Bacillus siamensis和Bacillus subtilis中2個β-D-葡萄糖苷酶編碼基因克隆、表達及酶學研究
2020-03-23 06:07:38陳永敢谷風林蔡瑩瑩徐飛朱科學
熱帶作物學報 2020年2期
陳永敢 谷風林 蔡瑩瑩 徐飛 朱科學

摘? 要:采用RACE技術擴增2株Bacillus屬菌株XY18、XY20的β-D-葡萄糖苷酶編碼基因bgl全長,構建重組載體pET28a(+)/bgl、pET28b(+)/bgl,導入E. coli BL21誘導表達,采用Ni親和層析純化蛋白。經酶學性質測定發現:2個蛋白均為弱酸性蛋白,溫度為40?℃時酶活力達到最大,具有一定耐熱性。以上結果表明,為優化2株Bacillus屬菌株參與香草蘭發酵工藝,可提供適當弱酸性條件、適宜溫度。
關鍵詞:Bacillus;β-D-葡萄糖苷酶;基因克隆
中圖分類號:Q939.124??? 文獻標識碼:A
Abstract: The full length of bgl of Bacillus sp. XY18 and Bacillus sp. XY20 was amplified by RACE. The recombinant vectors pET28a (+)/bgl and pET28b (+)/bgl were constructed, then transferred into E. coli BL21 for induction expression, and the protein was purified by Ni affinity chromatography. The enzymatic properties of the two proteins were determined. The two proteins were weak acidic proteins, and the enzyme activity reached the maximum at 40 ℃ with certain heat resistance, suggesting that appropriate weak acid condition and temperature could be provided to optimize the vanilla curing process.
Keywords: Bacillus; β-D-glucosidase; gene cloning
β-D-葡萄糖苷酶(β-D-glucosidase, BGL)屬纖維素水解酶系重要組成酶,可水解芳基或烴基與糖基間糖苷鍵生成葡萄糖與苷元[1]。大部分β-D-葡萄糖苷酶對底物糖基結構專一性較差,能水解C-S鍵、C-N鍵、C-F鍵等多種糖苷鍵。此外,有些對糖基C4和C2結構也不專一,能同時水解β-葡萄糖苷鍵和β-半乳糖苷糖。甚至,有些β-D-葡萄糖苷酶對C6位專一性也不高,也能水解木糖。因此,β-D-葡萄糖苷酶水解酶能適應多種底物[2]。
Bacillus屬菌株分布廣泛,菌落表面粗糙不透明,可利用蛋白質、多糖及淀粉,能分解色氨酸形成吲哚,在分子遺傳操作中應用廣泛。至今已從土壤中分離到B. subtilis、B. licheniformis、B. pumilus等多個種,并克隆及表達β-D-葡萄糖苷酶編碼基因[3-5]。研究表明,香草蘭發酵過程中Bacillus屬菌株參與風味物質形成,水解香蘭素葡萄糖苷的Bacillus屬菌株β-D-葡萄糖苷酶在何種條件下發生催化,如何能達到最大催化效率,如何篩選高效產酶菌株,以便改進發酵方法提高香蘭素產量。對分離菌株β-D-葡萄糖苷酶進行研究可為解決上述問題提供參考信息。選擇本實驗室分離到的2株菌株XY18和XY20進行β-D-葡萄糖苷酶編碼基因全長克隆,采用E.coli進行表達,研究β-D-葡萄糖苷酶學性質。通過以上工作,了解2株菌株β-D-葡萄糖苷酶酶學特征。
1? 材料與方法
1.1? 材料
菌株Bacillus siamensis XY18、Bacillus subtilis XY20均為中國熱帶農業科學院香料飲料研究所產品加工研究室保存。
1.2? 方法
1.2.1? RACE擴增bgl全長? 采用細菌基因組DNA抽提試劑盒(UNIQ-10,Sangon)提取基因組DNA。基于根據NCBI數據庫基因同源性獲得的菌株XY18及XY20 β-D-葡萄糖苷酶編碼基因部分片段序列,設計引物,采用RA?CE技術擴增獲得全長,擴增所采用的引物見表1。
1.2.2? 質粒重組、融合蛋白表達及純化? UCm-T載體的重組:目的DNA片段和載體連接采用PCR產物T/A pUCm-T克隆試劑盒(Sangon)進行。反應混合物置于0.5 mL Eppendorf管中16?℃連接1 h。將重組pUCm-T載體及pET28a(+)/pET28b(+)進行酶切并連接,構建重組載體。菌株XY18使用pET28a(+),菌株XY20使用pET28b(+)。取1 μL重組的pET28b(+)質粒轉化BL21菌株,42?℃熱擊90 s,冰上靜置2 min后涂平板(30 ?g/mL卡那霉素),37?℃培養過夜。重組的pET28a(+)采用34 ?g/mL氯霉素篩選。小試培養選擇最佳的誘導條件后重組蛋白的大量誘導。超聲破碎菌體后采用鎳瓊脂糖親和層析純化蛋白。
1.2.3? β-D-葡萄糖苷酶酶活的測定? 酶的測定方法:取酶液10 μL,加入pH 6.5緩沖液80 μL,再加入50 mmol/L?pNPG 10?μL,于37?℃保溫10 min。空白對照以100?℃滅活2 min酶液代替。反應后,加入1 mol/L Na2CO3 100 μL顯色,用酶標儀測405?nm處吸光度(OD405)。1個酶活力單位(U)定義為:pH 6.5、37?℃條件下,每分鐘生成1 μmol pNP所需的酶量。
1.2.4? pH、溫度對β-D-葡萄糖苷酶活性及穩定性的影響? pH、溫度對β-D-葡萄糖苷酶活性影響測定方法參照Michlmayr等[6]方法并略有改動。維持37?℃恒定,在pH 4.0~9.0范圍內以1.0幅度變化測定pH對酶活影響。維持pH 6.5恒定,在溫度25~50?℃范圍內以5.0?℃幅度變化測定溫度對酶活影響。pH 6.5條件下,酶液置于30、40、50、60和70?℃處理后再測定酶活力。pH 4.0~9.0范圍內以1.0幅度變化處理酶液后再測定酶活力。
1.3? 數據處理
采用DNAMAN 6.0構建質粒圖譜,數據統計分析采用Excel 2010結合Origin 8.6軟件進行處理。
2? 結果與分析
2.1? bgl基因全長獲取
根據前期研究已獲得的β-D-葡萄糖苷酶編碼基因bgl部分片段的序列信息,設計引物,采用RACE技術進行染色體位移,擴增獲得菌株XY18和XY20 β-D-葡萄糖苷酶編碼基因全長,CDS區分別為1398和1410 bp。采用BLASTn進行比對,結果表明,菌株XY18 bgl序列與B. amyloliquefaciens的β-D-葡萄糖苷酶編碼基因序列相似性為96%,而菌株XY20 bgl序列與B. subtilis的β-D-葡萄糖苷酶編碼基因序列相似性為99%。其反映了獲得的序列為Bacillus屬菌株β-D-葡萄糖苷酶編碼基因。
2.2? 重組載體構建
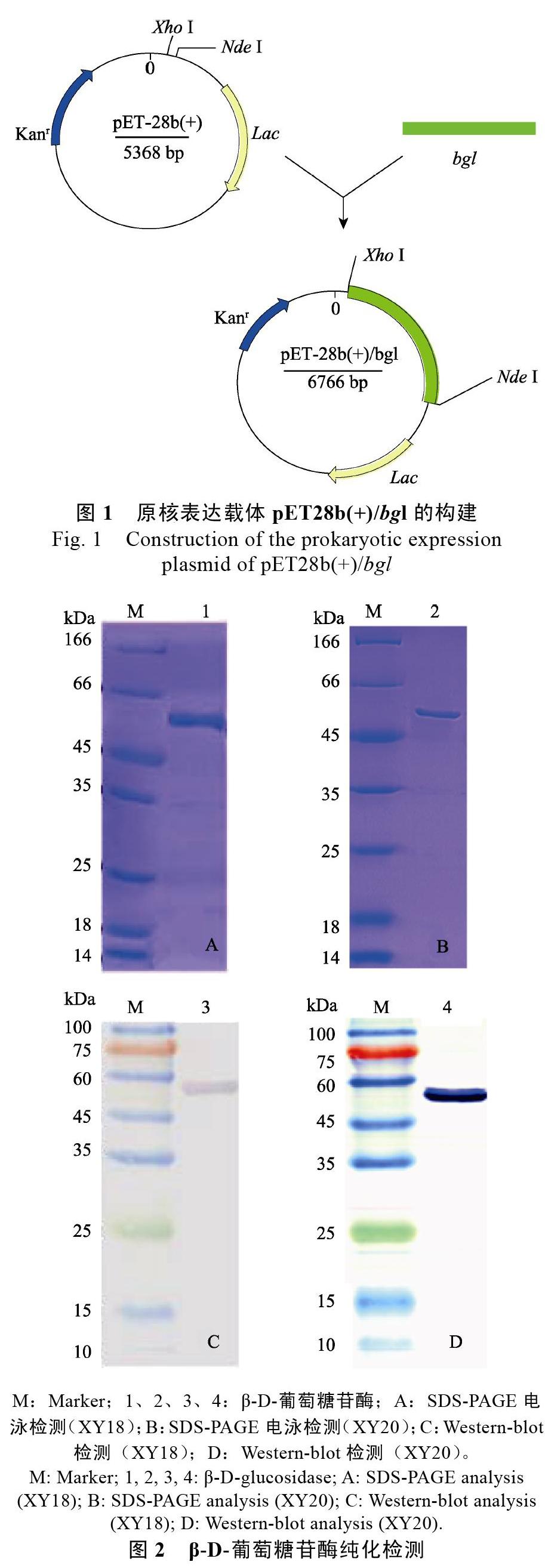
質粒pET28b(+)與含目的片段(XY20)的重組pUCm-T載體經NdeI、XhoI限制性酶切并構建重組pET28b(+)/bgl表達載體,圖1為重組載體pET28b(+)/bgl(以XY20為例)構建框架圖。pET28a(+)與來源于XY18目的片段連接。
2.3? β-D-葡萄糖苷酶純化
分別將含重組蛋白的E. coli BL21在20和37?℃進行誘導表達,并以誘導前菌體沉淀作為對照。融合蛋白(54 kDa)在37 ℃誘導比20 ℃誘導條件下表達量高,因此,選擇37 ℃誘導菌體上清。純化后均獲得SDS-PAGE水平上單一條帶產物,為更進一步確認蛋白純度與表達量,Western blot顯色結果表明,融合蛋白得到大量表達并經純化后獲得單一蛋白產物(圖2)。
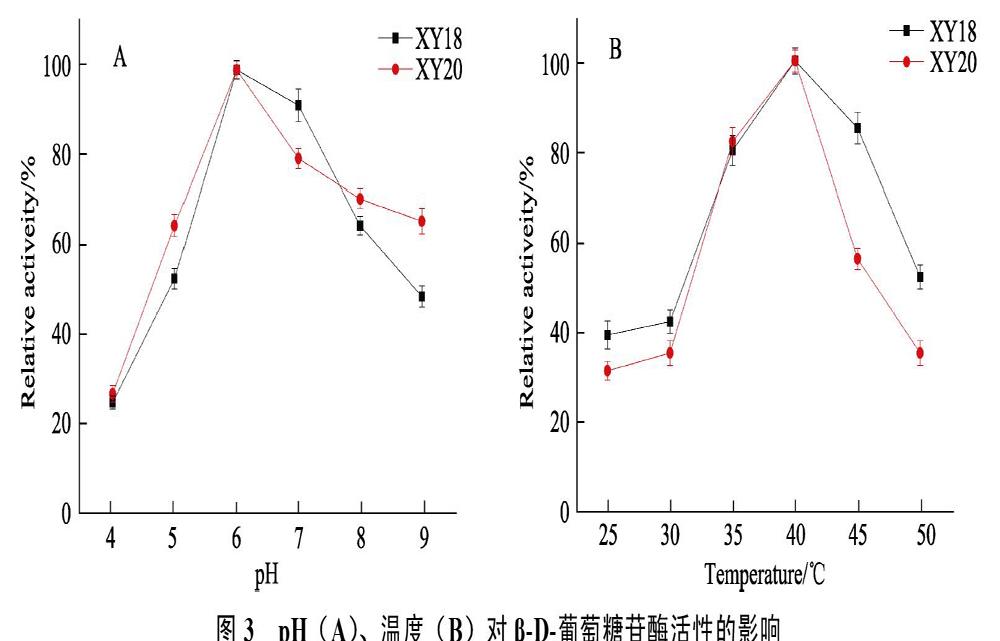
2.4? pH、溫度對β-D-葡萄糖苷酶活性的影響
以最高酶活100%來衡量pH、溫度對β-D-葡萄糖苷酶活力影響。如圖3所示,以pH4.0為起點,隨著pH增加,酶活性增加,在pH6.0時酶活性最大;隨著pH進一步增加,酶活性逐漸降低。以溫度25?℃為起點,隨著溫度增加,酶活性增加,直至40?℃時酶活性最大,然而溫度進一步增加后,酶活性迅速降低。
2.5? pH、溫度對β-D-葡萄糖苷酶穩定性的影響
如圖4所示,在30~70?℃范圍內,溫度越低,酶活性越大,然而2株菌對pH穩定性則略有差異。以pH4.0為起點,隨著pH增加,酶活性增加。菌株XY18在pH為6.0時最穩定,菌株XY20在pH為7.0時最穩定。達到最大酶活后,隨著pH進一步增加,2株菌株酶活性迅速降低。
3? 討論
決定香草蘭品質最重要因素之一為香蘭素含量,而香草蘭發酵目的在于讓豆莢中香蘭素葡萄糖苷接觸β-D-葡萄糖苷酶從而水解產生香蘭素[7]。對香草蘭β-D-葡萄糖苷酶進行純化發現,該酶分子量為201 kDa,由大小相同的4個亞基組成,最適pH為6.5,最適宜溫度為40?℃[8]。內源β-D-葡萄糖苷酶對香蘭素生成的貢獻至今仍存在爭議。Odoux[9]研究發現,根據香草蘭發酵后生成香蘭素含量來衡量,在發酵過程中內源β-D-葡萄糖苷酶僅有效利用其催化活性40%,大部分活性都在其發酵過程中丟失。該課題組進一步研究香蘭素葡萄糖苷、β-D-葡萄糖苷酶活及細胞組織破裂間關系,他們又提出雖然β-D-葡萄糖苷酶酶活在發酵過程中發生失活,但香蘭素葡萄糖苷轉化完全。因此認為,β-D-葡萄糖苷酶并未影響香蘭素葡萄糖苷的水解程度,只影響其水解速率快慢[10]。本課題組研究已證實,內源β-D-葡萄糖苷酶在發酵過程中并未完全參與香蘭素葡萄糖苷水解,在殺青24 h后發生失活,而后續香蘭素葡萄糖苷水解則由外源Bacillus β-D-葡萄糖苷酶來完成,因此,對外源Bacillus菌株β-D-葡萄糖苷酶了解,有助于探討香蘭素葡萄糖苷水解機制及改進發酵工藝。
β-D-葡萄糖苷酶是一種常見水解酶,之前研究已從Bacillus polymyxa、Aureobasidium pullulans、Bacillus circulans subsp. alkalophilus、Thermobifida fusca分離到產β-D-葡萄糖苷酶菌株,這些菌株都具有較強耐熱能力,但由于Bacillus屬菌株在參與發酵過程中產生其他異味,且來源于非食品環境,是否可用于發酵尚有待研究[11-14]。菌株XY18、XY20所產β-D-葡萄糖苷酶顯示一定耐熱能力,說明在發汗、干燥階段即使環境溫度較高,菌株均能存活且起到水解香蘭素葡萄糖苷作用。此外,在接近中性環境條件下,酶活力較高,也為提高催化活性所需環境條件提供參考。
參考文獻
Sternberg D, Vuayakumar P, Reese E T. β-Glucosidase: microbial production and effect on enzymatic hydrolysis of cellulose [J]. Canadian Journal of microbiology, 1977, 23(2): 139-147.
Saburi W, Okuyama M, Kumagai Y, et al. Biochemical properties and substrate recognition mechanism of GH31 α-glucosidase from Bacillus sp. AHU 2001 with broad substrate specificity[J]. Biochimie, 2015, 108: 140-148.
Asha B M, Pathma J, Sakthivel N. Isolation and characterization of a novel thermostable β-glucosidase from Bacillus subtilis SU40[J]. Applied Biochemistry and Microbiology, 2015, 51(1): 21-26.
Choi I S, Wi S G, Jung S R, et al. Characterization and application of recombinant β-glucosidase (BglH) from Bacillus licheniformis KCTC 1918[J]. Journal of wood Science, 2009, 55(5): 329-334.
Li L, Ji Q L, Hao JF, et al. Cloning and expression of β-glucosidase gene from Bacillus pumilus[J]. Applied me-
chanics and materials, 2013, 300-301:1271-1274.
Michlmayr H, Sch?mann C, Barreira Braz da Silva NM, et al. Isolation and basic characterization of a β-glucosidase from a strain of Lactobacillus brevis isolated from a malolactic starter culture[J]. Journal of Applied Microbiology, 2010, 108(2): 550-559.
Dignum M J W, Heijden R V D, Kerler J, et al. Identification of glucosides in green beans of Vanilla planifolia Andrews and kinetics of vanilla β-glucosidase[J]. Food Chemistry, 2004, 85: 199-205.
Odoux E, Chauwin A, Brillouet J M. Purification and characterization of vanilla bean (Vanilla planifolia Andrews) β-glucosidase[J]. Journal of Agricultural and Food Chemistry, 2003, 51(10): 3168-3173.
Odoux E. Changes invanillin and glucovanillin concentrations during the various stages of the process traditionally used for curing Vanilla fragrans beans in Réunion[J]. Fruits, 2000, 55: 119-125.
Odoux E. Glucosylated aroma precursors and glucosidase(s) in vanilla bean (Vanilla planifolia G. Jackson)[J]. Fruits, 2006, 61(3): 171-184.
González-candelas L, Aristoy M C, Polaina J, et al. Cloning and characterization of two genes from Bacillus polymyxa expressing β-glucosidase activity in Escherichia coli[J]. Applied and Environmental Microbiology, 1989, 55(12): 3173-3177.
Paavilainen S, Hellman J, Korpela T. Purification, characterization, gene cloning, and sequencing of a new β-gluco?sidase from Bacillus circulans subsp. alkalophilus[J]. Applied and Environmental Microbiology, 1993, 59(3): 927-932.
Spiridonov N A, Wilson D B. Cloning and biochemical characterization of BglC, a β-glucosidase from the cellulolytic actinomycete Thermobifida fusca[J]. Current Microbiology, 2001, 42(4): 295-301.
董智哲, 谷風林, 徐? 飛, 等. 不同產地香草蘭香氣成分及抗氧化活性比較研究[J]. 中國食品學報, 2015, 15(1): 242-249.
