
熊去氧膽酸通過干擾白介素-10對小鼠肥胖風險的影響
2020-03-17 09:30:18高黎黎張昊
中國美容醫學 2020年1期
高黎黎 張昊
[摘要]目的:探索熊去氧膽酸(UDCA)通過干擾脂肪組織中白介素-10(IL-10)的水平,對高脂飼料誘導小鼠肥胖風險的影響。方法:將C57BL/6J雄鼠簡單隨機化分為:Control組、DIO組、+UDCA組及+UDCA+IL-10組,于誘導0、6及8周后測量體重。取小鼠腸系膜白色脂肪(mesenteric white adipose tissue,mWAT),ELISA和western-blot法檢測mWAT中IL-10和IL-10Rα的蛋白濃度;RT-PCR法檢測產熱基因的表達水平。結果:誘導6周后,Control組小鼠體質量較其它高脂誘導組低,差異有統計學意義(P<0.05);8周后,高脂誘導小鼠中,DIO組小鼠體重最高,+DUCA+IL-10組次之,+DUCA組最低。DIO組mWAT中IL-10含量較Control組高(37.13pg/mg vs 16.29pg/mg, P<0.01),+DUCA組的IL-10含量(19.04pg/mg)較DIO組低(P<0.01),+DUCA+IL-10組的IL-10含量(33.87pg/mg)較+DUCA組高(P<0.05)。此外,各組mWAT中IL-10Rα水平的變化趨勢與IL-10相似。+DUCA組mWAT中多個線粒體產熱基因的表達較DIO組上調(P<0.05)。結論:UDCA可減少高脂誘導小鼠白色脂肪中IL-10和IL-10Rα水平,增加脂肪細胞線粒體產熱基因的表達,降低小鼠的肥胖風險;該效果可被外源性IL-10削弱。
[關鍵詞]熊去氧膽酸;白介素-10;產熱基因;肥胖;風險
[中圖分類號]R589 ? ?[文獻標志碼]A ? ?[文章編號]1008-6455(2020)01-0068-04
Effects of Ursodeoxycholic Acid on the Risk of Obesity in Mice Via Inhibiting Interleukin-10
GAO Li-li1,ZHANG Hao2
(1.Center for Medical Research and Innovation;2.Department of General Surgery,Shanghai Pudong Hospital/Fudan University Pudong Medical Center,Shanghai 201399,China)
Abstract: Objective ?To explore the effect of ursodeoxycholic acid(UDCA) on the risk of obesity via inhibiting interleukin-10(IL-10) level in high-fat diet-induced mice. Methods ?Male C57BL/6 mice were divided into four groups using simple random method, including Control group, DIO group, +UDCA group and +UDCA+IL-10 group, body weight were monitored at baseline, 6 weeks and 8 weeks after feeding. Meanwhile, the mesenteric white adipose tissue (mWAT) was collected, and the IL-10 and IL-10Rα protein level were detected by ELISA and western blot assays respectively, then the expression of thermogenic genes were analyzed by real-time PCR. Results ?Compared with other groups, the body weight value of the Control group was the lowest following 6 weeks induction (P<0.05). After 8 weeks, in high-fat diet-induced mice, the DIO group had the highest body weight, followed by the the +DUCA+IL-10 group and the lowest in the +DUCA group (P<0.05). In addition, the IL-10 level of mWAT in DIO group was higher than that in the Control group(37.13pg/mg vs 16.29pg/mg, P<0.01), it was lower in the +UDCA group(19.04pg/mg) than that in the DIO group (P<0.01), and it was higher in the +UDCA+IL-10 group(33.87pg/mg) than that in the +UDCA group (P<0.05). As well, the trend of IL-10Rα levels among the groups was similar with IL-10. Whereas most of the mitochondrial thermogenic genes abundance were down-regulated in the DIO group, and up-regulated in the +DUCA group (P<0.05). Conclusion ?We conclude that UDCA could reduce IL-10 and IL-10Rα levels in WAT, increase mitochondrial thermogenic genes expression in adipocytes, and finally alleviate the risk of obesity in high-fat-diet-induced mice. Importantly, the treatment of exogenous IL-10 could eliminate the role of UDCA in resisting obesity.
Key words: ursodeoxycholic acid; interleukin-10(IL-10); thermogenic genes; obesity; risk
近年來,一些研究證明了免疫系統對脂肪組織的增殖和活化有明顯影響,強調了抗炎(Ⅱ型)細胞因子在調節脂肪產熱中的重要性。嗜酸性細胞在刺激下產生白細胞介素-4(IL-4)/IL-13,激活產熱過程[1-2]。IL-33激活2型先天淋巴細胞(IL-C2),促進米色脂肪生源論[3]。最近,IL-33被證明通過調節解偶聯蛋白1(uncoupling protein 1,UCPl)的剪接促進脂肪細胞進行非耦合呼吸[4]。白細胞介素-10(interleukin-10,IL-10)是一種多細胞源、多功能的細胞因子[5]。近年來,有研究發現IL-10可通過調節脂肪細胞的染色質重構,限制白色脂肪組織的產熱和能量消耗[6]。因此,尋找有效藥物阻斷IL-10限制脂肪能耗或將成為治療高脂飲食誘發的代謝紊亂的研究目標。
熊去氧膽酸(UDCA)是親水性膽汁酸(BA)并已作為膽汁淤積和慢性肝炎的保肝藥物;它也是經美國食品藥品管理局批準用于治療原發性膽汁膽管炎(PBC)的一種藥物[7-8]。最近研究顯示,UDCA能調節多個分子靶點,發揮抗炎、降脂和改善糖代謝的作用,具有治療肥胖及肥胖所致的代謝性疾病的潛能[9]。但其具體的作用機制不是十分清楚。隨著現代社會肥胖和肥胖相關代謝性疾病的高發,闡明UDCA作為一種安全、有效的肥胖治療手段的機制備受關注。有研究顯示,UDCA能抑制高脂誘導小鼠肝臟IL-10的表達[10]。那它對脂肪內IL-10的調控如何,能否通過對IL-10水平的干擾影響脂肪組織的能量代謝水平,緩解肥胖。本實驗以高脂誘導小鼠為模型,探索UDCA通過干擾脂肪組織中IL-10的水平,對高脂飼料誘導C57BL/6J小鼠肥胖風險的影響,為研究UDCA減輕機體肥胖的作用機制提供理論依據。
1 ?資料和方法
1.1 動物模型:SPF級雄性C57BL/6J小鼠,8周齡, 體質量(20±2)g,購買自上海斯萊克實驗動物有限責任公司[SCXK(滬)2012-0002]。簡單隨機化分為4組,每組8只:①Control組(普通喂養);②DIO組(高脂誘導,美國,Research Diets,D12492);③+UDCA組(高脂+UDCA誘導);④+UDCA+IL-10組(高脂+UDCA+IL-10誘導)。從第7周起,對+UDCA組、+UDCA+IL-10組行UDCA灌胃2周(50mg/kg,1次/d,UDCA原液溶解在33%乙醇,33%二甲亞砜,33%吐溫-80中,使用時按1:50稀釋于0.5M NaCl溶液;UDCA,Sigma-Aldrich,U5127)。第8周起,對+UDCA+IL-10組行IL-10腹腔注射干預1周(10μg/kg,隔天1次,共計4次;IL-10,PEPRO-TECH,210-10)。干預結束后處死小鼠,取腸系膜脂肪組織(mesenteric white adipose tissue, mWAT)凍存。
1.2 方法
1.2.1 體質量檢測:分別檢測各組小鼠基線和誘導6周及8周后的體質量,檢測前禁食6h,正常給水。
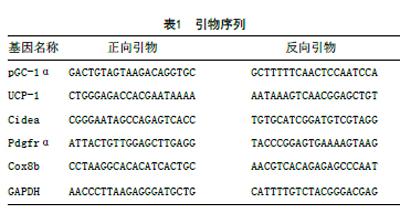
1.2.2 Real-time PCR:按說明書將儲于液氮的mWAT取50g研磨成勻漿后,提取總RNA并測其濃度及A260:A280值,取比值1.8~2.0者,然后逆轉錄成cDNA,采用兩步法PCR擴增標準程序:預變性,95℃ 10min,1個循環進行cDNA變性;PCR反應:95℃ 15s,59℃ 1min。40個循環放大;熔化曲線分析(60℃~95℃),以驗證單個產品的存在。引物由上海生工生物技術有限公司合成。內參為GAPDH基因,使用 delta-delta Ct方法計算mRNA水平:2-ΔΔCT,ΔΔCT=(Ct目標基因-CtGAPDH)實驗組-(Ct目標基因-CtGAPDH)對照組,見表1。
1.2.3 Western-blot法:組織勻漿后,14 000r/min離心5min提取蛋白,取上清定量后加入上樣緩沖液于8%的SDS-PAGE,完畢后轉膜。牛奶封閉后,分別加入1:1 000的IL-10Rα抗體(ab225820,abcam)及1:1 000的β-actin抗體(4967L,CST)4℃孵育過夜后,TBST 漂洗2遍,加入用辣根過氧化物酶標記的二抗,室溫輕搖1h,洗膜后曝光。
1.2.4 ELISA法:組織樣本mWAT(0.3g)置于800μl預冷提取緩沖液(100mM Trizma Base pH7.5, 10mM EDTA,100mM NaF,10mM Na4P2O7,10mM Na3VO4,2mM PMSF,0.1mg/ml 抑肽酶)。勻漿后,添加80μl 10% Triton X-100至樣品,冰上放置30min,4℃離心40min(20817g)。吸取上清,以牛血清白蛋白為參照,采用Bradford法測定蛋白濃度。定量測定IL-10蛋白濃度,ELISA(DuoSet ELISA、R&D Systems,Minneapolis,MN,USA),重復測量,取平均值。
1.3 統計學分析:采用SPSS 22.0軟件進行數據分析,數據采用x?±s形式表示,多組間比較采用單因素方差分析,組間兩兩比較采用t檢驗,檢驗水準α=0.05。采用GraphPad Prism 7軟件以x?±s形式作圖。
2 ?結果
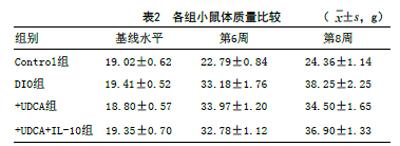
2.1 各組小鼠體質量比較:各組小鼠的體質量基線水平比較差異無統計學意義(F=1.79,P>0.05)。造模8周后,與Control組比較,DIO組小鼠體質量增長較為明顯,差異有統計學意義(t=16.86,P<0.001);與DIO組相比,+UDCA組體質量降低(t=4.55,P<0.001);與UDCA組相比,+UDCA+IL-10組體質量增加,差異有統計學意義(t=2.91,P<0.05)。見表2。
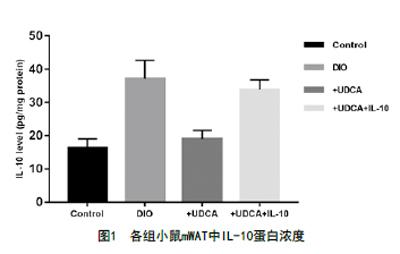
2.2 各組小鼠mWAT中IL-10濃度比較:ELISA測定mWAT的IL-10蛋白濃度,結果顯示DIO組IL-10含量為37.13pg/mg,較Control組16.29pg/mg增高(t=4.04,P<0.01),說明高脂誘導可以引起小鼠mWAT中IL-10含量的增高;+DUCA組IL-10含量為19.04pg/mg,較DIO組下降(t=3.51,P<0.01),說明+UDCA干預可以引起小鼠mWAT中IL-10的含量減低;+DUCA+IL-10組IL-10含量為33.87pg/mg,較+DUCA組增加(t=2.87,P<0.05),腹腔IL-10注射可提高mWAT中IL-10的濃度。見圖1。
2.3 各組小鼠mWAT中IL-10Rα表達水平:Western-blot檢測結果顯示,DIO組mWAT中IL-10Rα的蛋白水平較Control組和+DUCA組高,+DUCA+IL-10組IL-10Rα的蛋白水平較Control組和+DUCA組高,與IL-10含量的變化趨勢相似,見圖2。
2.4 各組小鼠mWAT中線粒體產熱基因的表達: 以各組小鼠mWAT中UCP1、Cidea、Cox8b、pGC-1α及Pdgfrα基因表達水平作為脂肪產熱指標,RT-PCR結果顯示,與Control組相比,DIO組中UCP1、Cidea及pGC-1α的表達下調,差異有統計學意義(P<0.01);與DIO組相比,+UDCA組中UCP1、Cidea、Cox8b及pGC-1α的表達上調,差異有統計學意義(P<0.05);與+UDCA組相比,+UDCA+IL-10組中UCP1、Cidea、Cox8b及pGC-1α的表達下調,差異有統計學意義(P<0.05)。UDCA干預能提高高脂誘導小鼠mWAT中線粒體UCP1的水平,促進產熱相關基因的表達,IL-10干預會減弱這種效應,見圖3。
3 ?討論
IL-10是目前公認的炎癥與免疫抑制因子,主要通過信號轉導及轉錄活化因子3 (signal transducer and activator of transcription 3,STAT3)發揮抗炎與免疫抑制作用[11]。關于IL-10對脂肪和能量穩態作用的觀點卻不同。Mauer等認為IL-10不具有抵抗肥胖的功能[12-13]。但P Rajbhandari等指出IL-10能參與抑制脂肪細胞產熱基因的表達,促進飲食誘導肥胖的發生;IL-10Rα是富集于白色和米色脂肪細胞表面的IL-10受體的亞單位,并隨著分化水平和機體肥胖程度逐漸上調[6]。本研究顯示高脂誘導能增加小鼠白色脂肪mWAT中IL-10濃度和IL-10Rα的表達,與后者研究結果基本一致。
在哺乳動物體內,存在3種類型脂肪組織:白色脂肪組織(White adipose tissue,WAT)、棕色脂肪組織(Brown adipose tissue,BAT)和米色脂肪組織。其中,一般情況下WAT以甘油三酯的形式儲存能量,線粒體含量少。BAT含有豐富的線粒體,線粒體中存在的UCP1可將部分用于制造ATP的能量轉化為熱量。然而,人們在WAT中發現了米色脂肪組織與BAT相似,米色脂肪細胞的生成通常伴隨著機體的產熱作用增強,脂肪代謝增加及體質量減輕[14]。米色脂肪組織可由前體細胞和白色脂肪細胞轉分化而來,表達高水平的UCP1[15-16]。因此,將豐富的白色脂肪轉化為米色脂肪,促進脂肪細胞線粒體UCP1產熱,是對抗肥胖的有效方法。
研究發現,高脂飲食導致的肥胖大鼠模型體內膽汁酸的水平明顯降低[17],而給予小鼠UDCA后能夠改善高脂飲食誘導的肥胖[18]。一方面,是由于膽汁酸會通過激活TGR5依賴型2型碘甲狀腺原氨酸脫碘酶(D2)來增加小鼠BAT和人類骨骼肌的能量消耗,D2是負責將甲狀腺素(T4)轉化為三碘甲狀腺原氨酸的主要酶(T3)[19-20]。生物活性T3誘導線粒體生物發生和UCP1的表達,促進游離脂肪酸氧化和增加能量消耗[21-22]。另一方面,本文通過動物實驗探索發現,UDCA對限制熱量消耗的分子有調節作用。UDCA干預可降低IL-10及IL-10Rα蛋白水平,提高線粒體部分產熱基因的表達,緩解IL-10對肥胖小鼠脂肪能量消耗的抑制;但外源性IL-10的輸入又會增加小鼠白色脂肪中IL-10和IL-10Rα,逆轉UDCA抵抗肥胖風險的作用。
綜上所述,UDCA干預可以降低高脂誘導小鼠白色脂肪中IL-10和IL-10Rα水平,增加脂肪細胞線粒體產熱基因的表達,預防小鼠的肥胖風險,該效果可被外源性IL-10削弱。
[參考文獻]
[1]Qiu Y,Nguyen KD,Odegaard JI,et al.Eosinophils and type 2 cytokine signaling in macrophages orchestrate development of functional beige fat[J].Cell,2014,157(6):1292-1308.
[2]Rao RR,Long JZ,White JP,et al.Meteorin-like is a hormone that regulates immune-adipose interactions to increase beige fat thermogenesis[J].Cell,2014,157(6):1279-1291.
[3]Lee MW,Odegaard JI,Mukundan L,et al.Activated type 2 innate lymphoid cells regulate beige fat biogenesis[J].Cell,2015,160(1-2):74-87.
[4]Odegaard JI,Lee MW,Sogawa Y,et al.Perinatal licensing of thermogenesis by IL-33 and ST2[J].Cell,2016,166(4):841-854.
[5]Saraiva M,OGarra A.The regulation of IL-10 production by immune cells[J].Nat Rev Immunol,2010,10(3):170-181.
[6]Rajbhandari P,Thomas BJ,Feng AC,et al.IL-10 signaling remodels adipose chromatin architecture to limit thermogenesis and energy expenditure[J].Cell,2018,172(1-2):218-233.
[7]Ali AH,Tabibian JH,Carey EJ,et al.Emerging drugs for the treatment of Primary Biliary Cholangitis[J].Expert Opin Emerg Drugs,2016,21(1):39-56.
[8]Ratziu V,De Ledinghen V,Oberti F,et al.A randomized controlled trial of high-dose ursodesoxycholic acid for nonalcoholic steatohepatitis[J].J Hepatol,2011,54(5):1011-1019.
[9]Lefebvre P,Cariou B,Lien F,et al.Role of bile acids and bile acid receptors in metabolic regulation[J].Physiol Rev,2009,89(1):147-191.
[10]Chen YS,Liu HM,Lee TY.Ursodeoxycholic acid regulates hepatic energy homeostasis and white adipose tissue macrophages polarization in leptin-deficiency obese mice [J].Cells,2019,8(3).pii:E253.
[11]Moore KW,De Waal Malefyt R,Coffman RL,et al.Interleukin-10 and the interleukin-10 receptor[J].Annu Rev Immunol,2001,19:683-765.
[12]Mauer J,Chaurasia B,Goldau J,et al.Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin[J].Nat Immunol,2014,15(5):423-430.
[13]Miller AM,Wang H,Bertola A,et al.Inflammation-associated interleukin-6/signal transducer and activator of transcription 3 activation ameliorates alcoholic and nonalcoholic fatty liver diseases in interleukin-10-deficient mice[J].Hepatology,2011,54(3):846-856.
[14]鄧露露,歐子豪,彭永正.米色脂肪細胞的誘導因子—抗肥胖的新策略[J].實用醫學雜志,2017,33(19):3162-3165.
[15]Wu J,Bostrom P,Sparks LM,et al.Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human[J].Ce11,2012,150(2):366-376.
[16]Schulz TJ,Huang P,Huang TL,et al.Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat[J].Nature,2013,495(7441):379-383.
[17]李德剛,高敏,沙妙青,等.從膽汁流量和膽汁酸成分改變初探“膽道不利是肥胖發病的中心環節”[J].世界中醫藥,2014,9(3):350-354.
[18]馬慧娟,劉晶,張柳,等.膽汁酸對高脂誘導肥胖小鼠的血糖和體重的影響及安全性分析[J].解放軍醫藥雜志,2015,27(10):25-28.
[19]Watanabe M,Houten SM,Mataki C,et al.Bile acids induce energy expenditure by promoting intracellular thyroid hormone activation[J].Nature,2006,439(7075):484-489.
[20]Thomas C,Gioiello A,Noriega L,et al.TGR5-mediated bile acid sensing controls glucose homeostasis[J].Cell Metab,2009,10(3):167-177.
[21]Klingenspor M,Ivemeyer M,Wiesinger H,et al.Biogenesis of thermogenic mitochondria in brown adipose tissue of Djungarian hamsters during cold adaptation [J].Biochem J,1996,316(Pt 2):607-613.
[22]Bianco AC,Sheng XY,Silva JE.Triiodothyronine amplifies norepinephrine stimulation of uncoupling protein gene transcription by a mechanism not requiring protein synthesis[J].J Biol Chem,1988,263(34):18168-18175.
[收稿日期]2019-05-20
本文引用格式:高黎黎,張昊.熊去氧膽酸通過干擾白介素-10對小鼠肥胖風險的影響[J].中國美容醫學,2020,29(1):68-71.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
