高郵湖大銀魚、太湖新銀魚Cytb和COⅠ基因序列多態性分析
2020-03-20 03:37:02李大命孫文祥唐晟凱劉燕山谷先坤劉小維張彤晴
水產科學 2020年2期
李大命,孫文祥,許 飛,唐晟凱,劉燕山,谷先坤,劉小維,張彤晴
( 1.江蘇省淡水水產研究所,江蘇省內陸水域漁業資源重點實驗室, 江蘇 南京 210097; 2.江蘇省高寶邵伯湖漁業管理委員會辦公室,江蘇 揚州 225009 )
大銀魚(Protosalanxhyalocranius)和太湖新銀魚(Neosalanxtaihuensis)均為一年生小型魚類,分別屬鮭形目、銀魚科的大銀魚屬和新銀魚屬,主要分布在我國黃河、淮河和長江中下游及其附屬湖泊[1]。銀魚營養價值和經濟價值均很高,是我國重要的經濟魚類。銀魚是典型的r-選擇物種,個體小,對環境變化敏感,種群消長快。近幾十年來,由于圍湖造田、過度捕撈、環境污染等多種不利因素的影響,銀魚天然資源持續衰退,分布范圍顯著縮小,個別物種漸危[2]。因此,加強銀魚資源保護勢在必行。
目前,有關銀魚的研究主要集中在移植增殖、生殖發育、種群生態、營養價值等方面[3-9],而有關銀魚的分子生物學研究資料較少。遺傳多樣性決定了物種的進化潛能和對環境變化的適應能力,是生物多樣性的一個重要方面。魚類線粒體DNA是一種閉合、環狀的雙鏈DNA分子,具有結構簡單、母系遺傳、不發生重組及進化速度快等特征,使其成為分子群體遺傳學和分子系統學研究的重要分子標記[10]。線粒體細胞色素b (Cytb)和細胞色素c氧化酶亞基Ⅰ(COⅠ)基因是線粒體DNA的蛋白質編碼序列,具有進化速度較快、易擴增等優點,適用于種群水平遺傳多樣性的檢測,已被廣泛地應用于魚類分子系統學和種群遺傳學研究[11-13]。
高郵湖是全國第六大淡水湖、江蘇省第三大淡水湖,水域面積780 km2。高郵湖具有漁業生產、工農業及生活用水、航運、旅游、泄洪行洪等多種功能,是具有豐富的生物多樣性和較高生產力的生態系統[14]。高郵湖有魚類63種,其中銀魚是高郵湖重要的漁業資源之一,也是漁民主要的捕撈對象[15]。近幾十年來,受生態環境污染、不合理漁業生產方式及涉湖、涉漁工程建設的影響,高郵湖漁業資源量顯著下降,漁業可持續發展受到嚴重威脅[15]。迄今為止,有關高郵湖魚類的遺傳學資料較少。有文獻報道了高郵湖日本沼蝦(Macrobrachiumnipponense)和湖鱭(Coilianasus)群體的遺傳多樣性水平[16-17],但尚未見有關銀魚遺傳多樣性的研究報道。本研究采用線粒體DNA Cytb和COⅠ基因序列作為分子標記,對高郵湖大銀魚和太湖新銀魚2個銀魚種群遺傳多樣性開展研究,一方面有利于豐富銀魚的分子生物學資料,另一方面可以為高郵湖銀魚種質資源保護及合理利用提供科學依據。
1 材料與方法
1.1 樣本采集和處理
2018年8—12月,在高郵湖水域采集大銀魚和太湖新銀魚各40尾。經測量,大銀魚體質量1.6359~5.6104 g,平均為3.3236 g;體長7.33~11.09 cm,平均為9.08 cm。太湖新銀魚體質量0.3674~1.2067 g,平均為0.6737 g;體長4.12~6.19 cm,平均為4.93 cm。剪取每尾個體的背部肌肉組織放入裝有無水乙醇的離心管中,帶回實驗室置于冰箱-20 ℃保存備用。
1.2 DNA提取、PCR擴增及測序
剪取銀魚肌肉組織約50 mg,超純水沖洗2次,用滅菌吸水紙吸干水分。采用廣譜性基因組提取試劑盒(Takara,中國)提取DNA。用1.0%瓊脂糖凝膠電泳和紫外分光光度計檢測DNA質量和含量,-20 ℃保存備用。
大銀魚、太湖新銀魚線粒體Cytb基因擴增引物為:L14321,5′-CCAGTGACTTGAAAAACCACCG-3′,H15634,5′-CTTAGCTTTGGGAGTTAAGGGT-3′[18]; COⅠ基因擴增引物為:F1,5′-TCAACCAAC CACAAAGACATTGGCAC-3′,R1,5′-TAGACTTC TGGGTGGCCAAAGAATCA-3′[19]。引物由上海生工生物工程有限公司合成。PCR反應總體積為50 μL,其中10×Buffer 5 μL,dNTP(10 mmol/L) 5 μL,正反引物各2 μL(10 μmol/L),Taq DNA聚合酶(5 U/μL) 0.4 μL,DNA模板25 ng,ddH2O補足體積。樣品在Eppendorf PCR儀上進行擴增。Cytb基因的反應條件:94 ℃預變性4 min,94 ℃變性40 s,55 ℃退火40 s,72 ℃延伸90 s,30個循環,72 ℃最后延伸10 min。COⅠ基因的反應條件:94 ℃預變性4 min,94 ℃變性40 s,56 ℃退火40 s,72 ℃延伸60 s,30個循環,72 ℃最后延伸5 min。
PCR擴增產物用1.5%瓊脂糖凝膠電泳檢測,送上海生工生物工程有限公司進行雙向測序,測序引物與擴增引物相同。
1.3 數據分析
利用BioEdit軟件[20]對測序所得序列進行拼接并輔以人工校對,通過BLAST (https:∥www.ncbi.nlm.nih.gov/BLAST/)檢索確定為目的片段。應用Mega 7.0軟件[21]對所有序列進行多重比對,統計堿基組成及變異位點。采用Kimura雙參數模型計算單倍型間的遺傳距離,基于鄰接法構建單倍型分子系統進化樹。利用DnaSP 5.1軟件[22]計算單倍型數、單倍型多樣性指數和核苷酸多樣性指數。應用Arlequin 3.5.1.2軟件[23]進行Tajima′sD和 Fu′sFs中性檢驗,同時結合核苷酸不配對分布圖分析種群歷史動態。
2 結 果
2.1 大銀魚和太湖新銀魚Cytb和COⅠ基因序列變異分析
經PCR擴增、測序和校對,獲得高郵湖大銀魚線粒體Cytb基因全長序列(1141 bp)。Cytb基因序列中堿基組成平均含量為:A(21.7%)、G(16.7%)、C(32.3%)和T(29.3%),堿基A+T的含量(51.0%)略大于堿基G+C的含量(49.0%)(表1)。40條Cytb基因序列檢出14個變異位點,總變異率為1.23%,其中7個是簡約位點,7個是單一位點。沒有插入和缺失位點。經分析獲得高郵湖大銀魚COⅠ基因序列片段(630 bp)。COⅠ基因序列中堿基組成平均含量為:A(21.6%)、G(19.2%)、C(33.2%)和T(26.0%),堿基A+T的含量(47.6%)略小于堿基G+C的含量(52.4%)(表1)。40條COⅠ基因序列檢出5個多態性位點,總變異率為0.79%,其中4個簡約位點,1個單一位點。沒有插入和缺失位點。
經PCR擴增、測序和校對,獲得高郵湖太湖新銀魚線粒體Cytb基因全長序列(1141 bp)。Cytb基因序列中堿基組成平均含量為:A(21.1%)、G(17.4%)、C(33.5%)和T(28.0%),堿基A+T的含量(49.1%)略小于堿基G+C的含量(50.9%)(表1)。40條Cytb基因序列檢出13個變異位點,總變異率為1.14%,其中4個是簡約位點,9個是單一位點。沒有插入和缺失位點。經分析獲得高郵湖太湖新銀魚COⅠ基因序列片段(630 bp)。COⅠ基因序列中堿基組成平均含量為:A(19.5%)、G(20.8%)、C(34.3%)和T(25.4%),堿基A+T的含量(44.9%)小于堿基G+C的含量(55.1%)(表1)。40條CO Ⅰ基因序列檢出2個變異位點,總變異率為0.32%,均為簡約位點。沒有插入和缺失位點。
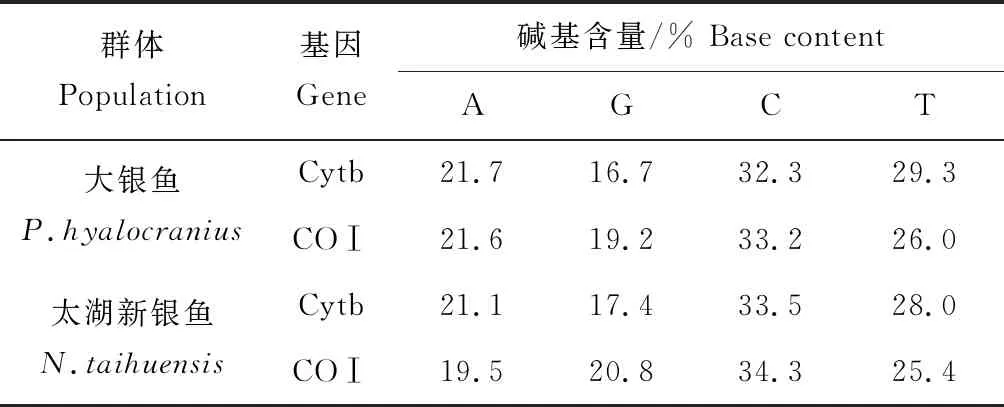
表1 大銀魚和太湖新銀魚Cytb和COⅠ基因序列堿基組成Tab.1 The base composition of Cytb and COⅠgene in icefishP.hyalocranius and N.taihuensis
2.2 大銀魚和太湖新銀魚種群遺傳多樣性參數
40尾大銀魚定義12個Cytb基因單倍型,分別命名為PHB1~PHB12(GenBank登錄號:MK523739~MK523750),其數量分別為11、1、6、2、2、4、1、2、2、7、1和1,其中PHB1是數量最多的單倍型,占比為27.5%。單倍型多樣性和核苷酸多樣性分別為0.871±0.031和0.00172±0.00019,平均核苷酸差異數為1.963(表2);40尾大銀魚定義6個COⅠ基因單倍型,分別命名為PHC1~PHC6(GenBank登錄號:MK523751~MK523756),其數量分別為16、1、8、3、10和2個,其中PHC1是數量最多的單倍型,占比為25.0%。單倍型多樣性和核苷酸多樣性分別為0.747±0.041和0.00202±0.00019,平均核苷酸差異數為1.272(表2)。
40尾太湖新銀魚定義9個Cytb基因單倍型,分別命名為NTB1~NTB8(GenBank登錄號:MK523757~MK523765),其數量分別為24、1、1、1、2、8、1、1和1個,其中NTB1是數量最多的單倍型,占比為60.0%。單倍型多樣性和核苷酸多樣性分別為0.609±0.078和0.00094±0.00027,平均核苷酸差異數為1.071(表2);40尾太湖新銀魚定義3個COⅠ基因單倍型,分別命名為NTC1~NTC3(GenBank登錄號:MK523766~MK523768),其數量分別為35、2和3個,其中NTC1是數量最多的單倍型,占比為87.5%。單倍型多樣性和核苷酸多樣性分別為0.232±0.085和0.00038±0.00014,平均核苷酸差異數為0.240(表2)。
2.3 大銀魚、太湖新銀魚單倍型系統進化樹
高郵湖大銀魚線粒體Cytb和COⅠ基因單倍型間的Kimura雙參數遺傳距離分別為0.001~0.004和0.002~0.006,相應地,太湖新銀魚2個基因單倍型間的遺傳距離分別為0.001~0.006和0.002~0.003。大銀魚和太湖新銀魚Cytb單倍型間的Kimura雙參數遺傳距離為0.124~0.130,相應地,大銀魚和太湖新銀魚COⅠ單倍型間的遺傳距離為0.124~0.130。
采用鄰接法和最大似然法構建大銀魚和太湖新銀魚Cytb和COⅠ基因單倍型分子系統進化樹(圖1、圖2)。結果顯示,2種方法構建的單倍型系統進化關系基本一致,大銀魚、太湖新銀魚的單倍型各自聚為一支,形成兩個獨立分支。
2.4 大銀魚、太湖新銀魚群體歷史動態
對高郵湖大銀魚、太湖新銀魚進行Tajima′sD和Fu′sFs中性檢驗(表3)。結果顯示,除基于COⅠ基因的大銀魚Tajima′sD中性檢驗結果為正值外,大銀魚和太湖新銀魚的2種中性檢測結果均為負值,且Fu′sFs中性檢測結果統計檢驗達到極顯著水平(P<0.01),說明大銀魚和太湖新銀魚進化過程中顯著偏離中性選擇。
對高郵湖大銀魚、太湖新銀魚Cytb和COⅠ基因進行核苷酸錯配分布分析(圖3、圖4)。由圖3、圖4可見,大銀魚、太湖新銀魚Cytb和COⅠ基因序列的核苷酸錯配分布圖均呈單峰型,結合中性檢驗結果,可以推斷出高郵湖大銀魚、太湖新銀魚群體在歷史上經歷過種群擴張。
3 討 論
3.1 高郵湖大銀魚和太湖新銀魚遺傳多樣性分析
遺傳多樣性是物種生存適應和發展的前提,也是評價種群資源狀況的重要依據,一個物種的遺傳多樣性水平越高,其進化的潛力及適應環境的能力也越強。反之,物種遺傳多樣性水平減少可能導致其成活率、生長與繁殖效率變低, 降低種群個體對環境變化的適應能力。衡量一個物種遺傳多樣性的指標主要是單倍型多樣性和核苷酸多樣性,單倍型多樣性和核苷酸多樣性的值越大,說明物種的遺傳多樣性越豐富[24]。本研究中,基于Cytb和COⅠ基因的高郵湖大銀魚單倍型和核苷酸多樣性分別為0.871±0.031、0.00172±0.00019和0.747±0.041、0.00202±0.00019。太湖新銀魚的遺傳多樣性參數分別為0.609±0.078、0.00094±0.00027和0.232±0.085、0.00038±0.00014。高郵湖大銀魚的遺傳多樣性明顯高于太湖新銀魚遺傳多樣性,這可能與兩種銀魚的有效種群大小有關。近幾年的高郵湖漁業資源調查結果顯示,大銀魚資源量明顯大于太湖新銀魚(未發表)。此外,大銀魚和太湖新銀魚的分布范圍、生態位、繁殖力、生長速率等方面的差異導致大銀魚有更強的環境適應能力[1-2]。根據Grant等[25]提出的標準,單倍型多樣性和核苷酸多樣性高低分別以0.5和0.005為臨界值。大銀魚的遺傳多樣性具有高單倍型多樣性和低核苷酸多樣性特征;基于Cytb的太湖新銀魚遺傳多樣性具有較高的單倍型多樣性和較低的核苷酸多樣性特征,而基于COⅠ的太湖新銀魚呈現出極低的單倍型多樣性和極低的核苷酸多樣性。已有文獻報道了我國不同水域大銀魚群體[12,26-29]和太湖新銀魚群體[26,30-31]的遺傳多樣性水平。通過比較可以得出,高郵湖大銀魚的遺傳多樣性處于中等水平,而太湖新銀魚的遺傳多樣性處于較低水平。另外可以看出,基于Cytb的大銀魚和太湖新銀魚的遺傳多樣性水平高于基于COⅠ的大銀魚和太湖新銀魚的遺傳多樣性水平,這是因為Cytb基因序列變化速率比COⅠ快,蘊含了更多的遺傳信息,與文獻報道結果一致[12,32-34]。
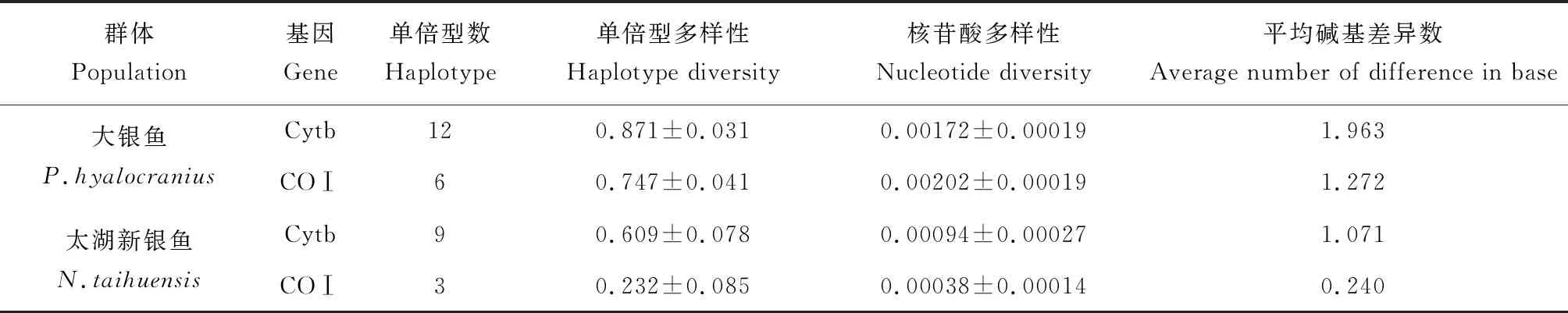
表2 大銀魚、太湖新銀魚種群遺傳多樣性參數Tab.2 Parameters of genetic diversity in populations of icefish P.hyalocranius and N.taihuensis
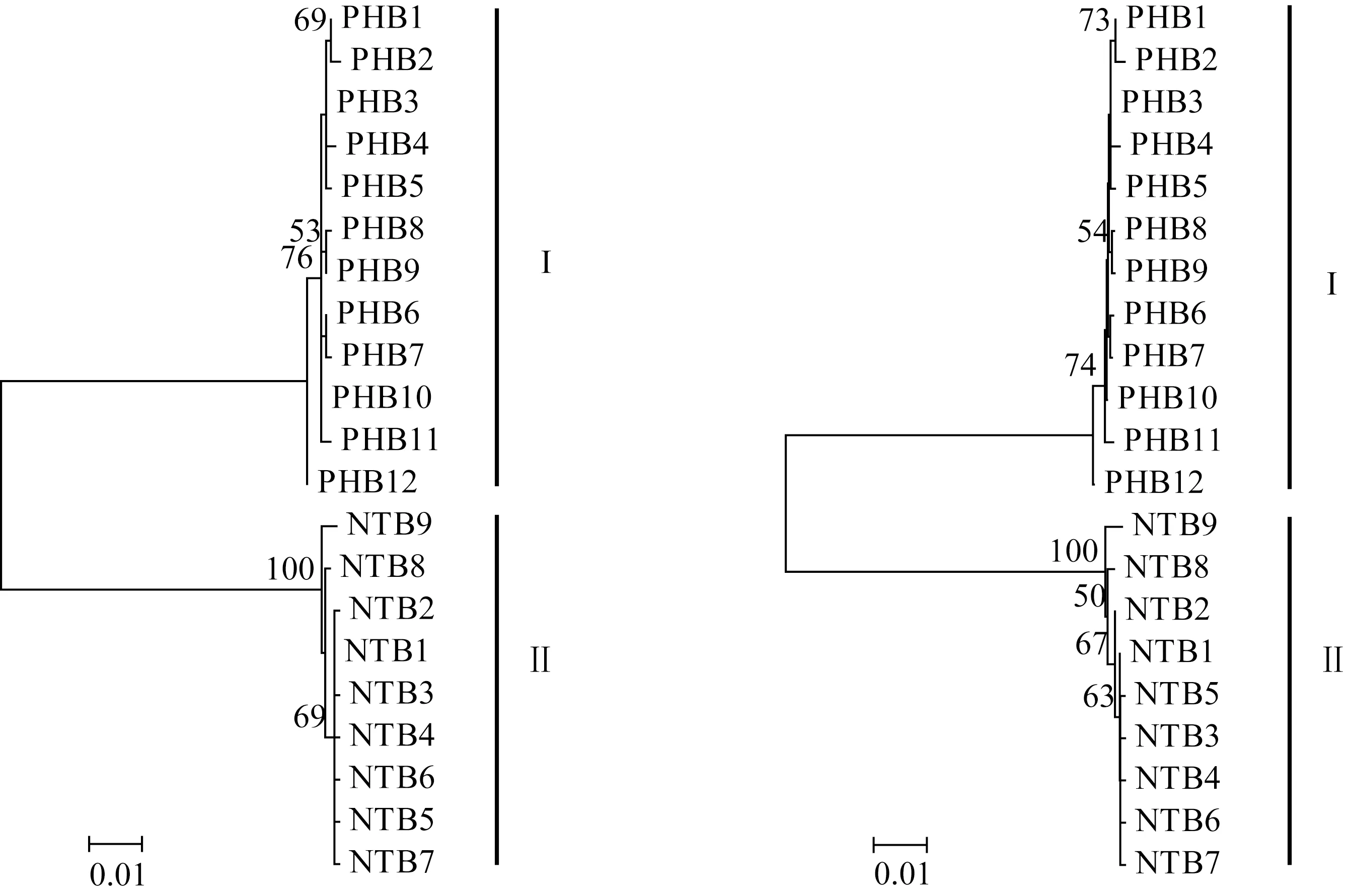
圖1 大銀魚和太湖新銀魚Cytb基因單倍型系統進化樹Fig.1 Phylogenetic tree of Cytb gene haplotypes of icefish P.hyalocranius and N.taihuensis
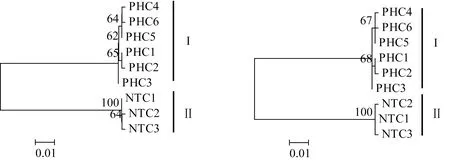
圖2 大銀魚和太湖新銀魚COⅠ基因單倍型系統進化樹Fig.2 Phylogenetic tree of COⅠ gene haplotypes of icefish P.hyalocranius and N.taihuensis
表3大銀魚和太湖新銀魚中性檢驗結果
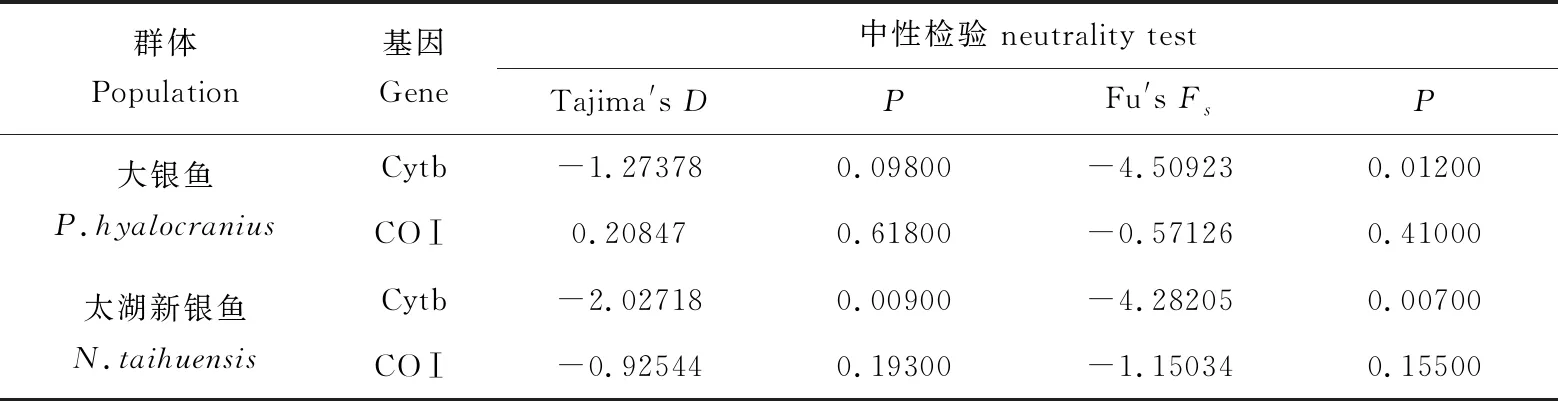
Tab.3 The results of neutrality test of icefish P.hyalocranius and N.taihuensis
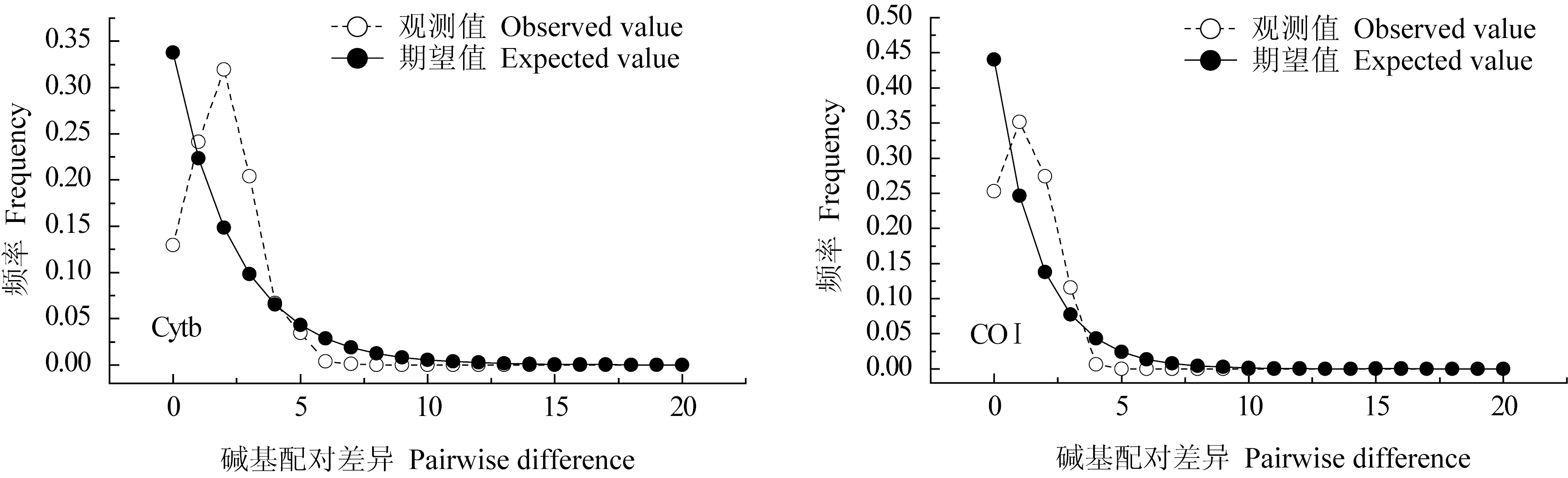
圖3 大銀魚核苷酸錯配分布曲線Fig.3 Mismatch distribution of icefish P.hyalocranius
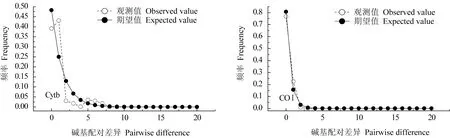
圖4 太湖新銀魚核苷酸錯配分布曲線Fig.4 Mismatch distribution of icefish N.taihuensis
魚類的遺傳多樣性特征與其進化歷史密切相關。Grant等[25]提出一個簡單模式,利用單倍型多樣性和核苷酸多樣性來估計種群的進化歷史:當單倍型多樣性大于等于0.5、核苷酸多樣性小于0.05時,是受瓶頸效應后種群數量的迅速擴張導致;當單倍型多樣性大于等于0.5、核苷酸多樣性大于0.005時,表示種群穩定,具有比較悠久的進化歷史;當單倍型多樣性小于0.5、核苷酸多樣性大于等于0.005時,種群經歷了輕微的瓶頸效應,幾乎沒有影響到核苷酸變異;當單倍型多樣性小于0.5、核苷酸多樣性小于0.005時,表明種群近期經歷了瓶頸效應。本研究結果表明,高郵湖大銀魚遺傳多樣性具有高單倍型多樣性和低核苷酸多樣性,屬于第一種進化模式。核苷酸多樣性積累所需時間比積累單倍型所需的時間長,這說明高郵湖大銀魚是由一個有效種群數量較小的種群,經快速擴張而來,但是仍然沒有達到積累核苷酸變異所需要的時間。太湖新銀魚單倍型多樣性略高于或明顯低于0.5,核苷酸多樣性明顯低于0.005,整體來看呈現出較低的遺傳多樣性水平,說明太湖新銀魚種群經歷了瓶頸效應,導致遺傳多樣性水平較低。另外,通過Tajima′sD和Fu′sFs中性檢驗也可以推斷種群進化歷史。當Tajima′sD和Fu′sFs呈負值,并且在統計學上有較顯著的標準,說明被研究種群經歷過一個擴張的歷史[35-35]。本研究中高郵湖大銀魚和太湖新銀魚的Tajima′sD和Fu′sFs值為負數,且基于Cytb基因的統計結果具有極顯著性。同時,高郵湖大銀魚和太湖新銀魚的核苷酸歧點分布曲線呈單峰型,均說明高郵湖大銀魚和太湖新銀魚在歷史上經歷過種群擴張。
3.2 高郵湖大銀魚和太湖新銀魚遺傳結構差異
研究表明,高郵湖大銀魚Cytb和COⅠ基因序列單倍型間的遺傳距離分別為0.001~0.004和0.002~0.006,相應地,太湖新銀魚2個基因單倍型間的遺傳距離分別為0.001~0.006和0.002~0.003。由此得出,高郵湖大銀魚、太湖新銀魚種群內單倍型之間的遺傳距離非常小,表明兩個種群內個體間尚未產生遺傳分化。另外,大銀魚和太湖新銀魚的種群遺傳結構明顯不同:一方面,兩個種群的單倍型數量數不同;另一方面,優勢單倍型所占比例差異明顯,基于Cytb和COⅠ的大銀魚優勢單倍型所占比例分別為27.5%和25.0%,相應地,太湖新銀魚優勢單倍型所占比例分別為60.0%和87.5%。這種差異導致太湖新銀魚遺傳多樣性較低,種群結構不穩定,適應環境能力下降。
3.3 高郵湖銀魚資源保護
本研究基于線粒體DNA Cytb和COⅠ基因序列研究了高郵湖大銀魚、太湖新銀魚遺傳多樣性水平和遺傳結構組成。大銀魚的遺傳多樣性處于中等水平,種群遺傳結構較為穩定;太湖新銀魚的遺傳多樣性處于較低水平,種群遺傳結構不穩定。結合銀魚的生物學特點,需要采取多種措施保護高郵湖銀魚資源:控制環境污染,修復和改善銀魚棲息環境;嚴格控制捕撈強度,減少人類活動干擾;開展人工魚苗和增殖放流,增加銀魚資源量,優化種群遺傳結構,提高遺傳多樣性水平,增強高郵湖銀魚資源可持續發展能力。
