骨骼源性因子對肌骨“Crosstalk”作用機制的研究進展
2020-03-26 03:56:50宗博藝李世昌孫朋袁子琪
中國骨質疏松雜志 2020年1期
宗博藝 李世昌 孫朋 袁子琪
華東師范大學青少年健康評價與運動干預教育部重點實驗室/體育與健康學院,上海 200241
肌肉衰減癥,又稱“肌少癥”,是指與增齡相關的全身進行性肌肉質量減少,肌強度下降或肌肉功能減退[1];骨質疏松癥是以骨量減少,骨組織微結構破壞,骨脆性增加為主要特征的全身性骨骼疾病[2]。兩種疾病具有相似的病理生理學基礎,針對肌肉、骨骼分子水平上的共同靶點施加干預成為治療肌骨系統疾病(如肌少-骨質疏松癥)的有效方式[3]。研究認為,除了對骨骼產生力學效應外,骨骼肌還可通過分泌白細胞介素-6(interleukin 6,IL-6)和鳶尾素等“肌肉因子”影響骨代謝[4]。然而迄今為止,骨骼作用于肌肉的生理途徑仍是一個全新的研究熱點。Brotto等[5]提出,既然肌肉和骨骼作為兩種聯系緊密的器官,為什么肌肉-骨骼的關系是單向的,只有肌肉影響骨骼,反之則不然?如果骨骼能夠分泌大量因子,為什么這些因子不能對肌肉產生影響甚至優化肌肉功能?
骨骼腔隙系統與血管系統相連,骨組織細胞可釋放因子到血液循環,影響包括肌肉在內的其他組織器官功能。研究表明,骨鈣素(osteocalcin,OCN)、無翅型MMTV整合位點家族成員3a(wingless MMTV integration site family member 3a,Wnt3a)、前列腺素E2(prostaglandin E2,PGE2)、胰島素樣生長因子-1(insulin-like growth factor 1,IGF-1)、機械生長因子(mechano growth facoors,MGF)、血管內皮生長因子(vascular endothelial growth factor,VEGF)、肝細胞生長因子(hepatocyte growth factor,HGF)和轉化生長因子(transforming growth factor-β,TGF-β)等因子均可對肌肉功能進行調控(圖1)。本文擬對骨骼源性因子與肌組織代謝之間的關系進行探討,以全新的角度探究肌骨間的分子串擾,以期加深對肌骨系統的認識與理解,為探索肌骨疾病“一石二鳥”的病理學、藥理學及運動干預方案提供新的依據。
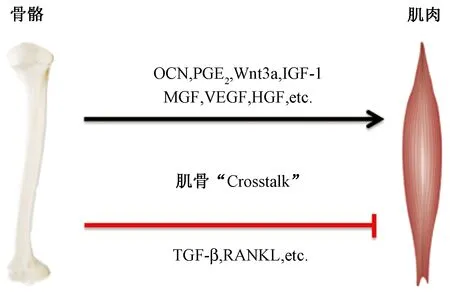
圖1 骨骼源性因子對骨骼肌的影響示意圖[5-7]Fig.1 Schematic diagram of the effect of bone-derived factors on the skeletal muscle[5-7]
1 骨鈣素
OCN是一種骨特異性非膠原蛋白,在骨骼系統中發揮重要作用。一方面作為骨礦化的調節劑,另一方面通過調節成骨細胞和破骨細胞活性來調控骨代謝[8]。OCN可分為羧化完全骨鈣素(carboxylated osteocalcin,cOCN)和羧化不全骨鈣素(uncarboxylated osteocalcin,uOCN)。其中,uOCN可進入循環激活包括胰腺、腦和睪丸等多組織生物學功能[9]。最新研究發現,骨骼肌也是uOCN的靶器官之一,uOCN可通過其特定的G蛋白偶聯受體C家族6組A亞型(GPRC6A),促進肌細胞生長并增加肌纖維強度[10-14]。在動物模型實驗中,外源性uOCN的補充可增加小鼠趾長伸肌橫截面積和抓握力,促進C2C12成肌細胞培養過程中肌管形成[13]。Mera等[15]研究提出,uOCN可增加老年鼠肌管中蛋白質合成,與健康對照相比,OCN-/-鼠表現出肌肉質量低和力量弱的特點。此外,人體實驗數據顯示,uOCN濃度過低會引發中老年女性的髖屈肌、髖外展肌和股四頭肌力量不足[12]。
肌肉衰老常伴隨著關鍵的肌肉細胞信號通路蛋白激酶B(protein kinase B,PKB,即Akt)/雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)和細胞外調節蛋白激酶(extracellular regulated kinase1/2,ERK1/2)的活化程度下降[16]。uOCN在C2C12成肌細胞中順序激活PI3K /Akt和p38MAPK途徑誘導成肌細胞增殖,激活GPRC6A/ERK1/2信號傳導機制增強肌源性分化,GPRC6A缺失會抑制C2C12細胞中Akt、p38絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和肌管中ERK1/2磷酸化,延緩肌細胞增殖與分化并下調MyHC表達[17]。此外,uOCN蛋白表達水平以肌肉類型特異性方式與mTORC1和叉頭框轉錄因子(forkhead box O,FOXO)信號通路中若干因子的表達和活性密切相關。循環uOCN和GPRC6A水平降低可引起相關肌肉類型的肌萎縮,加大后肢固定作用對uOCN/GPRC6A/ERK以及uOCN/GPRC6A/單磷酸腺苷活化蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)信號級聯的影響[18]。
研究證實,OCN可間接調控骨骼肌糖脂代謝,發展肌肉耐力。肌纖維GPRC6A- / -小鼠運動能力低于對照鼠約30%,運動后全身最大耗氧量,離體肌纖維耗氧量和骨骼肌內ATP濃度相對較低[19-20]。環磷腺苷效應元件結合蛋白(cAMP-response element binding protein,CREB)介導OCN調節肌管中葡萄糖攝取和肌纖維中糖酵解[21]。OCN信號活化可增強運動過程中肌糖原分解,誘導葡萄糖轉運蛋白4(glucose transporter 4,GLUT4)向質膜轉運[22]。通過這些組合功能,OCN可提供增強三羧酸循環進程所必需的碳原子,從而補充骨骼肌運動時所需ATP。此外,骨骼作用于肌肉引發的“前饋調節環”決定了機體的運動適應[19]。首先,OCN增強肌纖維中葡萄糖和脂肪酸的攝取及分解代謝;其次,在運動過程中,由OCN引發的肌肉IL-6分泌增加,又促進葡萄糖和脂肪酸產生;最后,IL-6可通過調節骨吸收,增加uOCN的釋放。綜上,外源性uOCN補充可作為肌肉減少癥和骨骼肌功能紊亂的潛在治療方法,OCN介導的運動干預機制能夠有效緩解老年人和體弱患者肌肉流失,維持其運動系統工作能力。
2 無翅型MMTV整合位點家族成員3a
Wnt3a是經典Wnt/β-catenin通路的重要配體,通過旁分泌、自分泌的方式釋放于胞外結合細胞表面受體,以激活下游信號產生級聯效應。研究鑒定了Wnt3a介導的成骨細胞反饋機制,在成骨細胞增殖或分化的早期階段起促進作用,在成熟和礦化過程中起抑制作用。Wnt3a可上調Pdgfa、Cyr61和Tgfb3等早期成骨調節因子表達,下調Vdr和Rspo2等成熟和礦化的調節因子表達[23]。此外,Wnt3a還可阻斷c-Fos表達減弱核因子KB受體活化因子配體(receptor activator of NF-KB ligand,RANKL)驅動的破骨細胞生成,逆轉骨髓細胞響應RANKL升高的β-catenin磷酸化水平,并且還原RANKL誘導的Trap和Oscar等破骨細胞標記基因表達[24-25]。
研究表明,骨骼分泌Wnt3a受機械應力調控,繼而向肌細胞發出信號。骨細胞樣MLO-Y4細胞條件培養基(conditioned medium,CM)能夠使比目魚肌收縮力增加約25%,而CM內特異性抑制Wnt3a則顯著減弱了這種影響。Wnt3a可通過激活Wnt、Notch和信號傳導及轉錄激活因子3(signal transducers and activators of transcription 3,STAT3)等信號通路調節C2C12成肌細胞分化[26]。Ridgeway等[27]研究顯示,Wnt3a可激活MyoD和Myogenin等肌源性調節因子,并刺激β-catenin易位至C2C12肌細胞核,促進肌源性標志物MyoG、MHC和β-catenin途徑下游基因過表達。此外,骨源性Wnt3a作為調節肌組織細胞內Ca2+信號和Wnt/β- catenin途徑的重要媒介,還參與調控細胞內Ca2 +穩態作為增強肌肉收縮力的觸發機制,誘導肌質網(sarcoplasmic reticulum,SR)鈣釋放、SR鈣吸收和存儲鈣進入成肌細胞分化進程,并上調對肌管穩態和細胞肥大起關鍵作用的Ca2+信號基因(CamkII、Nfatc3和Srf)以及與線粒體生物合成和氧化應激反應相關基因(Pgc1α和Sod1)的表達[26]。相關研究還表明,Wnt3a具有促進大鼠骨髓間充質干細胞(bone marrow mesenchymal stem cells,BMSCs)擴增的能力[28]。一方面,Wnt3a可誘導β-catenin核轉位并激活BMSCs中Wnt通路,加快細胞增殖并保持分化潛力;另一方面,Wnt3a可通過下調CCAAT/增強子結合蛋白α(CCAAT/enhancer binding protein α,C/EBPα)和過氧化物酶體增殖物激活受體γ(proliferator-activated receptor gamma,PPARγ)的表達抑制BMSCs成脂分化[29]。Wnt3a信號介導BMSCs肌源性分化,為人類基于細胞水平的組織修復提供合理的理論基礎。
3 前列腺素E2
PGE2是一種內源性脂質介質,由花生四烯酸在磷脂酶、環氧合酶和前列腺素E合成酶作用下代謝產生。骨骼系統中,PGE2可參與調節成骨細胞及其前體細胞、破骨細胞和骨細胞功能及細胞間通訊[30-31]。PGE2通過激活p38MAPK、p44/p42 MAPK和應激活化蛋白激酶(stress-activated protein kinase,SAPK)/c-Jun 氨基末端激酶(c-Jun N-terminal kinase,JNK),在成骨細胞樣MC3T3-E1細胞中刺激骨保護素(osteoprotegerin,OPG)合成,促進骨形成[32]。另有研究發現,PGE2可刺激RANKL產生,以環磷酸腺苷(cyclic Adenosine monophosphate,cAMP)依賴性方式抑制成骨細胞中OPG的產生[33-34]。上述結果可能源于骨組織細胞內4種G蛋白偶聯受體亞型(EP1、EP2、EP3、EP4)通過不同的途徑發出信號,引起PGE2在骨骼中發揮雙向功能[35]。
骨源性PGE2主要結合EP4調控骨骼肌代謝,Mo等[36]實驗研究顯示,PGE2可誘導肌源性分化引起,MyoD和MyoG等轉錄調節因子表達上調。此外,PGE2結合EP4可加速細胞周期中G1-S相變以促進原代成肌細胞增殖,并作為骨骼肌中氧化應激調節劑,減少EP4誘發的成肌細胞活性氧產生。近期研究表明,成骨細胞和骨細胞中間隙連接蛋白43(connexin 43,Cx43)缺失會導致體內肌纖維形成受損,而機械應力增加骨細胞中PGE2釋放,可激活cAMP/激酶A(protein kinase A,PKA)和磷脂酰肌醇3-激酶(phosphoinositide 3-kinase,PI3K)/Akt途徑,誘導糖原合成酶3(glycogen synthase kinase 3,GSK-3)磷酸化和失活,β-catenin核轉位增加[11]。細胞核中β-catenin參與Cx43啟動子調控,上調Cx43 mRNA表達,逆轉由Cx43缺失引起的肌組織損傷[30]。另有研究發現,肌肉損傷引發系列炎癥反應,PGE2作為炎癥細胞因子,可結合EP4直接靶向肌組織,引起肌肉擴張,在許多病理性疾病如肌肉萎縮、肌肉營養不良中具有顯著意義[37-38]。骨細胞中PGE2分泌量比肌肉細胞多1 000倍,超生理水平的PGE2作用于損傷的肌肉,可促進骨骼肌的重塑和修復[39]。值得關注的是,骨骼肌中還存在一種靜止的肌肉特異性干細胞(muscle specific stem cells,MuSCs),可在整個生命過程進行組織再生,而MuSCs中EP4特異性遺傳消融將會間斷PGE2信號傳導,阻止此進程以導致肌力下降。PGE2和EP4相互作用可通過激活誘導增殖的轉錄因子NURR1的cAMP/phosphor-CREB途徑引起MuSCs擴增。由此,PGE2信號轉導能夠作為肌肉干細胞功能的“變阻器”。PGE2由骨細胞合成并分泌到干細胞生態位中以響應損傷,誘發MuSCs增殖,是肌纖維受損后修復的關鍵步驟[40]。
4 其他因子
離體實驗研究顯示,流動流體剪切應力可增加骨細胞樣MLO-Y4細胞分泌IGF-1、MGF、VEGF和HGF等因子[41]。IGF-1是骨基質中最豐富的生長因子,在成年期參與維持骨量。Xian等[42]報道,骨重建期間從骨基質釋放的IGF-1可激活mTOR刺激募集的MSCs向成骨細胞分化,以維持適當的骨微結構和質量。進入血液循環后,IGF-1還能夠促進成肌細胞增殖和分化,在機體發育過程中作為調節肌肉量的重要因子。骨骼肌中IGF-1信號傳導下游Akt活化會引發快速且顯著的肌肉肥大效應,絕對力量增加[43]。MGF作為IGF-1的剪接變構體和局部自分泌的生長因子,可通過延長細胞周期的G1期,在特異性培養基誘導期間增強BMSCs成骨分化。骨骼肌損傷后,MGF先于衛星細胞激活發生,同時促進衛星細胞增殖[44]。研究表明,BMSCs通過旁分泌釋放VEGF刺激成肌細胞增殖。Sassoli等[45]向C2C12細胞中添加MSCs衍生CM可增強VEGFR磷酸化并提高其表達水平,而KRN633(VEGFR抑制劑)處理后導致受體活化顯著減少,并減弱MSCs-CM對C2C12細胞生長和Motch-1(成肌細胞活化和增殖的關鍵決定因子)信號轉導的影響。HGF在骨形成中發揮多重功能,參與維持骨再生,血管生成和成骨細胞與破骨細胞之間的平衡,不僅刺激成骨細胞分化并抑制礦化,也可刺激破骨細胞生成。而骨源性HGF快速釋放可促進功能性肌肉組織的重塑并增強骨骼肌再生[46-47]。
自1988年鑒定BMP-2作為骨和軟骨形成的有效誘導劑以來,BMP超家族信號成為脊椎動物骨骼生物學研究中最豐富的信號之一[48]。成骨細胞產生的BMP-2可自由分泌或結合到骨基質中,隨后在破骨細胞介導的骨吸收過程中被釋放。結構活躍的BMP信號能促進去神經支配后的肌肉肥大并減少肌肉萎縮。另有研究還闡明了BMP/Smad1、5、8信號轉導在促進肌肉生長和維持成人肌肉力量中的重要作用[49- 50]。
骨組織中的TGF-β主要由骨細胞、成骨細胞和破骨細胞等通過自分泌和旁分泌途徑合成。與引起骨骼肌肥大反應的骨衍生因子相反,骨骼在病理狀態下會分泌過量TGF-β及其家族成員肌肉生長抑制素、活化素和生長轉化因子-11,通過激活素受體2B發出信號,抑制肌肉功能[51-52]。盡管TGF-β和后者均導致肌肉功能障礙,但它們的模式卻不同。腺相關病毒(adeno-associated viral vector,AAV)載體在肌肉中表達時,活化素會誘發全身性肌肉萎縮和惡病質。在活化素水平表達較高的肌組織中,存在嚴重的肌肉質量損失和峰值力降低,但比力值沒有變化。相比之下,體內用TGF-β處理后肌肉質量并無變化,但其纖維化和橫截面積降低,導致骨骼肌峰值力和比力值均降低[53]。Regan等[54]研究認為,Nox4/阿諾堿受體1(ryanodine receptor 1,RyR1)信號軸參與骨基質釋放的TGF-β引發RyR1氧化應激和骨骼肌內Ca2+穩態失衡,嚴重影響肌肉功能。
表1骨骼源性因子介導的骨骼肌生理進程與功能
Table1Physiological processes and functions of the skeletal muscle mediated by bone-derived factors

骨骼源性因子參與的骨骼肌生理進程與功能OCN誘導成肌細胞增殖與分化;改善肌肉能量代謝;增強肌肉力量和耐力Wnt3a誘導肌源性分化;調節氧化應激PGE2減輕炎癥反應;修復肌纖維損傷IGF-1誘導成肌細胞增殖與分化;增強肌肉力量MGF促進肌衛星細胞增殖VEGF促進成肌細胞增殖HGF增加肌肉重塑和再生BMP-2加快肌肉生長;維持肌肉力量TGF-β降低肌肉功能
5 小結與展望
肌肉與骨骼同屬運動系統兩大器官,在維持人體健康中的作用不容忽視。骨源性OCN等對骨骼肌代謝的正向調控包括:①促進成肌細胞增殖,誘導肌源性分化;②維持骨骼肌力量素質,減少氧化應激,增強肌骨系統運動適應性;③提高肌肉組織損傷后再生和重塑能力,加速肌組織修復(表1)。然而,在病理學狀態下,肌組織微環境中的骨骼源性TGF-β增加,會損害骨骼肌。骨質疏松患者體內骨重構失衡,引發多種骨骼源性因子分泌異常,潛在地影響肌組織結構與功能,降低肌肉質量。未來研究亟待解決如下問題:首先,多數骨骼源性因子能夠改善肌肉功能,但其發揮作用的最佳劑量尚無定論;其次,不同骨骼源性因子在肌組織代謝調控中是否存在交互效應甚至于相互制約的影響尚不明晰;再次,骨硬化蛋白等因子可制約骨骼肌發展[6],采取何種手段抑制其介導的肌骨間交流途徑尚不確定;最后,運動干預可直接刺激肌骨系統,延緩肌肉衰減和骨質疏松的發生,但運動對骨骼源性因子的調節效果尚不知曉。建議下一步以骨骼為作用靶點,繼續探尋骨骼源性因子在肌骨系統中的作用機理,力求同時提高骨骼和肌肉的強度和功能,為降低肌骨疾病的發生提供新的著力點。
