外源赤霉素誘導矮生山茶恨天高植株生長的轉錄組分析
2020-03-27 12:19:37王江英朱朋波湯雪燕孫明偉葛金濤趙統利李紀元惠林沖邵小斌
江蘇農業學報 2020年1期
王江英 朱朋波 湯雪燕 孫明偉 葛金濤 趙統利 李紀元 惠林沖 邵小斌


摘要:對0 mg/L、800 mg/L赤霉素處理的矮生山茶恨天高葉片、莖段和莖尖進行了Illumina HiSeq高通量轉錄組測序,共獲得476 635個Unigenes,其中181 115(38.00%)個Unigenes獲得了注釋;差異基因分析發現了43 624個差異基因,其中包括43 295個特異的和329個共同的差異基因;GO和KEGG富集分析結果顯示,恨天高葉片增大、莖段伸長及莖尖生長與糖代謝、苯丙烷生物合成代謝相關;莖尖生長還與植物激素信號傳導和二萜生物合成有關。通過赤霉素處理后恨天高葉片、莖段、莖尖轉錄組分析發現,莖尖組織中參與赤霉素生物合成的KAO、GA20ox、GA3ox和GA2ox基因表達量發生變化,赤霉素信號傳導過程中DELLA蛋白調控以及赤霉素和生長素、細胞分裂素等其他激素間的互作促進恨天高植株生長。
關鍵詞:矮生恨天高;赤霉素;轉錄組;差異表達基因
中圖分類號:S685.14文獻標識碼:A文章編號:1000-4440(2020)01-0047-10
Abstract:Illumina HiSeq high-throughput sequencing was performed on the leaves, stem segments and shoot tips of Camellia reticulata Hentiangao treated with ?mg/L0 and 800 mg/L gibberellin, respectively. A total of 476 635 unigenes were obtained, and 181 115 (38.00%) of them were annotated. In addition, 43 624 differentially expressed genes (DEGs), including 43 295 unique and 329 common, were identified. GO and KEGG enrichment analysis results showed that leaf enlargement, stem elongation and shoot tip growth were related to glucose metabdism and phenylpropanoid biosynthesis. Furthermore, many DEGs in shoot tip growth were involved in plant hormone signal transduction and diterpenoid biosynthesis. According to the transcriptome analysis, three factors were found to promote the growth of Hentiangao. The first was the expression changes of KAO, GA20ox, GA3ox and GA2ox genes involved in gibberellin biosynthesis in shoot tips. The second was the regulation of DELLA protein in gibberellin signal transduction. The third was the interaction of gibberellin with auxin, cytokinin and other hormones.
Key words:Camellia reticulata Hentiangao;gibberellin;transcriptome;differentially expressed genes
赤霉素(Gibberellins,GAs),二萜類植物激素,主要促進植株的營養生長,包括節間的延長、根的伸長和葉的擴展[1-3]。在赤霉素合成途徑中,萜烯合成酶TPSs、細胞色素P450s和GA氧化酶(GAoxs)將前體牻牛兒牻牛兒焦磷酸GGDP催化成活性赤霉素,其中貝殼杉烯酸氧化酶(KAO)屬于細胞色素P450,催化赤霉素合成前期階段。GA氧化酶包括GA2ox、GA20ox和GA3ox,是赤霉素合成和代謝后期階段的主要調控點,將非活性赤霉素催化成活性赤霉素[4-7]。前期研究發現,多種植物的KAO突變能誘導植株矮化,如水稻d35突變體[8]。梨矮化砧木中KAO表達量低于喬化和半矮化的品種[9]。相對于早期合成酶,后期生物合成酶易受植物生長發育和環境因素調節,這些酶在生物活性赤霉素水平的調控中起著關鍵作用。GA20ox和GA3ox過量表達后提高活性赤霉素的含量,加快植物生長,節間伸長,GA20ox和GA3ox功能缺失會引起植株矮化[10-11]。GA2ox的作用則是降低活性赤霉素的水平[11-12]。
赤霉素信號傳導途徑中,DELLA蛋白作為植物生長抑制劑可被活性赤霉素抵消[13],赤霉素信號啟動DELLA蛋白的降解,調控赤霉素反應基因的表達,從而促進植物的生長,所以DELLA蛋白對植物的生長發育起負反饋調節[14-16]。目前DELLA基因在擬南芥[17]、葡萄[18]、草莓[19]和甘藍型油菜[20]等植物中陸續被克隆及分析,為植物矮化性狀研究提供依據。
恨天高(Camellia reticulata Hentiangao)是云南山茶最珍貴的品種之一,其株形矮小,生長緩慢,花瓣平展,完全重瓣,形姿優美,葉濃花艷,具有極高的觀賞價值[21]。隨著城市的快速發展,矮生和盆栽木本花卉植物在園林綠化、居室美化等方面有著重要作用。通過轉錄組分析挖掘高效調控恨天高株高的關鍵基因,為培育微型山茶新品種奠定基礎。
本研究以云南山茶矮生品種恨天高為材料,利用轉錄組測序分析恨天高植株矮生與赤霉素合成代謝和信號傳導間的關系,以期獲得調控植物株高的重要基因,并揭示其矮生機制。
1材料與方法
1.1試驗材料
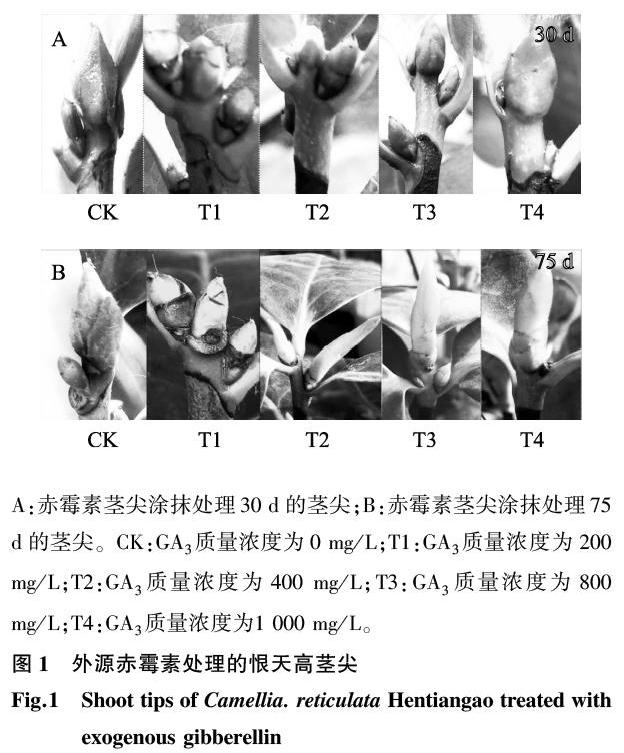
以立地條件相同且生長年齡相近的恨天高山茶(Camellia reticulate Hentiangao)為材料,利用不同質量濃度梯度[0 mg/L(CK)、200 mg/L(T1)、400 mg/L(T2)、800 mg/L(T3)和1 000 mg/L(T4)]的赤霉素(GA3)進行莖尖涂抹。75 d后采集各質量濃度處理后的葉片、莖段及莖尖組織進行液氮速凍,-80 ℃保存備用。
1.2莖尖和葉片總RNA提取
參照北京艾德萊生物科技有限公司RN53-EASYspin Plus植物試劑盒說明書提取樣品總RNA。利用1%的瓊脂糖凝膠電泳和Nanodrop 2000分光光度計檢測RNA完整度、濃度、A260/280、A260/230及核酸峰度是否正常,之后再經Agilent bioanalyzer 2100進行RNA質量檢測。
1.3赤霉素誘導恨天高植株生長的質量濃度篩選
分別利用0 mg/L、200 mg/L、400 mg/L、800 mg/L和1 000 mg/L的GA3涂抹5年生恨天高植株莖尖,每2 d涂1次,每個質量濃度處理3株,每株分別處理5個莖尖,30 d和75 d后分別測量莖尖生長值。
1.4文庫構建和轉錄組測序
恨天高葉片、莖段和莖尖總RNA質量分別檢測合格后,利用NEBNext Poly(A) mRNA Magnetic Isolation Module進行mRNA捕獲,利用NEBNext Ultra RNA Library Prep Kit for Illumina和Beckman Agencourt AMPure XP beads分別進行文庫構建和文庫純化,利用TruSeq PE Cluster Kit V4完成cBOT自動成簇,最后通過TrSeq SBS Kit v4-HS進行HiSeq測序。以上文庫構建和轉錄組測序均由蘇州金唯智生物科技有限公司完成。
1.5生物信息學分析
對測序結果原始圖像數據利用軟件Bcl2fastq(v2.17.1.14)進行圖像堿基識別,初步質量分析后得到原始測序數據。測序數據質量評估采用FastQC(v0.10.1)進行分析,使用Cutadapt(version1.9.1)對測序原始數據去除接頭以及低質量序列等,得到后續信息分析用的高質量有效序列。采用Trinity對樣品數據從頭組裝,組裝結果通過序列聚類進行進一步序列拼接和去冗余處理,得到長的非冗余Unigenes,利用TransDecoder軟件進行ORF分析[22]。
Unigenes表達計算使用RSEM軟件,該軟件使用FPKM(Fragment per kilo bases per million reads)方法計算基因表達量。利用EdgeR軟件進行基因差異表達分析,以變化2倍以上、Pvalue≤0.05且錯誤發現率(FDR)<0.01為差異表達基因篩選標準。最后基于差異表達基因進行GO功能注釋、KEGG代謝通路分析和COG功能注釋。
1.6實時熒光定量PCR驗證
利用實時熒光定量PCR驗證轉錄組測序的準確性,挑選5個差異表達基因設計引物,以山茶18S RNA為內標基因(所用引物見表1),生物學試驗重復3次。運用2-△△Ct計算方法求得差異基因的表達量。
2結果與分析
2.1外施赤霉素促進恨天高莖尖伸長
選擇立地條件相同、生長狀態良好且一致的三年生恨天高(Camellia reticulate Hentiangao)幼苗為材料(圖1A),利用不同質量濃度梯度的GA3進行莖尖涂抹(圖1B)。研究結果發現,GA3處理30 d后,對照組和試驗組莖尖長度差異顯著(P<0.05)(圖1C),其中CK的莖尖長度最短,T1、T2、T4處理分別是CK的1.23倍、1.58倍和1.92倍,T3為CK的2.53倍(圖2A);GA3處理75 d后,對照組和試驗組莖尖長度差異顯著(P<0.05)(圖1D),其中CK莖尖長度依舊最短,其次是T1處理和T2處理,為CK的1.59倍和1.93倍,而T4處理和T3處理分別為CK的2.27倍和3.61倍(圖2B)。比較發現,對照組和試驗組在2次測量的45 d間隔期,CK的莖尖少許伸長,試驗組除了T1處理長勢變化不明顯外,T2、T3、T4處理長勢旺,尤其T3處理(圖2C)。由此可以推測,外源GA3能誘導恨天高山茶的植株生長,并且隨著外施質量濃度的增高(0 mg/L、200 mg/L、400 mg/L),莖尖生長值也隨之增加,當質量濃度達800 mg/L時,莖尖生長最快,然而質量濃度達1 000 mg/L時,莖尖增加幅度卻有所下降,從而表明800 mg/L是誘導恨天高山茶植株生長的較佳質量濃度。
2.2轉錄組序列組裝及Unigene注釋分析
利用最佳赤霉素誘導質量濃度800 mg/L處理恨天高莖尖組織,0 mg/L的GA3處理作對照,75 d后采集葉片、莖段、莖尖進行轉錄組測序,其中葉片和莖段分別取2組樣品進行測試,莖尖只取1組樣品進行測試。
使用軟件Cutadapt對測序原始數據去除接頭以及低質量序列,獲得后續信息分析用的過濾數據。恨天高3種組織及對照共8個文庫產生了411 464 870條序列,約60.96 Gb轉錄組數據,Unigene平均長度為147.38~148.61 bp,G+C堿基含量為44.58%~46.84%(表2)。由表3可以看出,轉錄組數據經組裝產生了43 073 816個重疊群,476 635個Unigenes,平均長度為497.69 bp。在所有Unigenes序列中,序列長度小于500 bp的占71.60%。Unigene注釋分析顯示有181 115條Unigenes獲得注釋,占Unigenes總數的38.00%,其中獲得注釋較多的數據庫分別是Nr注釋172 121條(36.11%),COG注釋74 120條(15.55%),Swissport注釋106 818條(22.41%)和KEGG注釋25 837條(5.42%)(圖3)。
2.3差異表達基因篩選及分析
將Clean data比對到Unigene上,利用bowtie2軟件進行短reads比對,結果顯示,唯一比對到參考基因的reads數量在2.5×107~3.6×107,用于基因表達分析。在葉片、莖段和莖尖3種組織中,發現了特異的和共同的差異表達基因,共發現43 624個差異表達基因,包括43 295個特異差異表達基因和329個共同差異表達基因(圖4A)。
CK1(對照葉片樣品)與L1(800 mg/L外源赤霉素處理后葉片樣品1)比較中發現27 424個差異表達基因,其中上調表達基因16 495個,下調表達基因10 929個;CK1與L2(800 mg/L外源赤霉素處理后葉片樣品2)比較中發現23 450個差異表達基因,其中上調表達基因12 956個,下調表達基因10 494個;CK2(對照莖段樣品)與SS1(800 mg/L外源赤霉素處理后莖段樣品1)比較中發現20 773個差異表達基因,其中上調表達基因11 168個,下調表達基因9 605個;CK2與SS2(800 mg/L外源赤霉素處理后莖段樣品2)比較中發現21 040個差異表達基因,其中上調表達基因11 706個,下調表達基因9 334個;CK3(對照莖尖樣品)與ST1(800 mg/L外源赤霉素處理后莖尖樣品)比較中發現4 256個差異表達基因,其中上調表達基因1 734個,下調表達基因2 522個(圖4B)。在5組比較中,CK3與ST1比較獲得的上調和下調基因數量均最低,可能是由于莖尖組織相對于葉片和莖段而言,與植株伸長生長更為密切,獲得的差異表達基因控制植株生長的作用更大。
根據FPKM值將8個文庫中的基因表達水平分為極高、高、中、低和極低5類(圖4C),其中差異表達基因中占比例最大的是極低表達基因,FPKM為0~1,占比例最小的是極高表達基因,FPKM>60。另外,在葉片、莖段和莖尖等組織中檢測到共同差異表達基因329個,通過層次聚類分析將其劃分成A、B 2個簇(圖4D)。
2.4差異基因GO富集分析
通過GO富集分析對差異表達基因進行生物學功能研究,在錯誤發現率FDR<0.01時,43個GO條目富集在CK1/L1,42個GO條目富集在CK1/L2,39個GO條目富集在CK2/SS1,39個GO條目富集在CK2/SS2,37個GO條目富集在CK3/ST1。基于所有GO富集分析,確定37個共同GO類別和8個差異GO類別,在37個共同GO條目中,催化活性、結合、代謝過程、細胞過程、細胞組分和細胞器是富集基因最多的GO類別。在8個差異GO類別中,CK3/ST1無分子功能調節劑富集;鳥苷酸交換因子活性顯著富集在CK1/L1和CK1/L2;CK2/SS1和CK2/SS2無細胞外基質部分和細胞外區域部分富集;CK3/ST1無類核和細胞外基質富集;轉運狀態顯著富集在CK1/L1;CK3/ST1無生物附著富集。由此可以看出,在外源赤霉素處理下葉片、莖段和莖尖組織中的GO類別存在差異,誘導植株生長。
2.5差異基因KEGG富集分析
KEGG富集分析結果顯示,分別有30、30、18、14和27條代謝途徑富集在CK1/L1、CK1/L2、CK2/SS1、CK2/SS2和CK3/ST1,顯著富集的則為5、4、5、4和5條代謝途徑(圖5)。苯丙烷生物合成、戊糖和葡萄糖醛酸相互轉化過程是共同顯著富集的代謝途徑(圖5),倍半萜和三萜生物合成富集在CK1/L1、CK1/L2和CK2/SS2(圖5A,圖5B,圖5D),細胞色素P450對外源化合物代謝的影響、藥物代謝-細胞色素P450只富集在CK1/L1(圖5A),糖酵解/糖異生只富集在CK1/L2(圖5B),光合-天線蛋白、油菜素內酯生物合成只富集在CK2/SS1(圖5C),玉米素生物合成只富集在CK2/SS1和CK2/SS2(圖5C、圖5D),CK3/ST1顯著富集植物激素信號轉導、二萜生物合成和類胡蘿卜素生物合成(圖5E)。由此可以看出,CK3/ST1中所富集的植物激素信號轉導和二萜生物合成兩條途徑與恨天高植株生長關系較為密切,其他途徑則間接調控植株生長。
KEGG富集研究結果顯示,CK3/ST1所富集的植物激素信號轉導途徑中色氨酸代謝CH3、SAUR下調表達,玉米素生物合成AHP上調表達、A-ARR下調表達,二萜生物合成途徑DELLA下調表達,油菜素內酯合成TCH4上調表達、BSK和CYCD3下調表達(圖6A)。表明,植株生長受多種激素互作的影響,本研究中可以認為二萜生物合成途徑對植株生長首先起作用。KEGG富集結果顯示,二萜生物赤霉素生物合成途徑中KAO上調表達、GA20ox和GA3ox下調表達、GA2ox既有上調也有下調表達(CK1/L1上調,CK3/ST1下調)(圖6B)。說明,二萜生物合成途徑中DELLA、KAO、GA20ox、GA3ox和GA2ox調控恨天高植株生長。
2.6轉錄組的實時熒光定量PCR驗證
挑選5個差異表達基因,利用實時熒光定量PCR進行對照組和試驗組(800 mg/L GA3處理75 d)莖尖基因的表達量測定。結果顯示,實時熒光定量PCR(qPCR)與轉錄組測序(RNA-Seq)的結果在基因表達量變化幅度上具有一定差異,但基因的表達趨勢是一致的(圖7)。利用SPSS Pearson對qPCR和RNA-Seq進行雙變量相關性分析,結果顯示5個差異表達基因的qPCR和RNA-Seq在P<0.01水平(雙側)上顯著相關,相關系數為0.978(表4)。由此可以說明,轉錄組測序獲得的基因差異表達結果可靠。
3討論
恨天高作為天然矮生資源,是山茶矮化育種的優良親本材料。本研究采用Illumina HiSeq高通量技術對0 mg/L和800 mg/L GA3處理75 d的葉片、莖段和莖尖組織進行測序,組裝得到的181 115條Unigenes獲得了注釋。通過實時熒光定量PCR檢驗,qPCR和RNA-Seq結果顯著相關,因此,本研究轉錄組測序結果是可靠的。
GO富集分析發現,大量差異表達基因在催化活性、結合、代謝過程、細胞過程、細胞組分和細胞器等類別中顯著富集,在差異條目中,葉片、莖段和莖尖所富集的類別存在差異,可能是外源赤霉素對不同組織的誘導作用存在差異,也可能是組織本身存在差異所造成。KEGG通路分析發現,葉片、莖段和莖尖顯著富集的代謝途徑不完全相同,其中苯丙烷生物合成、戊糖和葡萄糖醛酸相互轉化過程是共同顯著富集的代謝途徑。苯丙烷生物合成是植物最重要的次生代謝途徑之一[23],而戊糖和葡萄糖醛酸相互轉化也是糖代謝的重要環節,參與植物多種發育過程[24],表明在外源赤霉素誘導過程中,苯丙烷生物合成、戊糖和葡萄糖醛酸相互轉化途徑中差異基因對恨天高植株生長具有重要作用。然而,相比于其他4組,CK3/ST1顯著富集的植物激素信號轉導、二萜生物合成和類胡蘿卜素生物合成途徑中差異表達基因直接參與調控恨天高植株生長。
本研究中,莖尖組織中外源赤霉素誘導的差異表達基因包括赤霉素信號傳導途徑中的DELLA,赤霉素生物合成途徑中的KAO、GA20ox、GA3ox和GA2ox等基因,DELLA、GA20ox、GA3ox和GA2ox下調表達,KAO上調表達。研究結果表明,DELLA蛋白在赤霉素信號傳導途徑中起中心開關的作用,對赤霉素誘導的反應非常靈敏,赤霉素處理5~10 min后蛋白水平顯著下降[25-26],本研究中外源赤霉素的誘導信號傳遞給DELLA蛋白,DELLA蛋白呈下調表達,從而誘導下游赤霉素代謝基因如KAO的上調表達。
高等植物赤霉素水平通過正反饋和負反饋來實現調節體內激素的平衡。當體內活性赤霉素水平低時,擬南芥GA20ox1、GA20ox2、GA20ox3和GA3ox1的表達量均顯著提高,外施GA3后,其表達量又顯著降低[27-28]。本研究中,外施赤霉素后恨天高莖尖中GA20ox、GA3ox也呈下調表達。當活性赤霉素水平增加時,擬南芥GA2ox表達量上調[29],在植物中過量表達GA2ox能誘導植株矮化,本研究中通過差異基因表達水平測定發現,莖尖中GA2ox下調表達,葉片中GA2ox呈上調表達,可能由于莖尖在外源赤霉素的誘導下,快速生長,積累較多的活性赤霉素,當活性赤霉素由莖尖傳輸到葉片時,則誘導葉片中的GA2ox上調表達,以抑制植株進一步生長,起到反饋調節作用。
另外,我們挑選了DELLA、GA20ox、GA3ox、GA2ox和KAO 5個差異表達基因進行了實時熒光定量PCR驗證,在莖尖組織中,實時熒光定量PCR(qPCR)與轉錄組測序(RNA-Seq)結果一致,DELLA、GA20ox、GA3ox、GA2ox呈下調表達,KAO上調表達。由此可以推測,800 mg/L外源赤霉素莖尖涂抹處理后,赤霉素質量濃度升高,DELLA蛋白接受信號呈下調表達,誘導下游KAO上調表達,GA20ox、GA3ox、GA2ox呈下調表達,多個基因相互作用,促進植株生長,當體內赤霉素質量濃度積累到一定程度時,GA2ox則反饋調節抑制植株繼續生長。外源赤霉素誘導植株生長的機理,以本文為基礎,后續還需要進一步深入研究。
本研究利用高通量轉錄組測序技術建立矮生山茶恨天高赤霉素誘導前后葉片、莖段和莖尖轉錄組數據庫,分析比較3種組織在外源赤霉素處理后生物反應途徑的差異,發現相對于葉片和莖段,莖尖對植株生長影響較大,進一步篩選出莖尖組織中的差異表達基因,為研究恨天高矮生機制以及培育矮生山茶品種提供基礎。
參考文獻:
[1]RICHAEDS D E, KING K E, AIT-ALI T, et al. How gibberellin regulates plant growth and development: a molecular genetic analysis of gibberellin signaling [J]. Annual Review of Plant Biology, 2001, 52: 67-88.
[2]張旭紅,孫美玉,李靖銳,等. 東方百合‘索邦GA20ox的克隆及表達分析[J]. 園藝學報, 2019, 46(1): 74-86.
[3]YAMAGUCHI S. Gibberellin metabolism and its regulation [J]. Annual Review of Plant Biology, 2008, 59: 225-251.
[4]DAVIDSON S E, ELLIOTT R C, HELLIWELL C A, et al. The pea gene NA encodes ent-kaurenoic acid oxidase [J]. Plant Physiology, 2003, 131(1): 335-344.
[5]LANGE T, HEDDEN P, GRAEBE J E. Expression cloning of a gibberellin 20-oxidase, a multifunctional enzyme involved in gibberellin biosynthesis [J]. Proceedings of the National Academy of Sciences, 1994, 91(18): 8522-8566.
[6]APPLEFORD N E J, LENTON J R. Gibberellins and leaf expansion in near-isogenic wheat lines containing Rht1 and Rhe3 dwarfing alleles [J]. Planta, 1991, 183(2): 229-236.
[7]COLES J P, PHILLIPS A L, CROKER S J, et al. Modification of gibberellin production and plant development in Arabidopsis by sense and antisense expression of gibberellin 20-oxidase genes [J]. The Plant Journal, 1999, 17(5): 547-563.
[8]SAKAMOTO T, MIURA K, ITOH H, et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice [J]. Plant Physiology, 2004, 134(4): 1642-1653.
[9]歐春青,姜淑苓,王斐,等. 梨貝殼杉烯酸氧化酶基因PcKAO1的克隆與表達分析[J]. 園藝學報, 2013, 40(5): 849-858.
[10]WANG J Y, WU B, LI J Y, et al. Overexpression and silent expression of CrGA20ox1 from Camellia reticulate ‘Hentiangao and its effect on morphological alterations in transgenic tobacco plants [J]. Plant Breeding, 2018, 9(3): 1-9.
[11]HUANG Y, WANG X, GE S, et al. Divergence and adaptive evolution of the gibberellin oxidase genes in plants [J]. BMC Evolutionary Biology, 2015, 15: 207-221.
[12]THOMAS S G, PHILLIPS A L, HEDDEN P. Molecular cloning and functional expression of gibberellin 2-oxidases, multifunctional enzymes involved in gibberellin deactivation [J]. Proceedings of the National Academy of Sciences, 1999, 96(8): 4698-4703.
[13]李巧峽,張麗,王玉,等. 赤霉素調控植物開花及器官發育的研究進展[J]. 中國細胞生物學學報, 2019, 41(4): 746-758.
[14]WILLIGE B C, GHOSH S, NILL C, et al. The DELLA domain of GA INSENSITIVE mediates the interaction with the GA INSENSITIVE DWARF1A gibberellin receptor of Arabidopsis [J]. The Plant Cell, 2007, 19(4): 1209-1220.
[15]DILL A, THOMAS S G, HU J, et al. The Arabidopsis F-Box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation [J]. The Plant Cell, 2004, 16(6): 1392-1405.
[16]SUN T P. Gibberellin-GID1-DELLA: a pivotal regulatory module for plant growth and development [J]. Plant Physiology, 2010, 154(2): 567-570.
[17]CHEN J H, CHENG T L, WANG P K, et al. Genome-wide bioinformatics analysis of DELLA-family proteins from plants [J]. Plant Omics, 2013, 6(3): 201-207.
[18]張文穎,王晨,朱旭東. 葡萄全基因組DELLA 蛋白基因家族鑒定及其應答外源赤霉素調控葡萄果實發育的特征[J]. 中國農業科學, 2018, 51(16): 3130-3146.
[19]LI W J, ZHANG J X, SUN H Y. FveRGA1, encoding a DELLA protein, negatively regulates runner production in Fragaria vesca [J]. Planta, 2018, 247(4): 941-951.
[20]賈永鵬,李開祥,昝領兄,等. 甘藍型油菜全基因組DELLA蛋白基因家族的鑒定和表達分析[J]. 中國油料作物學報, 2019, 41(3): 360-368.
[21]管開云.中國茶花圖鑒[M]. 杭州: 浙江科學技術出版社, 2014:53-54.
[22]張恩亮,馬玲玲,楊如同,等. IBA誘導楸樹嫩枝扦插不定根發育的轉錄組分析[J]. 林業科學, 2018, 54(5): 48-61.
[23]WANG Y, WEATHERS P J. Sugar proportionately affect artemisinin production [J]. Plant Cell Report, 2007, 26(7): 1073-1081.
[24]HU Y L, GAI Y, YIN L, et al. Crystal structures of a Populus tomentosa 4- coumarate: CoA ligase shed light on its enzymatic mechanisms[J]. Plant Cell, 2010,22(9):3093-3104.
[25]ITOH H, UEGUCHI-TANAKA M, SATO Y, et al. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei [J]. Plant Cell, 2002, 14(1): 57-70.
[26]TYLER L, THOMAS S G, HU J, et al. DELLA proteins and gibberellin regulated seed germination and floral development in Arabidopsis [J]. Plant Physiology, 2004, 135(2): 1008-1019.
[27]MITCHUM M G, YAMAGUCHI S, HANADA A, et al. Distinct and overlapping roles of two gibberellin 3-oxidases in Arabidopsis development [J]. The Plant Journal, 2006, 45(5): 804-818.
[28]RIEU I, RUIZ-RIVERO O, FERNANDEZ-GARCIA N, et al. The gibberellin biosynthetic genes At GA20ox1 and At GA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle [J]. The Plant Journal, 2008, 53(3): 488-504.
[29]RIEU I, THOMAS S, POWERS S J, et al. Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis [J]. Plant Cell, 2008, 20(9): 2420-2436.
(責任編輯:陳海霞)
