得出7個環境因子(DO、pH、EC、Temp、Tur、COD、TN)對漓江大型底棲動物功能攝食類群組成與分布具有關鍵影響,主要反映了水體的有機污染物和物理因素。春季COD與刮食者呈顯著的正相關性,這主要是因為COD反映了水中有機質的含量或者說有機物污染程度,而本研究中刮食者是以中華園田螺等耐污種為優勢種,這與其他研究中COD與刮食者的響應關系結論相同[36]。春季和夏季DO與漓江大型底棲動物功能攝食類群相關性較強,這是因為春季和夏季是底棲動物生長和繁殖的季節,其新陳代謝增強,攝食活動會大量消耗水中的溶解氧,所以影響了底棲動物的組成與分布[37]。夏季pH與底棲動物功能攝食類群有較強的相關性,是因為絕大部分底棲動物適宜的pH值為6—8[37-38],而本研究中夏季pH值達到最大為8.49,超出了適宜范圍,對底棲動物的生存產生了影響。天然水體中的pH主要取決于游離二氧化碳的含量及碳酸平衡,夏季植物生長旺盛光合作用強烈致使溶于水體中的二氧化碳減少導致pH值變大。夏季和秋季EC與漓江底棲動物功能攝食類群相關性較強,這是由于電導率是度量水環境總溶解離子量的參數,而離子主要通過影響細胞膜滲透壓的平衡對大型底棲動物產生了影響[39-40]。已有研究指出氮元素與底棲動物功能攝食類群有較好相關性[23,41],本研究中秋季TN與濾食者呈顯著正相關,這與吳東浩等在西苕溪上游得到TN與濾食者呈負相關的結論相反[42],這是因為漓江的濾食者中優勢種河蜆為耐污種。冬季Temp與底棲動物功能攝食類群有較強的相關性,這是因為冬季溫度過低致使底棲動物的活動減弱[37],使得溫度成為冬季限制底棲動物的主要環境因子。不同時期不同區域對底棲動物功能攝食類群起主要影響的環境因子不盡相同,需要充分探究底棲動物的生活習性與生境狀況之間的響應關系。
4 結論
漓江大型底棲無脊椎動物功能攝食類群中以刮食者占據絕對優勢地位,撕食者和捕食者處于劣勢地位。基于大型底棲無脊椎動物功能攝食類群的時空分布特征分析可知,各功能攝食類群的時空分布可反映漓江生境在時空上的微變化及人類活動對生境的干擾,其分布特征明顯受食物資源時空變化的限制。在春季,對功能攝食類群有較強影響的環境因子為COD和DO,夏季為EC、DO和pH,秋季為TN和EC,而在冬季為Tur和Temp。基于功能攝食類群參數對漓江生態系統評價,可知漓江中下游生境穩定性明顯高于上游,村鎮居民生活對漓江生態壞境的影響大于旅游觀光的影響。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
