基于生物信息學分析的hsa-miR-206靶基因預測
2020-04-02 07:05:28康治理武振方劉曉偉
東南國防醫藥 2020年2期
康治理,武振方,劉曉偉,許 斌
0 引 言
MicroRNAs(miRNAs)是一種長度約為19~24個堿基的單鏈小分子的非編碼RNA家族,它是由一段具有發卡結構的70~80個堿基大小的單鏈RNA前體經過Dicer酶的作用剪切生成,通過并入到RNA誘導的沉默復合物(RISC)與靶mRNA的5’及3’UTR端(3' untranslated regions,3'-UTR)特異性堿基配對,在轉錄水平抑制靶mRNA翻譯或者誘導靶mRNA降解,從而調控靶基因的表達[1]。其靶向mRNA的主要方式包括翻譯抑制、mRNA切割和mRNA穩定性改變。早期研究發現,miRNA對生物學過程的調控至關重要,如發育時間、細胞死亡、細胞增殖、免疫、神經系統模式等[2-3]。越來越多的數據表明,miRNA是許多重要的生命過程的主要調節者,這些生命過程包括細胞增殖、細胞凋亡、病毒感染和細胞癌變等[4-5]。
miR-206是目前最具研究潛力及特性的microRNAs之一,最初在骨骼肌中發現并被認為是骨骼肌特異性miRNA[6]。參與了包括腫瘤在內的諸多疾病的發病機制,目前研究表明miR-206在腫瘤等多種疾病中異常表達并在增殖、分化、凋亡、侵襲和轉移中發揮重要作用[7]。miR-206定位在人類第6號染色體上,是骨骼肌特異性表達的“肌特異性miRNA(myomiR)”家族成員之一[8]。miR-206通過與其靶mRNA的3'端非翻譯區堿基不完全互補配對,抑制mRNA翻譯或直接使其降解而發揮生物學特性,調控細胞的增殖、分化、轉移等生物學行為[9]。靶基因與miR-206結合位點越多則受其調控程度越大,由此形成一個巨大分子調控網絡,在個體成長發育、生理過程及疾病發展過程中發揮重要作用。然而其通過作用于哪些靶基因,發揮何種生物學效應在許多實驗研究中都尚未闡釋清楚,有關hsa-miR-206的生物信息學研究也較少。本研究擬通過生物信息學的方法探索對hsa-miR-206進行定位和序列保守性研究、靶基因預測、功能富集分析(GO分析)、信號通路分析(KEGG)、蛋白互作分析(protein protein interaction,PPI)等分析,為今后進一步探索hsa-miR-206在腫瘤發生、發展及分子機制等方面提供線索和理論依據。
1 材料與方法
1.1 miR-206的序列保守性分析應用miRbase(http://www.mirbase.org/)在線查找各物種已被明確的成熟堿基序列,并對比分析miR-206序列在各物種之間的保守性,用美國國家醫學圖書館國家生物技術信息中心(National Center for Biotechnology Information, NCBI)開發的生物大分子序列比對搜索工具(Basic Local Alignment Search Tool,BLAST)(https://blast.ncbi.nlm.nih.gov/Blast.cgi)進一步分析其物種保守性。
1.2miR-206的靶基因預測運用在線靶基因預測軟件,miRDB(http://www.mirdb.org/),TargetScan V7.2(http://www.targetscan.org/)和DIANA-microT(http://diana.imis.athenainnovation.gr)等3個數據庫對has-miR-206可能存在的靶基因進行預測,為得到更加準確的結果,用Venn Diagrams在線作圖工具畫韋恩圖得到3個數據庫預測結果的交集來降低假陽性率。
1.3miR-206靶基因的GO功能注釋及KEGG通路富集分析運用DAVID6.8(database for annotation, visualization and integrated discovery)(http://David.abcc.ncifcrf.gov/)對靶基因集合進行GO分析以及基于KEGG數據庫(Kyoto Encyclopedia of Genes and Genomes)(http://www.genome.jp/)的Pathway分析,選擇人類全基因組作為背景基因,用"Functional Annotation Chart"分析工具計算P值,以P<0.05為顯著性閾值,分別得到基因集合相對于背景具有統計學意義的GO分析與Pathway分析結果。
1.4關鍵靶基因篩選利用在線工具STRING(https://string-db.org)[10]聯合Cytoscape軟件[11]構建miR-206靶基因蛋白互作網絡(PPI),并運用Mcode插件[12]篩選出關鍵基因(hub gene)。
2 結 果
2.1 miR-206的染色體定位及序列保守性分析檢索在線數據庫miRBase,發現人has-miR-206基因序列號為MIMAT0000462,其成熟體堿基序列為“53-UGGAAUGUAAGGAAGUGUGUGG-74”。通過NCBI BLAST對小鼠(mmu)、大鼠(rno)、野豬(ssc)等10個物種的miR-206序列對比分析,發現miR-206的成熟序列在各物種間高度保守,見表1。
2.2hsa-miR-206的靶基因預測結果使用TargetScan、miRDB、DIANA-microT 3個在線預測軟件分別得到hsa-miR-206的靶基因分別為897個、944個、1148個,再進行交集得到預測靶基因420個,見圖1。
2.3hsa-miR-206靶基因GO功能注釋和KEGG通路富集分析將3個經典數據庫預測到的靶基因取最終交集420個作GO分析,發現miR-206的靶基因主要富集在高爾基體、內膜、囊泡等細胞組件(P<0.05),參與DNA結合、蛋白結合、轉錄調節活性等分子功能(P<0.05),富集于生物合成調節、有機物代謝過程調節、轉錄調節等生物學過程(P<0.05)。見表2。KEGG信號通路分析顯示主要富集在Wnt信號通路、T細胞受體信號通路、癌癥通路、調節肌動蛋白細胞骨架等(P<0.05)。見表3。
表1 部分物種成熟的miR-206保守序列

序列號(miRBase)物種名稱序列(5'→3')MIMAT0000462人hsa-miR-20653-UGGAAUGUAAGGAAGUGUGUGG-74MIMAT0000239小鼠mmu-miR-206-3p46-ACAUGCUUCUUUAUAUCCUCAUA-67MIMAT0017004小鼠mmu-miR-206-5p8-ACAUGCUUCUUUAUAUCCUCAUA-30MIMAT0017154褐家鼠rno-miR-206-5p13-ACAUGCUUCUUUAUAUCCUCAU-34MIMAT0000879褐家鼠rno-miR-206-3p51-UGGAAUGUAAGGAAGUGUGUGG-72MIMAT0006606家犬cfa-miR-20639-UGGAAUGUAAGGAAGUGUGUGG-60MIMAT0036052山羊chi-miR-20653-UGGAAUGUAAGGAAGUGUGUGGU-75MIMAT0023846黑線倉鼠cgr-miR-20650-UGGAAUGUAAGGAAGUGUGUGG-71MIMAT0002318猩猩ppy-miR-20653-UGGAAUGUAAGGAAGUGUGUGG-74MIMAT0013864野豬ssc-miR-20650-UGGAAUGUAAGGAAGUGUGUGA-71
表2 miR-206靶基因的GO功能分析
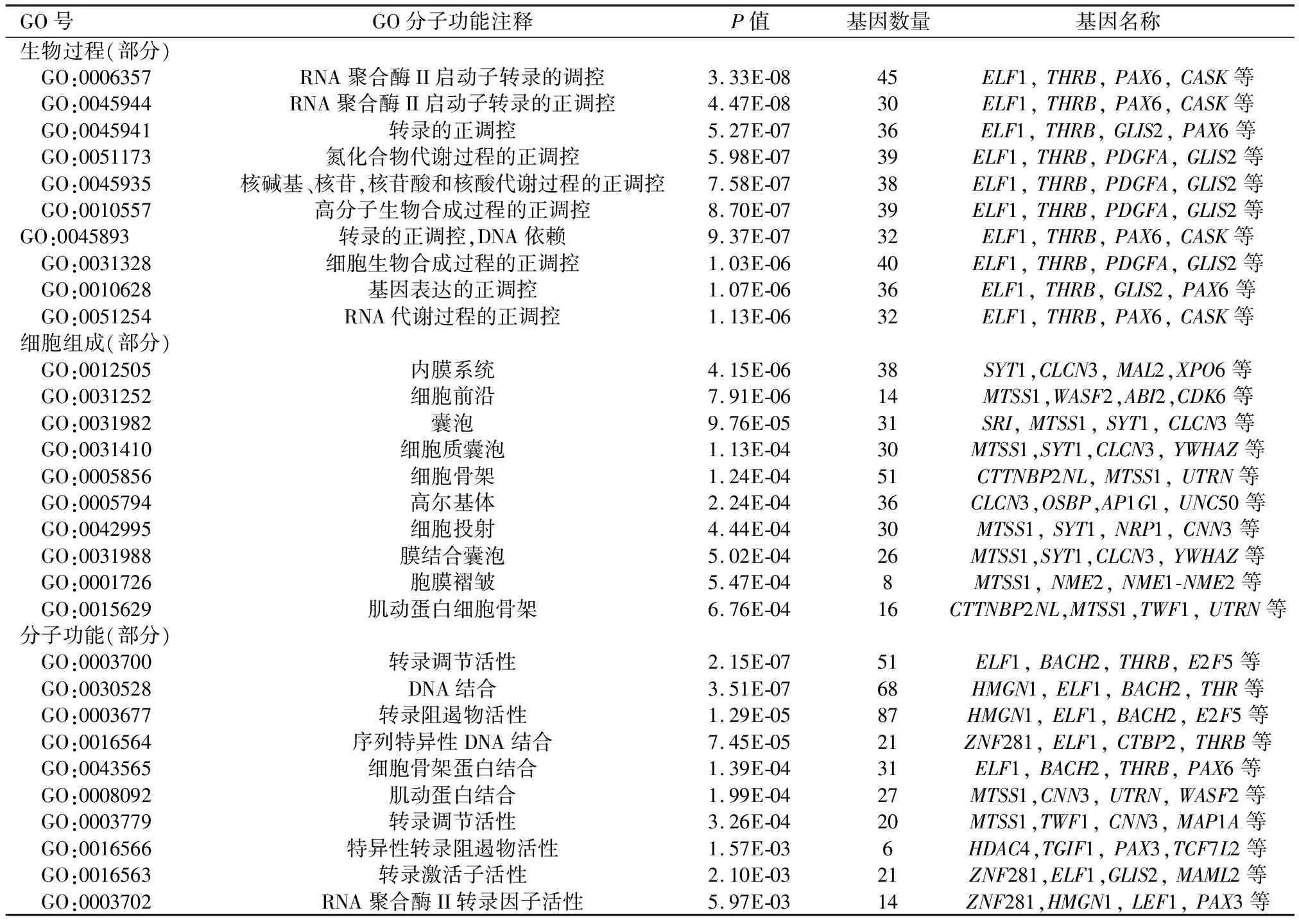
GO號GO分子功能注釋P值基因數量基因名稱生物過程(部分) GO:0006357RNA聚合酶II啟動子轉錄的調控3.33E-0845ELF1,THRB,PAX6,CASK等 GO:0045944RNA聚合酶II啟動子轉錄的正調控4.47E-0830ELF1,THRB,PAX6,CASK等 GO:0045941轉錄的正調控5.27E-0736ELF1,THRB,GLIS2,PAX6等 GO:0051173氮化合物代謝過程的正調控5.98E-0739ELF1,THRB,PDGFA,GLIS2等 GO:0045935核堿基、核苷,核苷酸和核酸代謝過程的正調控7.58E-0738ELF1,THRB,PDGFA,GLIS2等 GO:0010557高分子生物合成過程的正調控8.70E-0739ELF1,THRB,PDGFA,GLIS2等GO:0045893轉錄的正調控,DNA依賴9.37E-0732ELF1,THRB,PAX6,CASK等 GO:0031328細胞生物合成過程的正調控1.03E-0640ELF1,THRB,PDGFA,GLIS2等 GO:0010628基因表達的正調控1.07E-0636ELF1,THRB,GLIS2,PAX6等 GO:0051254RNA代謝過程的正調控1.13E-0632ELF1,THRB,PAX6,CASK等細胞組成(部分) GO:0012505內膜系統4.15E-0638SYT1,CLCN3,MAL2,XPO6等 GO:0031252細胞前沿7.91E-0614MTSS1,WASF2,ABI2,CDK6等 GO:0031982囊泡9.76E-0531SRI,MTSS1,SYT1,CLCN3等 GO:0031410細胞質囊泡1.13E-0430MTSS1,SYT1,CLCN3,YWHAZ等 GO:0005856細胞骨架1.24E-0451CTTNBP2NL,MTSS1,UTRN等 GO:0005794高爾基體2.24E-0436CLCN3,OSBP,AP1G1,UNC50等 GO:0042995細胞投射4.44E-0430MTSS1,SYT1,NRP1,CNN3等 GO:0031988膜結合囊泡5.02E-0426MTSS1,SYT1,CLCN3,YWHAZ等 GO:0001726胞膜褶皺5.47E-048MTSS1,NME2,NME1-NME2等 GO:0015629肌動蛋白細胞骨架6.76E-0416CTTNBP2NL,MTSS1,TWF1,UTRN等分子功能(部分) GO:0003700轉錄調節活性2.15E-0751ELF1,BACH2,THRB,E2F5等 GO:0030528DNA結合3.51E-0768HMGN1,ELF1,BACH2,THR等 GO:0003677轉錄阻遏物活性1.29E-0587HMGN1,ELF1,BACH2,E2F5等 GO:0016564序列特異性DNA結合7.45E-0521ZNF281,ELF1,CTBP2,THRB等 GO:0043565細胞骨架蛋白結合1.39E-0431ELF1,BACH2,THRB,PAX6等 GO:0008092肌動蛋白結合1.99E-0427MTSS1,CNN3,UTRN,WASF2等 GO:0003779轉錄調節活性3.26E-0420MTSS1,TWF1,CNN3,MAP1A等 GO:0016566特異性轉錄阻遏物活性1.57E-036HDAC4,TGIF1,PAX3,TCF7L2等 GO:0016563轉錄激活子活性2.10E-0321ZNF281,ELF1,GLIS2,MAML2等 GO:0003702RNA聚合酶II轉錄因子活性5.97E-0314ZNF281,HMGN1,LEF1,PAX3等
表3 miR-206靶基因KEGG通路富集分析結果

IDKEGG通路名稱基因數量P值參與基因hsa04360軸突導向131.11E-04NRP1,MET,EFNB2,MAPK1,KRAS,SEMA6D等hsa04520黏附連接101.47E-04ACTB,MAPK1,MET,WASF2,LEF1,WASL等hsa04320背腹軸形成53.59E-03NOTCH3,MAPK1,KRAS,ETS1,CPEB1hsa04310Wnt信號通路115.55E-03CTBP2,SFRP1,CCND2,PPP2R5A,NFAT5等hsa05216甲狀腺癌56.23E-03MAPK1,KRAS,LEF1,TCF7L2,TPM3hsa05211腎細胞癌79.19E-03MAPK1,KRAS,ETS1,PAK3,MET,VEGFA等hsa04810肌動蛋白細胞骨架調節139.50E-03GIT1,ACTB,MAPK1,PFN2,KRAS,ARPC3等hsa05130致病性大腸桿菌感染61.55E-02ACTB,YWHAZ,ARPC3,YWHAQ,WASL,NCLhsa04660T細胞受體82.15E-02MAPK1,KRAS,PAK3,CBL,NFAT5,NFATC2等hsa05200癌癥通路162.20E-02CTBP2,PDGFA,CBL,MET,IGF1,LEF1等

圖1 數據庫預測hsa-miR-206的靶基因個數
2.4miR-206預測靶基因所編碼蛋白質間的相互作用分析通過STRING在線工具篩選出靶基因蛋白互作網絡,導入Cytoscape,利用Mcode插件篩選出關鍵基因(Hub基因)共27個,見圖2。
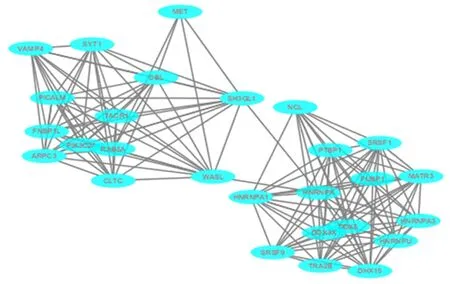
圖2 miR-206 預測靶基因所編碼蛋白質間的相互作用分析
3 討 論
miRNA是一類小分子非編碼內源性調節RNA,特能結合到靶mRNA的3’非翻譯區(Untranslated region,UTR)從而抑制翻譯或者直接降解靶mRNA,具有轉錄后水平的多基因調節功能。miRNA通過堿基互補配對原則特異性結合其靶基因,形成一個巨大的分子調控網絡,廣泛參與到人類生命活動和疾病的各個方面中去[13]。目前已經在自然界中發現了超過三萬種miRNA,參與到細胞的增殖、分化、凋亡及機體的免疫調節和發育等過程,近年來的研究發現miRNA在多種疾病中的表達紊亂[14]。深入研究miRNA與疾病的相關性將有望為疾病的臨床診斷及治療提供新思路和新策略。
隨著技術的進步,更多的技術被用來量化臨床標本的miRNA,如Northern blotting、基因芯片分析、高通量測序,實時定量PCR等[15-16]。然而,由于通過實驗途徑一一驗證miRNA的靶基因較為困難且繁瑣,故而篩選miRNA靶基因,預測其潛在生物學功能,對指導miRNA的實驗研究具有重要意義。
考慮到靶基因預測過程中miRNA與mRNA雙鏈特異結合的熱穩定性、miRNA與靶基因結合位點的序列匹配、序列的保守性等因素,本研究選用3種經典的miRNA靶基因預測工具Targetscan、miRDB和DIANA-microT,用不同的計算方法預測靶基因,取交集作為靶基因集合。miRbase是由曼徹斯特大學的研究人員開發的一個在線的miRNA數據庫,該數據庫中收錄了來自200多個物種,接近4萬個miRNA序列和注釋的最全面的miRNA數據庫,通過瀏覽和搜索等方式進行檢索,每個條目中包含了該miRNA前體莖環結構的序列,其經過切割后獲得的成熟miRNA的序列,以及它在其他物種間的分布情況等的基本信息。本文通過查找各個物種的miR-206與人類miR-206通過NCBI BLAST比對分析發現miR-206的成熟序列在各物種之間高度保守,提示其可能具有重要的生物學功能。利用DAVID在線分析工具進行GO分析和KEGG分析,GO分析從細胞定位、分子功能,生物學途徑分析靶基因負富集于那些生物學過程,KEGG分析結果表明靶基因參與哪些細胞代謝通路。通過GO分析我們得知miR-206主要參與生物合成調節、有機物代謝過程調節、轉錄調節等生物學過程,KEGG分析表明miR-206主要參與Wnt信號通路、T細胞受體信號通路、癌癥通路等。這與已有的研究及文獻報道的hsa-miR-206在多種疾病中已揭示的一些作用機制相一致,證明各生物信息學軟件的預測結果具有一定可靠性,能夠為對hsa-miR-206的進一步研究提供指導方向。隨后通過String11.0在線數據庫和Cytoscapev3.6.1軟件對420個差異靶基因進行互作預測分析發現,各靶標蛋白質之間存在復雜的相互作用,共篩選出27個關鍵基因,如DDX5、FUBP1、DHX15等,因此我們推測這些基因可能在miR-206調控的生物學效應中發揮重要作用。DDX5是在各種惡性腫瘤中過表達的ATP依賴性RNA解旋酶,越來越多的證據表明,DDX5通過促進細胞增殖和轉移而參與癌變和癌癥進展,研究顯示,DDX5通過激活mTOR/S6K1誘導胃癌細胞的生長。特異的mTOR抑制劑依維莫司的治療顯著減弱了DDX5介導的細胞增殖[17]。體外實驗表明,FUBP1通過增強糖酵解和ATP產生來促進神經母細胞瘤的細胞增殖并抑制細胞凋亡[18]。DHX15是DEAH-box RNA解旋酶家族的杰出成員,有研究表明[19],DHX15在肝細胞肝癌中顯著上調,其高表達與不良預后相關,提示其在肝細胞肝癌進程中的關鍵作用。
最近的一系列研究證實miR-206在多種人類疾病中具有重要的生物學功能,尤其集中于腫瘤相關疾病,許多研究者發現has-miR-206可在多種腫瘤組織與正常組織間有差異性表達。Zhou等[20]研究發現,與鄰近的正常組織相比,miR-206在乳腺癌細胞系和乳腺癌組織中的表達上調,而全長Neurokinin-1的表達與腫瘤淋巴結轉移(TNM)階段和淋巴結轉移呈負相關,miR-206結合全長神經激肽-1信使RNA的3'-非翻譯區,調節蛋白質表達。與正常鄰近組織相比,宮頸癌樣品中的miR-206表達明顯下調。多變量Cox回歸分析顯示,miR-206表達降低是整體生存的獨立不利預后因素。此外,miR-206模擬物在HeLa細胞中的轉染能夠減少細胞增殖,促進細胞凋亡并抑制細胞入侵和遷移[21]。Chang等[22]研究指出FOXD2-AS1通過充當miR-206海綿來上調miR-206靶基因膜聯蛋白A2(ANXA2)的表達。總之,結論是FOXD2-AS1在HCC中起癌基因的作用,并通過“刺激”miR-206部分上調ANXA2的表達。miR206可以通過調節REST/HDAC4/Sp1/Sp4/BDNF軸來調節MeHg誘導的神經細胞死亡[23]。在宮頸癌中,觀察到lncRNA HOTAIR抑制后的遷移和侵襲的表型效應,至少部分是通過抑制Hela細胞miR206表達來調節MKL1[24]。在肺鱗狀細胞癌中,miR206通過下調MET和EGFR的mRNA和蛋白水平來抑制EBC-1細胞增殖、遷移和侵襲,另外,ERK1/2和AKT信號轉導的磷酸化被癌細胞中的miR-206恢復所抑制[25]。
在動物實驗方面,關于miR-206的研究主要集中于miR-206經由TGF-β1/Smad3信號通路影響衛星細胞的增殖和分化來抵抗骨骼肌萎縮[26],miR-206通過靶向Notch3基因來調節骨骼肌細胞的增殖和細胞周期停滯[27],miR-206靶向的AAV有效地下調了miR-206的表達并增加了成熟小鼠肌肉中的內源性治療基因的表達,且治療顯著改善了小鼠的運動功能和營養不良[28]。而關于miR-206在腫瘤方面的動物模型目前仍較少,這一點可以為將來的研究方向作以參考。
盡管已經有了一些認識,但是我們對miR-206靶基因功能的認識還只是冰山一角,因為miRNAs在不同條件下調控不同細胞的不同靶點發揮不同的作用,這可能是因為同一miRNA能夠調控多個靶基因,信號轉導通路不同,還可能是因為生物功能的異質性、某種疾病的位置或階段不同等,從而導致了這種功能上的巨大差異[29-30]。本研究聯合采用TargetScan、miRDB、DIANA-microCT3個靶基因預測數據庫得到可信度較高的靶基因集合,并對靶基因進行GO功能注釋和KEGG信號轉導通路富集分析,并進行關鍵基因篩選,將有助于對miR-206所參與的生物學過程有一個基本的認識,為進一步深入研究其功能奠定了基礎。但由于預測靶基因過程中不可避免的存在假陽性率,所以對預測得到的靶基因及其發揮的生物學功能需要進行進一步的實驗驗證。
猜你喜歡
現代畜牧科技(2021年4期)2021-07-21 06:13:00
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
電子制作(2018年18期)2018-11-14 01:48:24
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
