NaCl 脅迫對黃瓜幼苗葉片CsPLDα和CsNMAPK 表達(dá)及信號分子的影響
2020-04-09 06:23:56姚佳麗李魯俊楊鳳娟王秀峰魏珉史慶華
姚佳麗,李魯俊,楊鳳娟,2,3*,王秀峰,2,魏珉,3,4,史慶華,3,4
1.山東農(nóng)業(yè)大學(xué)園藝科學(xué)與工程學(xué)院/作物生物學(xué)國家重點實驗室,山東 泰安 271018
2.農(nóng)業(yè)農(nóng)村部黃淮地區(qū)園藝作物生物學(xué)與種質(zhì)創(chuàng)制重點實驗室,山東 泰安 271018
3.山東果蔬優(yōu)質(zhì)高效生產(chǎn)協(xié)同創(chuàng)新中心,山東 泰安 271018
4.農(nóng)業(yè)農(nóng)村部黃淮海設(shè)施農(nóng)業(yè)工程科學(xué)觀測實驗站,山東 泰安 271018
黃瓜是設(shè)施栽培的主要蔬菜作物之一,在生產(chǎn)過程中經(jīng)常遇到鹽害等逆境脅迫。高濃度鹽漬化逆境主要通過滲透、離子和氧化脅迫使植物受到傷害[1-3],從而影響植物能量和物質(zhì)代謝[4],使植物生長受到抑制甚至導(dǎo)致死亡[5]。植物一般通過調(diào)節(jié)自身的形態(tài)特征與生理生化過程來適應(yīng)輕度鹽脅迫[6];而通過合成滲透調(diào)節(jié)物質(zhì)、區(qū)隔過量離子及清除活性氧等響應(yīng)來減輕高鹽環(huán)境造成的脅迫損害,這些響應(yīng)主要包括信號感知、信號轉(zhuǎn)導(dǎo)、基因表達(dá)及相應(yīng)產(chǎn)物產(chǎn)生等在內(nèi)的信號系統(tǒng)[7]。細(xì)胞膜是植物細(xì)胞感知并傳遞環(huán)境脅迫信號的最重要部位。鹽脅迫下,植物通過細(xì)胞膜將胞外脅迫信號傳遞至胞內(nèi),進而在胞內(nèi)引發(fā)一系列信號級聯(lián)反應(yīng);其自身也能作為前體參與第二信使1,4,5-三磷酸肌醇(IP3)、二脂酰甘油(DAG)、磷脂酸(PA)等合成,而參與磷脂信號通路[8]。
磷脂酶D(PLD)能夠水解膜磷脂產(chǎn)生第二信使分子PA,PA能激活促分裂蛋白激酶(mitogen-activated protein kinase,MAPK)信號流和離子通道活性等引發(fā)一系列次級反應(yīng),從而完成細(xì)胞響應(yīng)外部信號的過程[9,10]。在信號傳遞過程中,MAPK級聯(lián)信號系統(tǒng)起著承上啟下,將信號整合、放大和傳遞的作用[11]。林峰[12]研究發(fā)現(xiàn),AtPLDα1缺失使突變體對鹽更加敏感,而外施PA則可緩解鹽脅迫現(xiàn)象;Li等[13]研究發(fā)現(xiàn),將番茄SpMPK3轉(zhuǎn)入擬南芥中可顯著提高鹽脅迫和干旱脅迫下種子的發(fā)芽率和抗氧化能力,表明SpMPK3是響應(yīng)擬南芥萌發(fā)和發(fā)育階段干旱和鹽脅迫的正調(diào)控因子。劉文瑜等[14]研究發(fā)現(xiàn)低濃度的NO可激活抗氧化系統(tǒng)酶活性以減輕鹽脅迫對蒺藜苜蓿種子的傷害。鹽脅迫抑制黃瓜幼苗植株的生長及光合電子傳遞、降低葉片凈光合速率[15],嚴(yán)重加劇膜脂過氧化程度[16]。Yu等[17]和莊寶程[18]研究發(fā)現(xiàn),PA能與MAPK級聯(lián)信號中的成分特異性結(jié)合,表明PLD信號轉(zhuǎn)導(dǎo)途徑與MAPK級聯(lián)信號轉(zhuǎn)導(dǎo)途徑在響應(yīng)逆境信號中存在關(guān)聯(lián)性。故本試驗擬通過NaCl處理模擬鹽害,測定處理前0 h和處理后0.5 h、1 h、3 h、5 h和10 h黃瓜幼苗葉片中磷脂酶Dα(CsPL Dα)和促分裂原活化蛋白激酶(CsNMAPK)基因表達(dá)、PA、NO和活性氧等信號物質(zhì)的變化,以期探明CsPLDα和CsNMAPK及各種信號分子對鹽脅迫的響應(yīng)機制,為豐富黃瓜耐鹽機理提供理論依據(jù)。
1 材料與方法
1.1 材料與設(shè)計
以‘新泰密刺’黃瓜品種為試材。試驗于2017 年4~6 月在山東農(nóng)業(yè)大學(xué)本部玻璃溫室內(nèi)進行。經(jīng)浸種催芽后的種子播于裝有基質(zhì)(草炭:蛭石:珍珠巖=2:1:1)的穴盤中,待第1 片真葉展平時,將幼苗轉(zhuǎn)入盛5 L 營養(yǎng)液的水培盆中,每盆6 株。營養(yǎng)液中大量元素參照山崎配方[19]略加修改,微量元素參照Arnon 配方,pH 值用H2SO4調(diào)節(jié)在6.0~6.5 之間。待幼苗長至3 葉1 心時,一次性使?fàn)I養(yǎng)液中的NaCl 濃度達(dá)100 mM,分別在處理0 h、0.5 h、1 h、3 h、5 h 和10 h 后取樣并測定相關(guān)指標(biāo),重復(fù)3 次。
1.2 測定項目與方法
CsNMAPK和CsPLDα的mRNA 表達(dá)量參照Li 等[20]、Livak 和Schmittgen[21]的方法,通過實時熒光定量PCR 進行測定。CsNMAPK和CsPLDα的蛋白表達(dá)量通過Western blot 進行測定,所用抗體由艾比瑪特生物醫(yī)藥(上海)有限公司制備。PA 含量采用雙抗體夾心法測定,提取參照陶銀華[22]的方法并加以修改。NO 含量采用蘇州科銘生物技術(shù)有限公司一氧化氮含量測定試劑盒進行測定。O2.-的組織化學(xué)染色參考Jabs 等[23]的方法。H2O2染色用3,3’-diaminobenzidine(DAB),參考Veljovic-Jovanovic 等[24]的方法。
超氧化物歧化酶(SOD)活性測定參照Prochazkova 等[25]方法;過氧化氫酶(CAT)和過氧化物酶(POD)活性測定參照Cakmak 等[26]方法;抗壞血酸過氧化物酶(APX)活性測定參照Nakano 和Asada[27]的方法。
1.3 數(shù)據(jù)處理與分析
采用Excel 2010 軟件處理數(shù)據(jù)和繪圖,采用DPS V14.10 軟件進行統(tǒng)計、相關(guān)性分析,采用新復(fù)極差法進行差異顯著性檢驗(P<0.05)。
2 結(jié)果與分析
2.1 NaCl 脅迫對黃瓜幼苗葉片中CsPLDα和CsNMAPK 表達(dá)量的影響
圖1 所示,隨NaCl 處理時間延長,黃瓜幼苗葉片中CsPLDα和CsNMAPK基本呈先升高后降低趨勢。處理1 h 后CsPLDα表達(dá)量達(dá)最大值,為0 h 表達(dá)量的6.65 倍,差異顯著;而處理3 h 后CsNMAPK表達(dá)量達(dá)最大值,為0 h 表達(dá)量的15.56 倍,差異顯著,且兩基因表達(dá)量均在處理10 h 后略有上升。
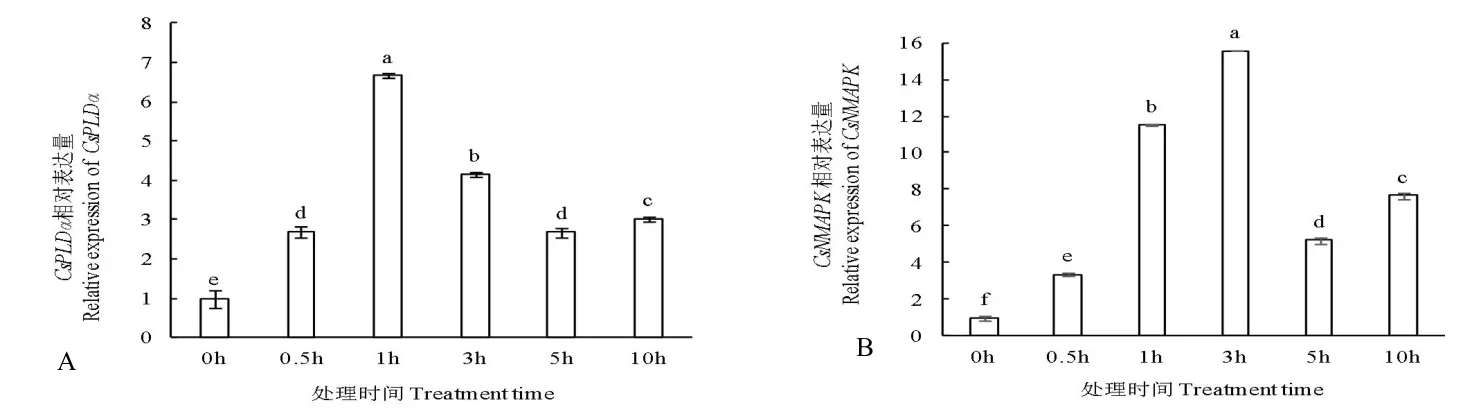
圖1 NaCl 脅迫對黃瓜幼苗葉片中CsPLDα(A)和CsNMAPK(B)相對表達(dá)量的影響Fig.1 Effects of NaCl stress on relative expression of CsPLDα(A)and CsNMAPK(B)in cucumber seedling leaves
2.2 NaCl 脅迫對黃瓜幼苗葉片中CsPLDα和CsNMAPK 蛋白表達(dá)量的影響
NaCl 處理后0~10 h 范圍內(nèi),黃瓜幼苗葉片中CsPLDα蛋白表達(dá)量均得到明顯誘導(dǎo),在處理1 h后表達(dá)量較高,處理5 h后的表達(dá)量亦稍高于其它處理。隨NaCl處理時間延長,幼苗葉片中CsNMAPK表達(dá)量呈先增后降趨勢,處理3 h 后表達(dá)量較高,其它處理時間下表達(dá)量差異不明顯(圖2)。
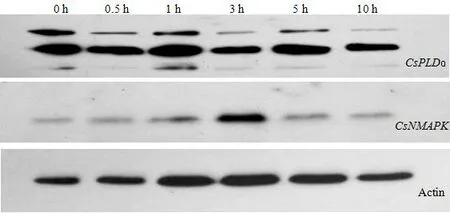
圖2 NaCl 脅迫對黃瓜幼苗葉片中CsPLDα和CsNMAPK 蛋白表達(dá)量的影響Fig.2 Effects of NaCl stress on CsPLDα and CsNMAPK protein expression in cucumber seedling leaves
2.3 NaCl 脅迫對黃瓜幼苗葉片中磷脂酸含量的影響
由圖3A 可以看出,隨處理時間延長,黃瓜幼苗葉片中PA 含量亦呈先升高后降低趨勢。在NaCl處理3 h 后,PA 含量達(dá)到較高值,比處理0 h 顯著增加18.6%,隨后其含量逐漸降低。
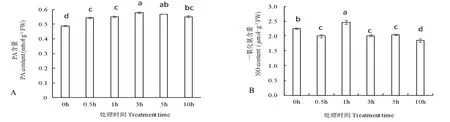
圖3 NaCl 脅迫對黃瓜幼苗葉片中磷脂酸(A)和一氧化氮(B)含量的影響Fig.3 Effects of NaCl stress on the content of PA(A)and NO(B)in cucumber seedling leaves
2.4 NaCl 脅迫對黃瓜幼苗葉片中一氧化氮含量的影響
由圖3B 可知,NaCl 處理0~10 h 范圍內(nèi),黃瓜幼苗葉片中一氧化氮(NO)含量在處理1 h 后顯著升高,比處理0 h 增加9.1%,隨后逐漸下降,且其它處理時間,其含量均低于0 h 處理。
2.5 NaCl 脅迫對黃瓜幼苗葉片中過氧化氫和超氧陰離子含量的影響
圖4 顯示,隨處理時間延長,黃瓜幼苗葉片中過氧化氫(H2O2)和超氧陰離子(O2·-)含量均呈逐漸升高趨勢。

圖4 NaCl 脅迫對黃瓜幼苗葉片中過氧化氫(A)和超氧陰離子(B)含量的影響Fig.4 Effects of NaCl stress on the content of H2O2和O2·in cucumber seedling leaves
2.6 NaCl 脅迫對黃瓜幼苗葉片中抗氧化酶活性的影響
如圖5 所示,隨處理時間延長,黃瓜幼苗葉片中抗氧化酶活性基本呈先降低后升高再降低趨勢,在處理0.5 h 后,其SOD、CAT、POD 和APX 活性均顯著低于0 h。SOD 活性在處理3 h 后達(dá)到較高值,但與處理0 h 無顯著差異,處理0.5 h、1 h、5 h 和10 h 后其活性均顯著低于處理0 h 后的活性;CAT、POD 和APX 活性均在處理5 h 后達(dá)較高值,分別比處理0 h 增加31.3%、33.0%和15.6%,在處理10 h 后略有降低。

圖5 NaCl 脅迫對黃瓜幼苗葉片中超氧化物歧化酶(A)、過氧化氫酶(B)、過氧化物酶(C)和抗壞血酸過氧化物酶(D)活性的影響Fig.5 Effects of NaCl stress on the activities of SOD(A),CAT(B),POD(C)and APX(D)in cucumber seedling leaves
3 討論
研究表明,植物能夠感受并迅速應(yīng)答各種脅迫的信號轉(zhuǎn)導(dǎo)過程,與特異的磷脂酶D(PLD)和下游產(chǎn)物磷脂酸(PA)激活有關(guān)[28];PA 是植物中重要的脂質(zhì)信號分子,被稱為“脂質(zhì)第二信使”,參與多種逆境脅迫相關(guān)的信號傳導(dǎo)途徑[29]。而許多信號途徑依靠支架蛋白提供的定位點,與信號通路中的成員結(jié)合進而有效傳遞信號,特別是MAPK 級聯(lián)途徑。當(dāng)用NaCl 處理擬南芥細(xì)胞或幼苗時,AtPLDα1可被激活,產(chǎn)生第二信使PA 來參與鹽響應(yīng)過程[17,30]。林峰[12]研究表明,外施PA 可緩解植物因缺失AtPLDα1對鹽的敏感性。Yu 等[17]研究發(fā)現(xiàn),鹽脅迫激活的PLDα1所產(chǎn)生的PA,能誘導(dǎo)MAPK6 激酶活性增加,進而引起下游機制響應(yīng)來提高植物抗鹽性;Lee[8]研究發(fā)現(xiàn)用正丁醇抑制PLD活性后,MAPK 活性亦受到抑制,而外施PA 能激活MAPK 活性。本試驗結(jié)果表明,短時間NaCl脅迫處理后,黃瓜幼苗葉片中CsPLDα和CsNMAPK表達(dá)量呈先升高后降低趨勢,CsPLDα表達(dá)量在處理1 h 后達(dá)最大值,而CsNMAPK表達(dá)量在處理3 h 后達(dá)最大值;兩者的蛋白表達(dá)量亦分別在處理1 h 和3 h 達(dá)最大值;PA 含量亦呈先升高后降低趨勢,在處理3 h 后達(dá)較高值,Li 等[31]研究結(jié)果與之相一致。以上結(jié)果說明,鹽脅迫下黃瓜幼苗葉片中CsPLDα和CsNMAPK基因?qū)aCl 脅迫響應(yīng)存在時間差,且CsPLDα響應(yīng)早于CsNMAPK。NO 作為一種氣體生物活性分子,在植物的生長發(fā)育和逆境信號應(yīng)答過程中發(fā)揮重要功能[32]。Zhang 等[33]研究發(fā)現(xiàn)PLD 和PA 能夠參與NO 緩解鹽脅迫信號轉(zhuǎn)導(dǎo)過程;Zhang 等[34]研究表明ABA 誘導(dǎo)氣孔關(guān)閉信號轉(zhuǎn)導(dǎo)過程中,NO 位于PLDα1/PA 信號下游;同時發(fā)現(xiàn)NO 能夠激活葉肉細(xì)胞的PLD 活性。Raho 等[35]研究發(fā)現(xiàn),植物通過PLD 和PLC/DGK 途徑產(chǎn)生PA 需要NO 參與。Kumar 等[36]和Clark 等[37]研究發(fā)現(xiàn)激活的MAPKs 級聯(lián)途徑可引發(fā)NO 大量產(chǎn)生,而NO 亦能激活MAPK 活性來提高煙草和擬南芥的抗病能力。本試驗結(jié)果表明,NO 可迅速被NaCl 誘發(fā),且在處理1 h 后黃瓜幼苗葉片中NO 達(dá)最大值,與CsPLDα表達(dá)量出現(xiàn)最大值同步,且早于CsNMAPK信號響應(yīng)。
鹽脅迫下,植物體內(nèi)會產(chǎn)生并積累大量的氧自由基,從而引起細(xì)胞膜脂過氧化。作為植物抗氧化的第一道防線,SOD 可通過清除過剩的超氧陰離子,催化活性氧發(fā)生歧化反應(yīng)產(chǎn)生H2O2,來提高植物的抗氧化能力[38];而POD 和CAT 能催化H2O2與其底物反應(yīng)進而分解H2O2,從而解除逆境脅迫產(chǎn)生的活性氧對植物的傷害[39]。本試驗結(jié)果表明,NaCl 脅迫下,黃瓜幼苗葉片中SOD 活性在3 h后達(dá)較高值,CAT、POD 和APX 活性均在5 h 后達(dá)較高值;而葉片中H2O2和O2-·含量卻隨處理時間延長呈逐漸增加趨勢。Ji 等[40]研究表明,過表達(dá)黃瓜CsPLDα可通過提高抗氧化酶SOD、POD、CAT及APX 的活性來提高煙草幼苗對NaCl 脅迫的抗性。而王靜[41]研究也發(fā)現(xiàn),過表達(dá)CsNMAPK能通過增加黃瓜幼苗葉片中的NO 含量和減少葉片中的O2·-和H2O2含量來提高對高濃度NO3-的抗性。說明以上保護機制可能是由受CsPLDα介導(dǎo)的CsNMAPK級聯(lián)途徑來激活[42]。
4 結(jié)論
本試驗條件下,經(jīng)100 mM NaCl 處理后,黃瓜幼苗葉片中CsPLDα和CsNMAPK 基因表達(dá)量和蛋白表達(dá)量分別在處理1 h 和3 h 達(dá)最大值,說明此兩基因表達(dá)存在時間差;PA 含量呈先升高后降低趨勢,在處理3 h 后達(dá)較高值;NO 含量在處理1 h 顯著升高;H2O2和O2-含量隨處理時間延長呈逐漸增加趨勢;SOD 活性在處理3 h 后達(dá)較高值,而CAT、POD 和APX 活性均在處理5 h 后達(dá)較高值。以上結(jié)果表明,黃瓜幼苗葉片中CsPLDα和CsNMAPK基因表達(dá)對鹽響應(yīng)存在時間差,且CsPLDα表達(dá)早于CsNMAPK表達(dá)。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(2015年3期)2015-11-11 17:20:00
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
