不同UV-B輻射對杧果樹果實品質及其抗氧化的影響
2020-03-23 06:07:38王紅楊成坤岳堃郭鈺柬周開兵
熱帶作物學報 2020年2期
王紅 楊成坤 岳堃 郭鈺柬 周開兵
摘? 要:以‘臺農一號杧果樹為試驗材料,自然光為對照(CK),研究不同UV-B輻射[24、96 kJ/(m2·d)]對杧果果實的損傷及其抗氧化的影響,并監測株產、果實品質和生理損傷、抗氧化保護機制等有關的生理生化指標的動態變化。結果表明:與CK相比,96 kJ/(m2·d)的杧果樹體株產和單果重均顯著下降,其果實可溶性糖下降、可滴定酸含量升高繼而糖酸比下降,導致果實品質變劣。2個UV-B輻射處理的果皮丙二醛含量與CK無顯著差異,而果肉則顯著高于CK。96 kJ/(m2·d)處理的果皮維生素C和未知化合物含量均顯著高于CK,而過氧化物酶(POD)活性顯著低于CK,對其他抗氧化酶活性和還原性保護性成分含量則一直無顯著影響。2個UV-B輻射處理的果肉抗氧化酶活性及還原型GSH含量隨時間均呈先上升后下降的趨勢。與CK相比,2個UV-B輻射處理的果肉超氧化物歧化酶和過氧化氫酶活性均顯著下降,96 kJ/(m2·d)處理的果肉POD活性、類黃酮和還原型GSH含量均顯著低于CK,而24 kJ/(m2·d)處理的POD活性與CK無顯著差異,類黃酮和還原型GSH含量僅在后熟期顯著低于CK,而多酚含量幾乎不受影響。可見,增強UV-B輻射[96 kJ/(m2·d)]對樹體和果肉引起損傷,對果皮無損傷。
關鍵詞:增強UV-B輻射;杧果果實;損傷;抗氧化
中圖分類號:S667.7???? 文獻標識碼:A
Abstract: In order to evaluate the injures and responses of the anti-oxidation of mango fruits under two different doses of enhanced UV-B radiation treatments, the adult trees of the variety ‘Tainong No.?1 in the field were treated with enhanced UV-B radiation at the levels of 24 kJ/(m2·d) and 96 kJ/(m2·d), and those with natural sunlight were used as the control (CK). The dynamic changes of physiological and biochemical indexes related to plant yield, fruit quality and physiological damage, antioxidant protection mechanism, etc. were observed. The results showed that the yield per tree and single fruit weight of 96 kJ/(m2·d) was significantly decreased compared with CK, and the fruit soluble sugar was decreased, titratable acid content was increased, and then the sugar-acid ratio was decreased, resulting in the fruit quality was worse. The content of MDA in the pericarp of two UV-B radiation treatments was not significantly different from CK, but the flesh was significantly higher than the CK. The contents of vitamin C and unknown compounds in pericarp treated with 96 kJ/(m2·d) were significantly higher than the CK, while the POD activity was significantly lower than the CK, and there was no significant effect on the activity of other antioxidant enzymes and content of reductive protective components. The antioxidant enzyme activity and reduced GSH content of the flesh treated with two UV-B radiations increased first and then decreased with time. Compared with CK, the activity of SOD and CAT in the flesh treated with UV-B radiation were significantly decreased. POD activities, flavonoids and reduced GSH contents in the flesh treated by 96 kJ/(m2·d) were significantly lower than the CK, while POD activities treated by 24 kJ/(m2·d) showed no significant difference from CK. Flavonoids and reduced GSH contents was significantly lower than the CK only in the post-ripping stage, while polyphenol contents was almost unaffected. It can be seen that enhanced UV-B radiation [96 kJ/(m2·d)] caused damage to the tree body and flesh, but no damage to the pericarp.
Keywords: enhanced UV-B radiation; mango fruits; injures; anti-oxidation
太陽輻射中的紫外(UV)輻射包含A區(320~ 400 nm)、B區(280~320 nm)和C區(200~280 nm)。其中,UV-C屬于滅生性輻射,多數植物受到UV-C輻射后幾乎立即死亡;UV-A輻射對植物基本無害;UV-B為生物有效輻射,絕大多數植物受到UV-B輻射后會產生脅迫及應激反應[1]。大氣臭氧層衰減和臭氧濃度季節性變化形成“臭氧空洞”,導致到達地球表面太陽輻射中的UV-B輻射增加,此時的UV-B輻射稱為“增強UV-B輻射”(或稱:UV-B輻射增強,enhanced UV-B radiation)。
UV-B輻射會損害生命,其可通過對DNA的直接破壞,使蛋白質和酶失活,造成膜和其他細胞結構的破壞以及產生高活性化學物質(被稱為“自由基”)來影響植物生長發育[2-5]。于妮娜[6]研究表明,增強UV-B輻射顯著改善設施桃果實品質,提高了果皮類胡蘿卜素和花青苷的含量,有利于果實著色。也有研究表明,隨著UV-B輻射的增強,葡萄果實大小及產量降低且葡萄著色指數下降,進而影響葡萄酒的質量[7]。另外,增強UV-B輻射誘導果實中防御酶活性提高進而限制了蘋果采后炭疽病的發展[8]。補增UV-B輻射通過降低香蕉葉片光合速率和氮在光合碳循環組分的分配,限制香蕉葉片光合速率和降低其生物產量[9]。可見,不同強度的增強UV-B輻射對不同的果樹樹種產生了不同的影響。
根據相關預測,今后70年內全球大氣臭氧濃度將減少2%~10%,到達地球表面的UV-B輻射將增加4%~20%[2]。可見,增強UV-B輻射將會是未來的農業逆境,許多作物會陸續遭遇增強UV-B輻射的光脅迫。鑒于上述樹種和增強UV-B輻射強度差異表現不同的生物學效應,有必要就特定的作物開展增強UV-B輻射對其影響的生物學效應研究。
杧果(Mangifera indica L.)是熱帶特色明顯且已產業化的果樹,在海南產區主要分布于長夏無冬、光照充足、紫外輻射強烈的西部、西南部、南部和東南部,杧果能夠很好地適應這種強紫外輻射環境。目前,前人探討了杧果幼苗離體葉片和成年樹葉片在增強UV-B輻射脅迫下的應激特點,并且在成年樹上的結果表明,24 kJ/(m2·d)的增強UV-B輻射處理未引起‘臺農一號杧果和‘金煌杧果樹體損傷,而72 kJ/(m2·d)及其以上處理則引起了‘臺農一號杧果和‘金煌杧果樹體損傷[10-12]。前人曾報道,低劑量增強UV-B輻射處理對葡萄樹體光合作用具有促進效應[13]。同時為確保增強UV-B輻射處理果實后與對照之間在損傷和抗氧化方面具有差異,本研究設定了低劑量24 kJ/(m2·d)和更高劑量的96 kJ/(m2·d) 2個處理,基于此研究‘臺農一號杧果果實在增強UV-B輻射處理下的損傷和抗氧化特點,其為探討增強UV-B輻射影響果實發育和抗氧化機理提供參考,也為未來制定芒果抗增強UV-B輻射栽培技術奠定理論基礎。
1? 材料與方法
1.1? 材料
試驗地點位于海南省陵水縣英州鎮軍田村杧果園,該地區屬熱帶季風海洋氣候,日照充足,雨量充沛,年日照時數2261.6 h,年平均降水量1717.9 mm,年平均氣溫25.4?℃,園土為磚紅性沙壤土。選擇10年生且生長健壯、長勢均勻、無不良表現的成年‘臺農一號杧果(Mangifera indica Linn cv. Tainung No.1)樹9株作為試驗材料。12月到翌年2月為開花坐果期,3—4月為果實迅速膨大期,5月上中旬為果實成熟期(采收期)。
1.2? 方法
1.2.1? 試驗設計? 以自然光為對照(CK,UV-B輻射平均劑量為83.47 kJ/(m2·d)),設置2個UV-B輻射處理:24、96 kJ/(m2·d)。其中96?kJ/(m2·d)相當于環境中UV-B輻射增強了15%。自2018年3月21日(生理落果已結束)至5月8日一直在日照時間內進行人工模擬增強UV-B輻射處理。單株小區,重復3次。在試驗園搭建鋁合金棚架,在UV-B輻射處理的杧果樹樹頂部懸掛UV-B燈架,處理樣樹的燈架內懸掛紫外燈(購于北京電光源研究所,功率為40 W,波長峰值為313 nm,輻射劑量為24 kJ/(m2·d))作為UV-B輻射光源,光源燈外包被0.08 mm醋酸纖維素膜過濾UV-C;燈管離樹頂距離保持30 cm(在果實生長發育期間,新梢抽生慢,并且抹芽控梢,此高度能維持不變)。24?kJ/(m2·d)處理的紫外燈(1根燈管)位于樹頂正中位置,96 kJ/(m2·d)的紫外燈在樹頂正中位置十字交叉4根燈管。晴天進行人工模擬增強UV-B輻射處理,處理時間自日出至日落,陰天、雨天、陣雨和短暫陰天時段則都停止處理(關閉光源燈電源)。
1.2.2? 取樣及樣品處理? 自2018年3月21日開始處理時第1次取樣,此后分別于4月8日、4月22日和5月8日在田間取果樣,隨機采取樹冠中部外圍無病蟲害的果實,每次10個。在5月8日果實采收時,調查單株產量,在各單株上隨機采果30個用于后熟,在實驗室后熟1周達完全成熟,期間于5月11日和15日各取樣1次,各小區隨機取果樣10個。用水果刀快速分離果皮和果肉,切碎,混勻,液氮速凍后置于?80?℃超低溫冰箱保存。
1.2.3? 測定指標與方法? 采用蒽酮比色法測定果肉可溶性糖含量[14]。采用酸堿滴定法測定果肉可滴定酸含量[14]。同一個果實果肉可溶性糖含量與可滴定酸含量的比值為該果實糖酸比。采用硫代巴比妥酸比色法測定果皮和果肉丙二醛(MDA)含量[15]。采用氮藍四唑光還原法測定果皮和果肉超氧化物歧化酶(SOD)活性[16]。采用改良的愈創木酚過氧化氫法測定果皮和果肉過氧化物酶(POD)活性[16]。采用紫外吸收法測定果皮和果肉過氧化氫酶(CAT)活性[16]。采用Folin酚法測定果皮和果肉多酚總含量[17]。采用1%鹽酸甲醇提取法測定果皮和果肉類黃酮含量[18]。采用巰基試劑DTNB測定果皮和果肉還原型谷胱甘肽含量[15]。采用2,6二氯酚靛酚染色滴定法測定果皮和果肉維生素C含量[14]。采用高效液相色譜法測定果皮和果肉中杧果苷等呫噸酮類化合物含量[19]。
1.3? 數據處理
采用SAS 9.1.3軟件進行數據統計分析,采用ANOVA過程作方差分析,采用LSD法作多重比較分析。
2? 結果與分析
2.1? 不同UV-B輻射對樹體產量和果實品質的影響
2.1.1? 對株產和單果重的影響? 不同UV-B輻射處理對株產和單果重的影響如圖1所示。對杧果的株產而言,24 kJ/(m2·d)處理顯著高于96?kJ/(m2·d),增幅為44.87%,但與CK差異不顯著。而96?kJ/(m2·d)處理顯著低于CK,降幅為33.89%。就杧果單果重而言,24 kJ/(m2·d)處理低于CK但差異不顯著,96 kJ/(m2·d)處理顯著低于CK,降幅為9.35%。以上結果說明,96 kJ/(m2·d)處理能引起杧果果實變小,并通過抑制果實膨大而引起樹體減產。
不同小寫字母表示差異顯著(P<0.05)。
Different lowercase letters indicate significant difference (P<0.05).
2.1.2? 對果實營養風味品質的影響? 不同UV-B輻射處理對果實營養風味品質的影響如圖2所示。2個UV-B輻射處理和CK的果肉可溶性糖含量均隨時間呈上升趨勢。4月22日前,2個處理和CK間無顯著差異。此后,2個處理均顯著低于CK。2個處理和CK的果肉可滴定酸含量均隨時間呈持續下降趨勢。96 kJ/(m2·d)處理在整個處理期均顯著高于CK和24 kJ/(m2·d)處理,但24?kJ/ (m2·d)和CK間無顯著差異。2個處理和CK的果肉糖酸比均在總體上隨時間呈上升趨勢。在4月22日之前,2個處理與CK無顯著性差異;自5月8日采后到熟至5月15日,96 kJ/(m2·d)處理顯著低于CK和24 kJ/(m2·d)處理。24 kJ/(m2·d)處理僅在5月15日顯著低于CK,其余處理時間均與CK無顯著差異。以上結果說明,24 kJ/(m2·d)處理對果實營養風味品質無顯著影響,但96?kJ/ (m2·d)處理通過提高果肉可滴定酸含量而導致綜合風味品質變劣。
2.2? 不同UV-B輻射對杧果果皮和果肉活性氧的損傷
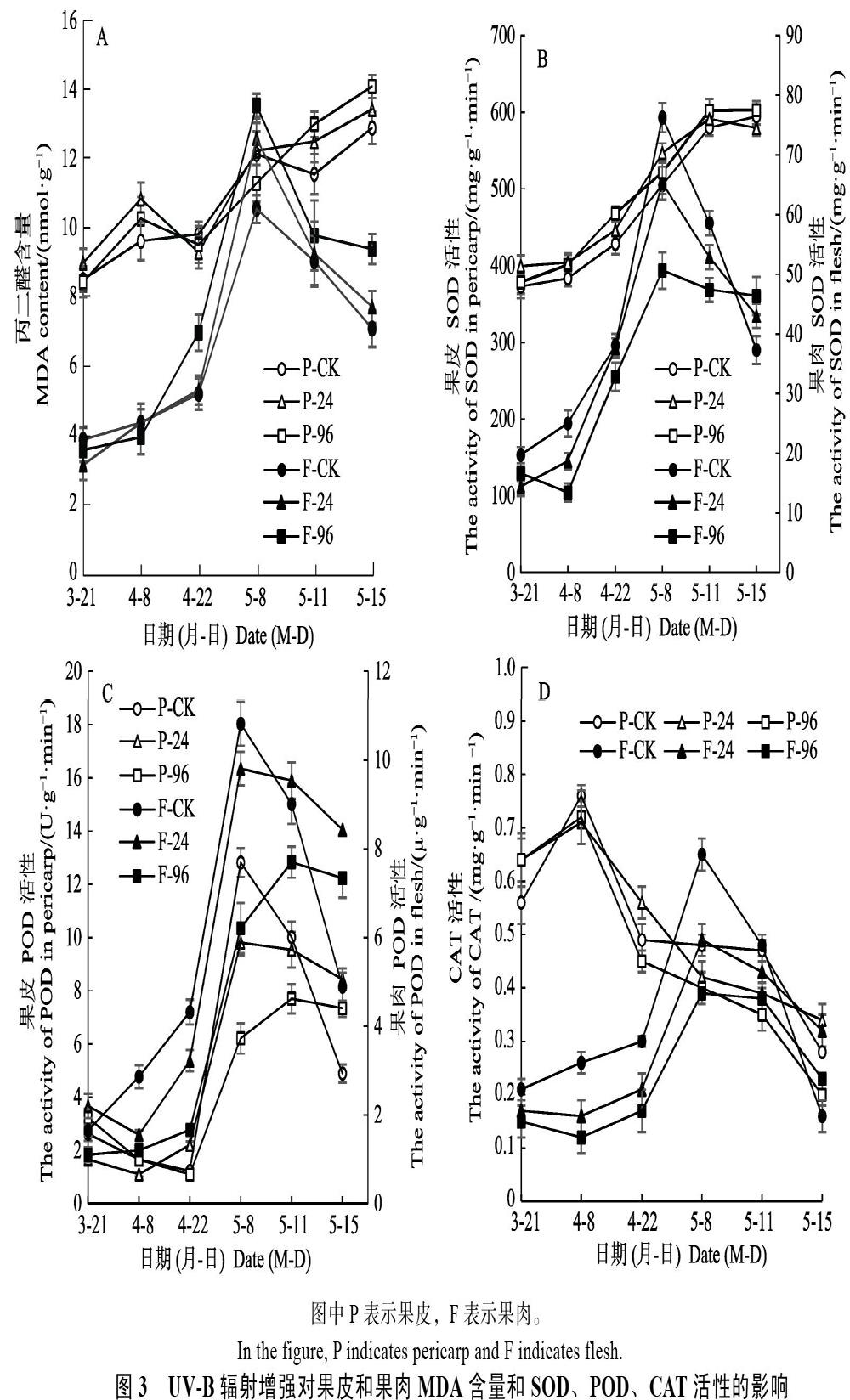
不同UV-B輻射對杧果果皮和果肉MDA含量的影響如圖3A所示。就果皮的MDA而言,2個UV-B輻射處理和CK總體上均隨時間呈上升趨勢。在整個處理期間2個處理和CK間均無顯著性差異。就果肉的MDA而言,2個處理和CK均在田間生長發育期間呈先上升,在采果后呈下降趨勢。在4月8日前,其MDA含量在2個處理與CK間無顯著差異。此后至5月8日,96 kJ/(m2·d)處理的MDA含量顯著高于CK,24 kJ/(m2·d)處理除5月8日顯著高于CK外,則一直與CK無顯著差異。可見,2個處理均未導致果皮出現明顯的細胞膜酯過氧化損傷,24 kJ/(m2·d)處理也未導致果肉明顯的細胞膜脂過氧化損傷,但96 kJ/(m2·d)處理會導致果肉明顯的細胞膜酯過氧化損傷。在果實后熟期間因其已脫離人工模擬增強UV-B輻射環境,可能是果肉細胞膜酯過氧化損傷減弱而引起此期間MDA含量下降。
2.3? 不同UV-B輻射對果皮和果肉抗氧化酶系活性的影響
2.3.1? 不同UV-B輻射對SOD活性的影響? 不同UV-B輻射處理對杧果果皮和果肉SOD活性的影響如圖3B所示。就果皮的SOD活性而言,2個處理和CK在整個處理期間均隨時間呈上升趨勢,但2個處理和對照均無顯著性差異。2個處理和CK的果肉SOD活性均在采果前呈上升、在采果后呈下降趨勢。就果肉SOD活性而言,在4月22日之前,CK與2個處理間均無顯著差異,但5月8日至5月11日,96?kJ/(m2·d)處理顯著低于CK,24 kJ/(m2·d)處理始終與CK無顯著差異。可見,2個處理均未導致果皮明顯的活性氧損傷,24 kJ/(m2·d)處理也未導致果肉明顯的活性氧損傷,但96 kJ/(m2·d)處理導致后熟期的果肉出現明顯的活性氧損傷,即清除O2?活性氧自由基的能力下降,造成O2?的積累而導致損傷。
2.3.2? 不同UV-B輻射對POD活性的影響? 不同UV-B輻射對杧果果皮和果肉POD活性的影響如圖3C所示。2個處理和CK的果皮POD活性均在采果前呈上升、在采果后呈下降的趨勢。就果皮POD活性而言,在4月22日之前,處理與CK間無顯著差異,但5月8日至5月11日,96 kJ/(m2·d)處理顯著低于CK和24?kJ/(m2·d)處理,而24 kJ/(m2·d)處理僅在5月8日顯著低于CK。2個處理和CK的果肉POD活性隨時間也呈先上升后下降的趨勢。自4月8日至5月8日,CK果肉POD活性顯著高于96 kJ/ (m2·d)處理,而與24 kJ/(m2·d)處理無顯著差異。24 kJ/(m2·d)處理的果肉POD活性自4月22日至5月11日顯著高于96 kJ/(m2·d)處理。96 kJ/(m2·d)在處理開始后果肉POD活性和在后熟期前半段的果皮POD活性均比CK低,說明96 kJ/(m2·d)處理導致果肉在田間清除O22?活性氧自由基的能力下降,造成O22?的積累而損傷果肉。
2.3.3? 不同UV-B輻射對CAT活性的影響? 不同UV-B輻射對杧果果皮和果肉CAT活性的影響如圖3D所示。2個處理和CK的果皮CAT活性總體上隨時間呈下降趨勢;96?kJ/(m2·d)果皮CAT活性僅在5月11日顯著低于CK,其余時間3者間均無顯著差異。2個處理和CK的果肉CAT活性均在采果前呈上升、在采果后呈下降的趨勢。就果肉CAT活性而言,自4月8日至5月11日,96 kJ/(m2·d) 顯著低于CK,24 kJ/(m2·d)果肉CAT活性自4月8日至5月8日顯著低于CK,且除了5月8日顯著低于96?kJ/(m2·d)處理外,其余時間2者間均無顯著差異。可見,除96 kJ/(m2·d)處理在4月8日和5月11日抑制果皮清除H2O2活性氧自由基的能力外,2個處理在其余更長的時間里基本對果皮清除H2O2活性氧自由基的能力無明顯影響,但2個處理會導致果肉清除H2O2活性氧自由基的能力下降。
2.4? 不同UV-B輻射對果皮和果肉還原性保護成分的影響
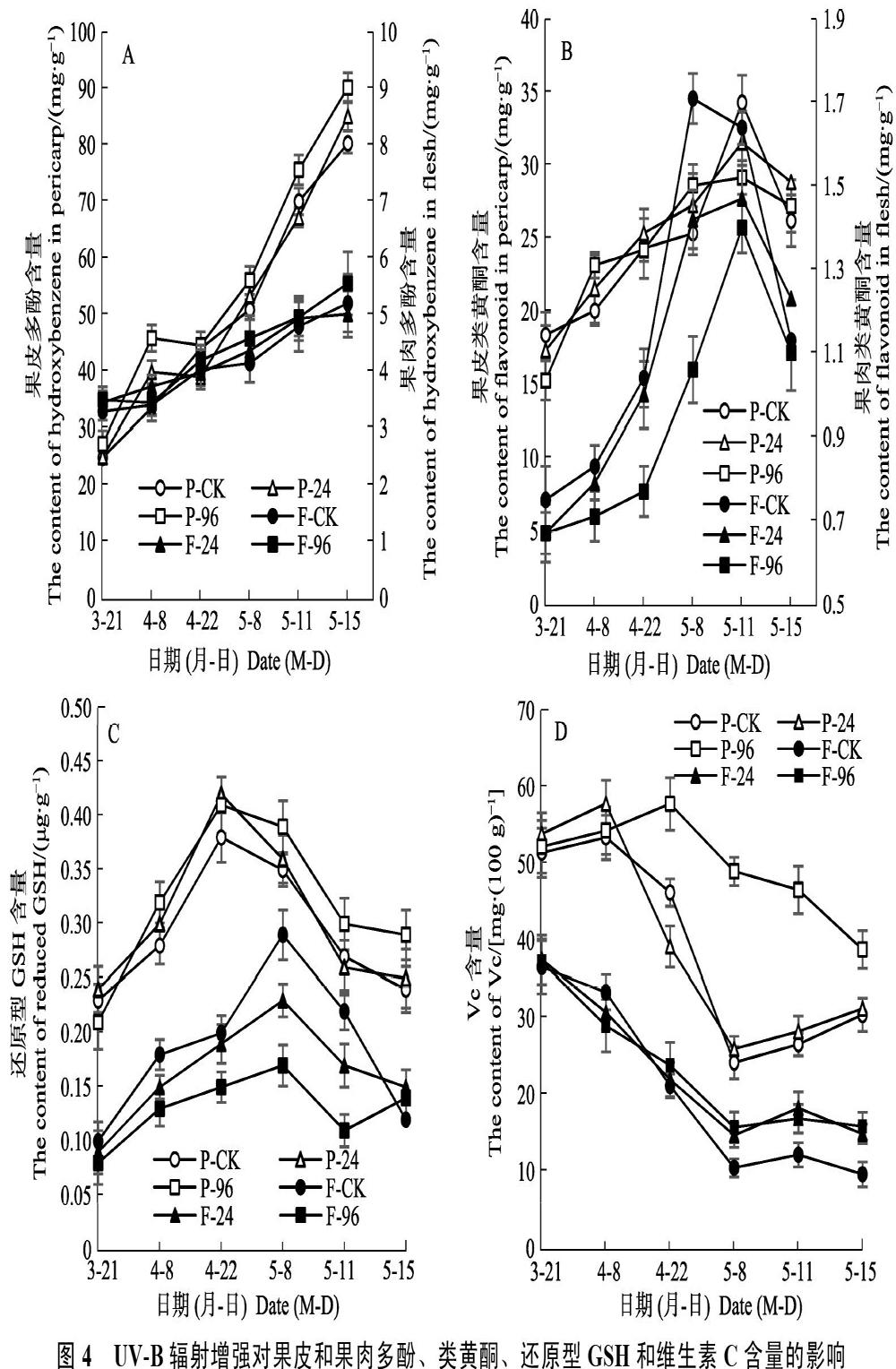
2.4.1? 不同UV-B輻射對多酚的影響? 不同UV-B輻射處理對杧果果皮和果肉多酚含量的影響如圖4A所示。2個處理和CK的果皮和果肉的多酚含量在總體上隨時間均呈上升的趨勢。在整個試驗處理期內3者間均無顯著差異。可見,2個處理在大多數時間里對果皮和果肉中多酚含量無顯著影響。
2.4.2? 不同UV-B輻射對類黃酮的影響? 不同UV-B輻射處理對杧果果皮和果肉類黃酮含量的影響如圖4B所示。2個處理和CK果皮類黃酮含量均隨時間呈不斷上升趨勢。CK果皮類黃酮含量僅在5月11日顯著高于2個UV-B輻射處理,3者在其余時間里相互間均無顯著差異。2個處理和CK的果肉類黃酮含量均在采果前呈上升、在采果后呈下降趨勢。就果肉類黃酮含量而言,在4月22日之前,UV-B輻射處理與CK間無顯著差異,但自4月22日至5月11日,96 kJ/(m2·d)處理顯著低于CK,24?kJ/(m2·d)處理僅在5月8日和5月11日顯著低于CK,其余處理時間3者間無顯著差異。可見,2個UV-B輻射處理未能通過增加果皮類黃酮含量來減輕損傷,而96 kJ/(m2·d)處理可通過消耗果肉中的類黃酮來盡可能清除活性氧自由基和減輕損傷。
2.4.3? 不同UV-B輻射對還原型GSH的影響? 不同UV-B輻射處理對杧果果皮和果肉還原型GSH含量的影響如圖4C所示。2個處理和CK的果皮還原型GSH含量隨時間呈先上升后下降趨勢;96 kJ/(m2·d)處理僅在5月15日顯著高于CK,其余處理時間3者之間無顯著差異。2個處理和CK的果肉還原型GSH含量均在采果前呈上升、在采果后呈下降趨勢。自4月8日至5月11日,96 kJ/(m2·d)處理顯著低于CK,24 kJ/(m2·d)處理僅在采后后熟期顯著低于CK。可見,2個UV-B輻射處理未能通過提高果皮還原型GSH含量來減輕損傷,但其可通過消耗果肉中的還原型GSH來盡可能清除活性氧自由基和減輕損傷。
2.4.4? 不同UV-B輻射對維生素C的影響? 不同UV-B輻射處理對杧果果皮和果肉維生素C含量
的影響如圖4D所示。2個處理和CK的果皮維生素C含量隨時間呈不斷下降趨勢。96 kJ/(m2·d)果皮維生素C含量除4月8日外,均顯著高于CK和24 kJ/(m2·d)處理。24 kJ/(m2·d)果皮維生素C含量僅在4月8日顯著高于CK,其余處理時間與CK無顯著差異。2個處理和CK的果肉維生素C含量隨時間呈不斷下降趨勢。4月22日前,3者之間均無顯著性差異,4月22日后,2個UV-B輻射處理果肉維生素C含量均顯著高于CK,但2個處理之間無顯著差異。可見,96 kJ/(m2·d)處理可通過增加果皮和果肉維生素C積累來竭力抵御活性氧積累而導致的細胞損傷。
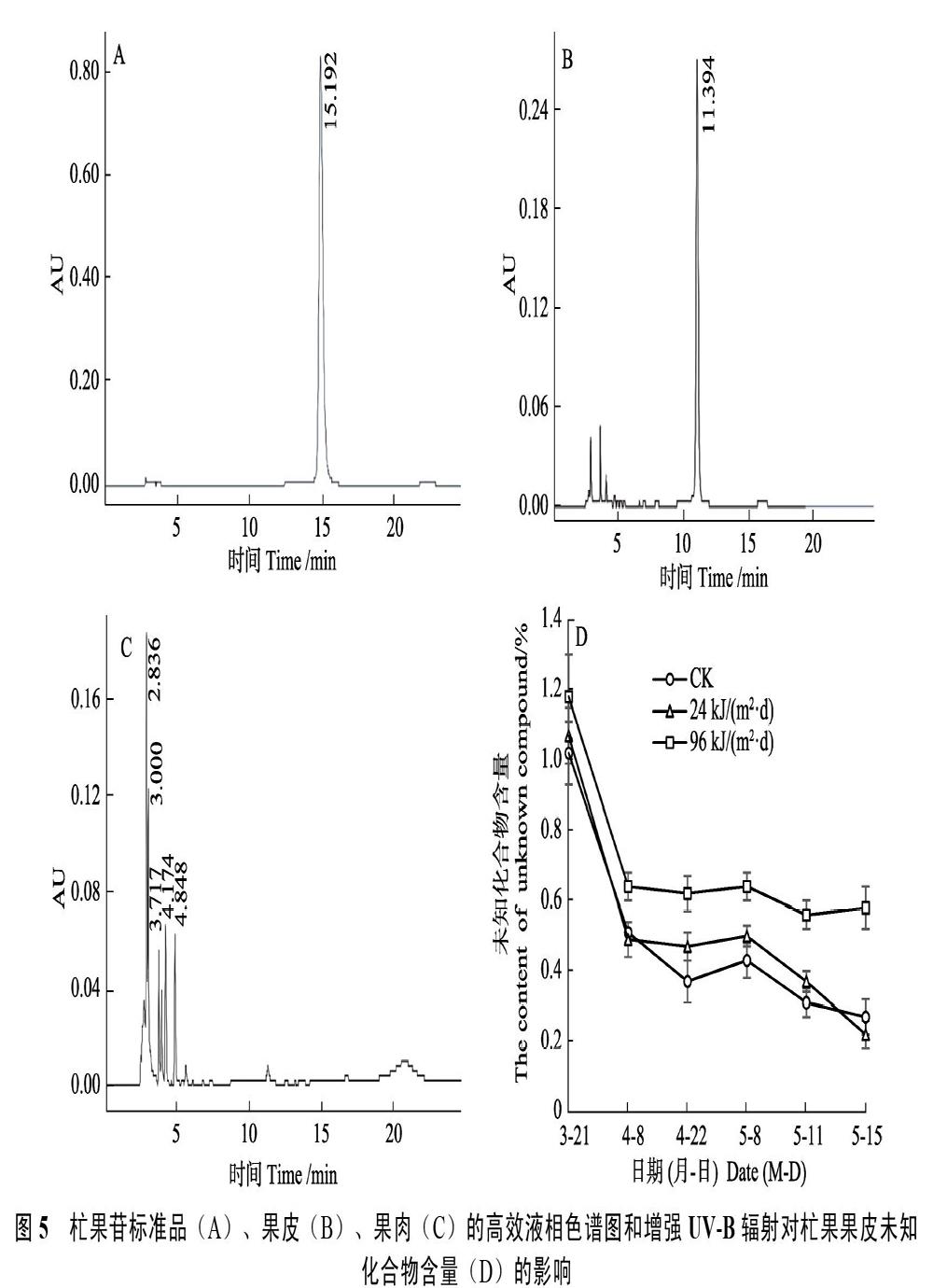
2.4.5? 未知化合物? 杧果苷標準品的液相色譜圖如圖5A所示。在(15.467±0.014)?min時間范圍內出現最大峰面積。杧果果皮的液相色譜圖如圖5B所示,所有樣品均未在標準品的出峰時間范圍內出峰而均在(11.397±0.004)?min時間范圍內出現最大峰面積。由于該峰不屬于杧果苷的檢測峰,說明果皮中可能存在某未知化合物。杧果果肉的液相色譜圖如圖5C所示,在5~25 min范圍內未出現任何檢測高峰,說明果肉不含杧果苷和上述果皮中存在的未知化合物。
不同UV-B輻射處理對杧果果皮上述未知化合物含量的影響如圖5 D所示。2個處理和CK未知化合物含量隨時間均呈不同程度的下降趨勢。在4月22日至5月15日,96 kJ/(m2·d)未知化合物含量顯著高于CK和24 kJ/(m2·d),而24 kJ/(m2·d)與CK間無顯著差異。推測96 kJ/(m2·d)可能通過增加果皮積累該未知化合物來盡可能清除活性氧自由基或吸收UV-B輻射而耗散脅迫,進而減輕損傷,來提高果實抵御高強度UV-B輻射逆境的能力。
3? 討論
Yin等[20]研究表明,增強UV-B輻射下的玉米籽粒產量連續3 a分別下降了14.6%、18.2%和
16.8%。棉花單株鈴數與單鈴重均隨UV-B輻射的增強而降低,導致籽棉產量顯著降低,且增強UV-B輻射越強,抑制作用越明顯[21]。方芳等[22]
研究表明,高強度增強UV-B輻射處理會導致采后葡萄果實的外在品質及內在品質急劇下降。鮮見增強UV-B輻射對多年生果樹產量的研究,連續3 a在‘臺農一號和‘金煌杧果上的研究發現,高強度增強UV-B輻射能通過抑制果實膨大而導致減產和能引起營養風味品質變劣[23-24]。本研究表明,96 kJ/(m2·d)的增強UV-B輻射可引起杧果成年樹減產、果實大小和風味品質變劣,均取得了與前人的研究結果一致的結果。
人工模擬增強UV-B輻射照射杧果果實時,一部分光被果皮反射掉,一部分光被果皮吸收,還有一部分光穿透果皮后被果肉吸收。本研究發現,果皮吸收的部分未對果皮細胞膜造成損傷,果肉吸收的部分則造成果肉細胞膜損傷。
杧果苷(1,3,6,7-四羥基呫噸酮-C2-?-D-葡萄糖苷)是一種主要從杧果樹體中分離出來的生物活性成分,其具有吸收UV-B輻射的能力和強大的抗氧化活性[25-26]。對從川西獐芽菜分離得到的24個呫噸酮類化合物并進行抗氧化活性篩選,發現呫噸酮類化合物有較好的抗氧化能力[27]。在杧果苷等呫噸酮類化合物檢測體系中,能同時分離檢測到杧果苷、新杧果苷和異杧果苷等呫噸酮類化合物[28]。基于此,本研究發現,果皮樣品出現不屬于杧果苷的檢測峰,其說明這種未知成分很可能屬于呫噸酮類化合物。呫噸酮類化合物具備吸收UV-B輻射的能力和抗氧化性,高強度增強UV-B輻射處理通過顯著提高其含量來耗散高強度增強UV-B輻射脅迫和清除活性氧自由基,進而保護果皮免遭增強UV-B損傷。至于本研究所檢測到的可能為未知呫噸酮類化合物,其分子結構尚不清楚,有必要進一步開展該單體化合物的分離純化和結構鑒定研究。
本研究結果發現,高強度增強UV-B輻射處理刺激果皮積累維生素C,可能藉此增強清除活性氧自由基的能力,這與前人用外源維生素C處理豌豆葉片而減輕豌豆葉片活性氧損傷的研究結果相印證[29],進而使果皮免遭高強度增強UV-B輻射損傷[30]。
高強度增強UV-B輻射一方面促使果肉和果皮通過增加維生素C的積累來提高果實清除活性氧自由基的能力[29-30]。另一方面,通過抑制POD、CAT酶活性和還原型GSH含量來削弱清除活性氧自由基的能力,進而引起活性氧自由基積累。同時還原型GSH也能通過吸收UV-B輻射而耗散高強度增強UV-B輻射脅迫[31-32],高強度增強UV-B輻射處理可能通過抑制其積累而失去耗散脅迫的能力;前者增強的抗活性氧損傷能力可能在量上低于后者引起的減弱,進而果肉遭到活性氧損傷。總之,果肉的增強UV-B輻射損傷歸咎于果肉POD、CAT活性減弱和還原型GSH積累的減少。
4? 結論
增強UV-B輻射處理[96 kJ/(m2·d)]使‘臺農一號杧果單果重下降、樹體減產以及風味品質變劣,即對樹體產生了損傷。增強UV-B輻射處理對杧果果皮沒有造成損傷,可能歸因于未知呫噸酮類化合物和維生素C的積累,兩者清除了活性氧自由基和前者吸收一部分增強UV-B輻射而耗散脅迫。增強UV-B輻射通過增加果肉維生素C的積累來提高果肉清除活性氧自由基的能力,但其同時抑制了果肉POD、CAT的活性和還原型GSH含量而削弱了清除活性氧自由基的能力,可能后者削弱的活性氧自由基清除能力大于前者增強的部分,進而引起損傷。增強UV-B輻射處理可能誘導果皮產生一種未知呫噸酮類的化合物,其也許具備極強的抗氧化活性而可能開發為醫藥、獸藥、飼料、化妝品和保健品等,為杧果果實的綜合開發利用開創了新途徑。采用合適的逆境脅迫處理促使果肉POD、CAT活性和還原型GSH含量等升高,可能會提高‘臺農一號杧果果肉對增強UV-B輻射脅迫的抗性,這對防止‘臺農一號杧果果肉的增強UV-B輻射損傷具有一定參考意義。
參考文獻
Mckenzie R L, Aucamp P J, Bais A F, et al. Ozone depletion and climate change: impacts on UV radiation[J]. Photochemical and Photobiological Sciences, 2011, 10(2): 182-198.
Caldwell M M. Solar UV irradiation and the growth and development of higher plants[J]. Photophysiology, 1971, 4: 131-177.
Rozema J, Lenssen G M, Staaij W M V D J, et al. Effects of UV-B radiation on terrestrial plants and ecosystems: Interaction with CO2 enrichment[J]. Plant Ecology, 1997, 128(1-2): 183-191.
Jansen M A K. Higher plants and UV-B radiation: balancing damage, repair and acclimation[J]. Trends in Plant Science, 1998, 3(4): 131-135.
Hollósy F. Effects of ultraviolet radiation on plant cells[J]. Micron, 2002, 33(2): 179-197.
于妮娜. UV-B增強對設施桃碳氮營養分配和果實品質的影響[D]. 泰安: 山東農業大學, 2012.
孫? 瑩, 張振文, 岳泰新, 等. 不同強度紫外線(UV)-B輻射對葡萄品質影響[J]. 中國釀造, 2009(10): 15-18.
吳芳芳, 鄭有飛, 萬長健, 等. UV-B輻射增強對蘋果采后炭疽病發病情況和抗病相關酶活性的影響[J]. 生態環境, 2008, 17(3): 962-965.
孫谷疇, 趙? 平, 曾小平, 等. 補增UV-B輻射對香蕉葉片光合作用和葉氮在光合碳循環組分中分配的影響[J]. 植物學通報, 2000, 17(5): 450-456.
劉? 鵬, 周開兵, 王? 爽, 等. 芒果不同葉齡葉片在增強UV-B輻射處理下的損傷和保護響應[J]. 熱帶生物學報, 2010, 1(4): 350-356.
袁孟玲, 岳? 堃, 王? 紅, 等. ‘臺農一號 芒果成年樹在增強UV-B輻射處理下的損傷及其抗氧化響應[J]. 熱帶作物學報, 2018, 39(9): 1724-1731.
周開兵, 李世軍, 袁孟玲, 等. 杧果成年樹在增強UV-B輻射處理下的損傷與抗氧化響應[J]. 園藝學報, 2019, 46(7): 1279-1289.
周新明, 張振文, 惠竹梅, 等. UV-B輻射增強對葡萄光合作用日變化的影響[J]. 農業工程學報, 2009, 25(3): 209-212.
李合生. 植物生理生化實驗原理和技術[M]. 北京: 高等教育出版社, 2000: 134-138.
陳建勛, 王曉峰. 植物生理實驗學實驗指導[M]. 廣州: 華南理工大學出版社, 2006.
張蜀秋. 植物生理學實驗技術教程[M]. 北京: 科學出版社, 2011: 191-194.
Li X C, Hu Q P, Jiang S X, et al. Flos chrysanthemi indici protects against hydroxyl-induced damages to DNA and MSCs via antioxidant mechanism: A chemistry study[J]. Journal of Saudi Chemical Society, 2015, 19(4): 454-460.
曹健康, 姜微波, 趙玉梅. 果蔬采后生理生化實驗指導[M]. 北京: 中國輕工業出版社, 2007: 44-46.
馮? 旭, 王勝波, 鄧家剛, 等. 高效液相色譜法同時測定芒果葉中芒果苷與高芒果苷的含量[J]. 中成藥, 2008, 30(10): 1504-1506.
Yin L N, Wang S W. Modulated increased UV-B radiation affects crop growth and grain yield and quality of maize in the field[J]. Photosynthetica, 2012, 50(4): 595-601.
祁? 虹, 段留生, 王樹林, 等. 全生育期UV-B輻射增強對棉花生長及光合作用的影響[J]. 中國生態農業學報, 2017, 25(5): 708-719.
方? 芳, 王鳳忠. UV-B照射對采后葡萄果實品質和黃酮醇積累的影響[J]. 食品工業科技, 2017, 38(19): 272-277, 283.
周開兵, 李世軍, 袁孟玲. 增強UV-B輻射對芒果株產和果實品質及光合作用的影響[J]. 熱帶作物學報, 2018, 39(6): 1102-1107.
袁孟玲, 岳? 堃, 王? 紅, 等. 增強UV-B輻射對芒果成年樹光合作用及其產量與常規品質的影響[J]. 南方農業學報, 2018, 49(5): 930-937.
Kulkarni V M, Rathod V K. Mapping of an ultrasonic bath for ultrasound assisted extraction of mangiferin from Mangifera indica leaves[J]. Ultrasonics Sonochemistry, 2014, 21(2): 606-611.
Du S Y, Liu HR, Lei T T, et al. Mangiferin: An effective therapeutic agent against several disorders (Review)[J]. Molecular Medicine Reports, 2018, 18(6): 4775-4786.
羅翠婷, 毛雙雙, 陳河如, 等. 川西獐芽菜化學成分研究[J]. 中草藥, 2013, 44(8): 942-946.
宋澤璧, 吳? 瑩, 高? 慧. HPLC法測定鹽炙前后知母中新芒果苷、芒果苷和異芒果苷[J]. 現代藥物與臨床, 2015, 30(2): 145-148.
Dobrikova A G, Apostolova E L. Damage and protection of the photosynthetic apparatus from UV-B radiation. II. Effect of quercetin at different pH[J]. Journal of Plant Physiology, 2015, 184: 98-105.
劉景玲. UV-B輻射和干旱對丹參生物量及酚酸類成分含量的影響[D]. 楊凌: 西北農林科技大學, 2014.
王? 靜, 馮娜娜, 王? 丹, 等. UV-B輻射對紫花苜蓿幼苗品質和抗氧化性的影響[J]. 食品科技, 2016, 41(6): 56- 60.
劉景玲, 齊志鴻, 郝文芳, 等. UV-B輻射和干旱對丹參生長和葉片中酚酸類成分的影響[J]. 生態學報, 2015, 35(14) : 4642-4650.
