植物功能多樣性及其在苔蘚植物中應用的研究
2020-04-09 06:21:00張永昌郭水良
上海師范大學學報·自然科學版 2020年1期
張永昌,何 林,郭水良*
(1.上海師范大學生命科學學院,上海200234;2.遵義師范學院生物與農業科技學院,貴州遵義563006)
苔蘚植物是一類由水生向陸生過渡的、特殊的高等植物類群.由于苔蘚植物對環境特別敏感,對水分和礦質營養有特殊的吸收方式,能夠利用孢子進行遠距離傳播,并且有多種營養繁殖方式,很多苔蘚植物耐旱能力強,使其能廣泛分布于海洋以外的地球絕大部分生境中[1-5],是在種數上僅次于被子植物的高等植物類群.全世界苔蘚植物約有23 000種[5],中國有150科591屬3 021種,其中,蘚類有86科431屬1 945種,苔類有60科152屬1 050種,角苔類有4科8屬26種[6],是世界上苔蘚植物多樣性程度最高的國家之一.
苔蘚植物具有眾多的生物多樣性價值,是溫帶和亞高山生態系統的主要組成成分,在水分涵養、物質循環、氣候調節等方面發揮著重要作用,同時在環境監測、園林景觀、藥物開發等方面也至關重要.苔蘚植物在生態系統的生態功能以及功能多樣性,已經成為苔蘚植物生態學研究的熱點之一.
功能多樣性最初定義為影響群落中生態系統功能的所有物種及有機物的功能性狀值及其變動范圍,強調的是性狀的差異性[7].因其與生態系統功能關系緊密,功能多樣性研究日益受到人們的重視[8].目前,國內外對植物功能多樣性的研究主要集中在維管植物,已經比較成熟,研究領域包括:植物功能性狀沿環境梯度的變化規律(與氣候、海拔、地形等的關系)[9-16];植物功能性狀對干擾(包括土地利用變化、火燒、放牧等)的響應[17-18];植物功能多樣性與物種多樣性或生態系統功能之間的關系[19-22].
PARDOW 等[23]基于苔蘚植物生活型研究了法屬圭亞那熱帶雨林樹附生苔蘚植物的功能多樣性,以探究該地區苔蘚植物多樣性的熱點區域.BERDUGO 等[24]在美國Whiteface 山的云杉林帶研究了林窗處苔蘚植物物種多樣性和功能多樣性,并創新性地使用苔蘚植物的形態、生態和生殖的多種性狀組合,研究了沿自然冠層開放梯度的苔蘚植物功能差異.功能多樣性已繼物種多樣性和系統發育多樣性之后逐步成為研究苔蘚植物多樣性的又一重要指標,對開展苔蘚植物多樣性的研究和保護具有重要的意義.
基于此,本文作者對功能多樣性在植物類群的研究進展,及苔蘚植物功能多樣性的研究成果進行了綜述,并展望了未來苔蘚植物功能多樣性研究的方向.
1 植物功能多樣性概念
相對于物種多樣性,功能多樣性更能反映植物對環境驅動力的適應以及植物內不同功能之間的權衡關系,也更能體現植物在生態系統中的作用.目前,關于植物功能多樣性的研究涉及從低等植物到高等植物,從水生植物到陸生植物,從個體到群落等多個層次[14,25-28],但植物功能多樣性的概念至今還沒有標準的定義[29-31].TILMAN[7]將功能多樣性定義為能夠影響生態系統運作或行使功能的生物多樣性的成分,但很難據此含義對功能多樣性進行定量化的描述[30].目前普遍為人們所接受的功能多樣性定義是指某一群落內物種間功能性狀的變化范圍或指特定生態系統中所有物種功能性狀的數值和范圍[31].
2 功能性狀的選擇
植物功能性狀指影響植物生長、繁殖和生存的一系列植物屬性[32],在種子植物中包括了植株高度、生活型、種子干重、比葉面積、葉形、葉片氮含量等,人們常根據研究問題的需要和植物類群選擇不同的功能性狀,如在討論水分、溫度對植物的影響時,會選擇與蒸騰作用相關的比葉面積.在維管植物中,研究者常在一個或多個生態系統中,關注環境梯度與植物功能性狀變化間的關系,探討植物對環境變化的響應[15,33-34].與植物建立、生長、繁殖、傳播和維持有關的性狀也受到重視,這些性狀包括葉片(比葉面積、葉片形狀、葉片氮含量、葉片磷含量等)、植株(生長形態、株高等)、繁殖器官(花期長度、果實類型和成熟期、種子干重等),探討這些性狀在不同生境中的變異規律[11,14,16,35-38].苔蘚植物生活史特殊,配子體發達,沒有維管組織,又是變水植物,對環境特別是小生境變化非常敏感,在營養吸收和水分傳導方面有自己特殊的方式,從而表現出與其他高等植物明顯不同的生態適應特征.因此,在苔蘚植物中開展功能多樣性研究,人們常選擇與水分、營養和光合生理關系密切的生活型,與環境水分利用有關的葉片細胞特征(疣、中肋、莖附屬物等),和繁殖有關的孢子體特征(蒴齒長短、孢子大小等)[23,39-41].
3 功能多樣性指標及其計算方法
目前還沒有被普遍接受的功能多樣性測度方法,研究者常根據研究的實際情況選擇某種功能多樣性指數來反映植物功能性狀的變化[42-43].
MASON 等[44]依據物種多樣性的組成,把功能多樣性劃分為功能豐富度(functional richness)、功能均勻度(functional evenness)和功能離散度(functional divergence).SCHLEUTER 等[31]在前人的基礎上,對植物能多樣性指數做了進一步的研究,通過對不同功能多樣性指數的比較和分析,闡明了不同功能多樣性指數的特征及其與其他指數的相關性及缺點,并基于此總結發展了新的功能多樣性指數的計算方法.MCPHERSON 等[42]用R 語言編制了功能冗余指數的計算程序,并認為前人提出的功能多樣性指標不能很好地反映性狀數量、性狀值性質、性狀分布或豐度頻率敏感性等,對這些敏感性的更好理解能使研究者將多樣性指數與生態群落的基本特性聯系起來,從而有助于選擇最適合某一特定研究目的的指數.宋彥濤等[45]和陳又清[8]已對植物功能多樣性及其運算方法做了相關的綜述,在這里本文作者僅對常用的功能多樣性指數及運算方法進行簡要的介紹.
3.1 功能豐富度
功能豐富度是指物種或個體在群落中所占空間或功能范圍的大小[42],是體現生態空間利用程度的指數.該指數越大,生態空間利用程度越高[46],用來計算某一群落該性狀生態位空間占全部群落中該性狀生態位空間的比例[44]:
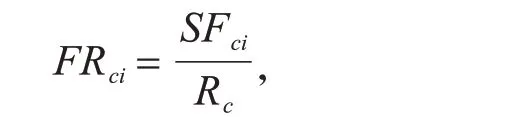
其中,FRci為在群落i 中性狀c 的功能豐富度;SFci為群落中物種所占據的生態位空間;Rc為所有群落中性狀c占據的生態位空間.
3.2 功能均勻度
功能均勻度指數(FE0)用來表征物種性狀在所占據性狀空間的分布規律,該指數越大,說明物種分布得越規律,對有效資源的利用越全面[44].
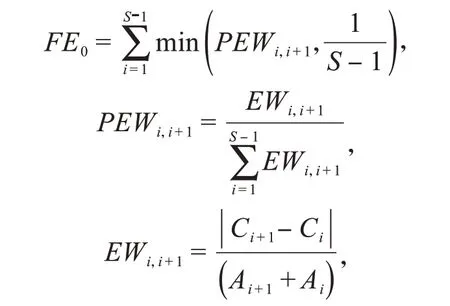
其中,S 為物種數;PEWi'i+1為物種i和i+1 性狀值權重差異的百分比;Ai為物種i的相對多度;Ci為物種i的性狀值.
3.3 功能離散度
功能離散度指數(FDS)表示群落內有機體性狀值的差異性,體現了群落內有機體的生態位分化和資源競爭程度,該指數越高,說明群落內有機體生態位互補程度越強,競爭越弱[44].
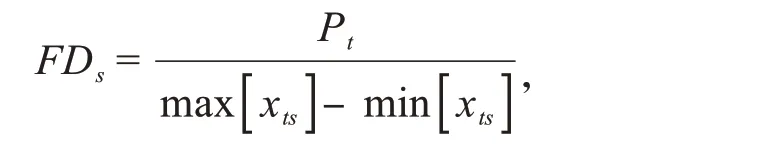
其中,Pt=Q1(Yt)-Q3(Yt);Yt為物種s 的性狀平均值重復次數,Yt=[(Xts,Xts,…,Xts)×As];Xts為群落中每個物種的性狀平均值;As為群落中物種s的多度;Q1和Q3分別為Yt的下四分位數和上四分位數.
在進行功能多樣性指數計算時,推薦使用R語言中的“FD”Package和FDiversity軟件進行計算.
4 維管植物功能多樣性研究現狀
理論上,植物周圍的種種環境因子都會在不同程度上影響其生長發育.在功能多樣性研究中比較關注氣候(溫度、水分、光照等)、土壤(pH 值、含氮量、含磷量等)、地形地貌(海拔、坡度、坡向等)、干擾(放牧、土地利用等)等因素.
(1)水作為植物生長的必需條件,對植物功能多樣性有重要的影響.ALBRECHT 等[16]在Kilimanjaro山寬廣的氣候和土地利用梯度上,分析了200種植物花和果實性狀的變化規律,發現研究區域植物的功能多樣性主要受降水的限制.SAIZ等[15]以南極洲以外其他大陸的100種旱地植物為研究對象,通過對植物空間分布、株高和葉面積指數在環境梯度上的變化式樣分析,發現降水量的季節性變化是影響群落功能多樣性的主要因素.NUNES 等[11]以地中海旱地生態系統的橡樹林地為研究對象,分析了13 個與植物建立、生長、繁殖、傳播和種群維持有關的性狀,發現功能離散度隨著干旱程度增加而降低,從而增加了干旱區種間的功能相似性.ORDONEZ 等[35]以在廣泛分布于歐洲的6 個被子植物類群為對象,估測了植物功能豐富度和離散度的地理格局,比較了葉面積指數、最大莖高、種子干重、莖比密度、生長型等5個功能性狀,根據將第四紀冰川期—間冰期氣候變化和當代環境因素(氣候、生產力和地形)作為評價功能多樣性決定因素的相對重要性,發現氣候變化可能會導致植物功能多樣性的長期失衡.
(2)土壤狀況也與植物的功能多樣性密切相關.ZHU 等[9]以中國黃土高原的16 個半干旱恢復草地為區域,研究了植被群落功能組成對土壤侵蝕的影響,定量計測了對恢復草地群落內13 個優勢植物中的9 個可能與土壤侵蝕相關的功能性狀,發現功能離散度與土壤侵蝕程度之間存在著較強的負相關關系,表明植物功能性狀多樣性的提高會減少該地區土壤侵蝕.ZUO 等[36]以半干旱草地的植物株高、比葉面積、比葉重、葉干重等性狀值為依據,研究放牧和土壤性質誘導植物功能多樣性對地上生物量和地下生物量的影響,發現放牧和土壤性質引起的生物多樣性變化對草地生態系統功能的影響強烈依賴于植物功能多樣性.張雪妮等[12]研究了土壤水鹽變化對艾比湖荒漠植物群落水平的功能性狀的影響,涉及植物形態、生理和化學等8 個功能性狀,并從多維性狀和一維性狀角度進行分析,發現不同生活型植物間的功能性狀在高水鹽環境下差異顯著,而在低水鹽環境下趨于相似,揭示了功能多樣性對土壤水分和鹽分變化的響應規律.ZHANG 等[47]在研究廢棄農田內植物的功能群組成、物種多樣性與土壤性狀的關系時,也得出了相似的結論.
(3)經緯度、海拔、坡向等地理因素通過影響水分、溫度等間接地影響植物功能多樣性,這種影響有明顯的尺度效應[10].STEINERT 等[48]研究了挪威東南部蟲煤植物群落的物種多樣性和功能多樣性,基于植物9 個花部特征計算了功能多樣性指數,發現海拔會影響物種豐富度和功能均勻度.LIU 等[13]比較了29 個亞熱帶湖泊水生植物的物種豐富度和功能多樣性,計測了物種豐富度和代表水生植物營養、再生和生態特征的13個功能性狀,發現與低海拔區域湖泊相比,高原湖泊的功能豐富度較低,但功能均勻度和分散度較高,功能豐富度與環境變量之間無顯著相關性,表明功能多樣性對環境變量的抗性較強.ZHANG 等[10]研究了高原高寒草甸植物功能群組成、物種多樣性與土壤性質(pH值、土壤總氮、土壤含水量)的關系,結果表明,植物群落特征和土壤理化性質在不同坡度水平上存在顯著差異,坡面通過影響水分等生態因子和土壤理化特性,直接或間接地影響高寒草甸植物的物種組成和功能群差異.朱云云等[49]以黃土高原不同坡向上的自然草地群落為對象,選擇了能很好反映植物競爭強度、響應環境變化的兩個功能性狀指標——株高和比葉面積,比較研究了植物功能性狀和功能多樣性指數隨坡向的變化規律,通過比較黃土高原馬欄地區山地的各坡向植物功能性狀和3 種功能多樣性測度指標的變化,發現陰坡和半陰坡的植物性狀占據了更多的生態位空間,資源利用較充分且競爭較低,并且出現了高度的生態位分化;而陽坡和半陽坡由于水分等條件的限制,植物性狀可占據的生態位空間有限,且在占據的生態位空間資源使用不足,導致生態位分化程度較低,資源競爭比較強烈,即陰坡對草地群落功能多樣性的影響更高.
(4)干擾也會強烈地影響植物的功能多樣性.研究者們先后研究了放牧、刈割、施肥等與植物物種多樣性和功能多樣性的關系[14,17-18,50].FORD 等[14]以英國威爾士西北部的兩個原始自然林地和兩個放牧林地為對象,用植物功能性狀變化探索放牧強度和植物功能多樣性間的聯系,將比葉面積、葉片干重(LDMC)與植物耐受性指數(遮蔭性、耐旱性和耐澇性)相結合,計算了群落加權平均值(CWM)和功能多樣性;結合LDMC、平均株高、生長形態、光照、水分指標,計算了林下植物群落的加權平均值和功能多樣性指數,發現與放牧林地相比,未放牧的林地其幼苗的建立,幼樹的補充,和樹級的功能多樣性更強,表明控制放牧強度有利于林地生態系統恢復.YANG 等[17]在海口市內選擇了461 個樣地,基于花粉、種子、葉的相關性狀,研究了土壤利用類型對植物功能多樣性的影響,發現土地利用遺留效應對木本植物在分類和功能多樣性上存在顯著差異,其中以居住區的土地利用遺留效應最為明顯,建議在快速城市化地區的保護項目中更多地關注居住區的植被,特別是低實用價值的物種.石明明等[18]以青藏高原高寒草甸中的17種共有植物為對象,利用比葉面積、株高、葉干物質含量、葉分配4種植物功能性狀,分析了圍封與放牧處理對植物功能性狀和功能多樣性的影響,發現不同植物功能性狀對圍封與放牧處理響應存在差異,放牧降低了比葉面積和植物高度,而葉干物質含量和葉分配在處理間無顯著差別,反映了植物對資源獲取和利用存在權衡.放牧在總體上降低了植物功能性狀(比葉面積、葉干物質含量、葉分配、植株高度)的種間平均差異及分離度,反映了植物對光資源的競爭減弱.該結果與孔彬彬等[50]、陳超等[51]等關于刈割對高寒草甸物種多樣性和功能多樣性關系的影響的實驗結果一致,放牧可能導致植物對地上資源的競爭轉變為對地下資源的競爭.因此,在高寒草甸地區,今后關于植物功能性狀多樣性的研究應加強地下過程相關性狀的研究.WANG 等[52]、DICKIE 等[53]、ZUIDAM 等[54]分別闡述了入侵種或引進外來種對本地植物功能多樣性的影響.UYTTENBROECK 等[55]基于生態位,發現食草動物取食行為會顯著地影響生態系統的植物功能多樣性.
5 苔蘚植物功能多樣性研究現狀
功能性狀的選擇是功能多樣性研究的基礎.已有部分學者開展了苔蘚植物功能性狀的研究.例如,葉吉等[56]通過對長白山北坡暗針葉林地面和樹附生苔蘚植物生物量的研究,發現苔蘚植物生物量及物種組成能夠很好地指示長白山北坡林型,生物量能夠很好地指示苔蘚植物的生態功能;楊武等[57]對源于不同生境的17 種蘚類植物葉片進行了比較解剖,發現蘚類植物中肋導水主細胞的有無、厚壁細胞是否分化、中肋細胞層數和密度、葉片細胞層數和密度、葉表附屬物等差異顯著.WANG等[58]比較了川西老齡冷杉林下28 種苔蘚植物的生長形態以及與光合作用相關的生理指標,發現12 種直立型和16 種平伏型苔蘚單位質量的氮與最大光合能力回歸關系的斜率相同,但前者將更多的氮用于合成光合色素,而后者的形態結構更有利于捕獲光能;同時發現苔蘚植物生長形態對其營養分配和光合效率有很強影響.SIERRA 等[59]以苔蘚植物生殖性狀為指標,預測葉附生苔蘚植物群落的形成時間.WANG 等[60]研究了西藏干旱/半干旱地區苔蘚植物形態特征及其對環境變化的響應,發現苔蘚植物的形態特征與環境變化密切相關,不同基質環境變化對苔蘚植物的影響差異較大,苔蘚植物的各種功能性狀密切相關,這些性狀的組合對環境變化有協同反應.
近年來,苔蘚植物群落的功能多樣性也開始受到關注,應用的功能特征有配子體形態[39]、生活型[23],以及與水分吸收保持有關的性狀[40]等.AH-PENG 等[39]應用苔蘚植物形態特征,發現La Réunion 島亞高山苔蘚植物的功能多樣性與物種多樣性存在正線性相關,地表分布的苔蘚植物群落具有更高的功能冗余,比樹附生苔蘚植物群落具有更強的抗干擾能力.基于與水分獲取和保持有關的功能性狀,HENRIQUES 等[40]研究了Azorean 苔蘚植物功能多樣性和種類構成特點,發現沿海和山地苔蘚植物在功能性狀構成上明顯不同,氣候變化不僅會影響苔蘚植物種類的組成,也會影響苔蘚植物的功能多樣性.PARDOW 等[23]基于生活型研究了法屬圭亞那熱帶雨林樹附生苔蘚植物的功能多樣性,發現低地雨林是該研究地區的苔蘚植物多樣性熱點區域.LISBOA等[41]研究發現隨著海拔的上升,蘚類植物孢子變大,葉片變長,高度增加,功能多樣性提高.BERDUGO 等[24]在美國Whiteface山的云杉林帶,創新性地嘗試使用苔蘚植物的形態、生態,以及生殖的多種性狀組合,研究了沿自然冠層開放梯度的苔蘚植物的功能差異,發現林窗大小與物種多樣性之間不存在顯著相關性,而指示種和植物功能性狀可以用來評價沿環境梯度的苔蘚植物群落生態的變化.
6 苔蘚植物功能多樣性研究展望
目前尚未檢索到國內關于苔蘚植物在不同生態系統中功能多樣性研究工作的報道,這使得我國的苔蘚植物多樣性保護策略僅立足于物種數量狀況,而沒有全面考慮物種的功能性狀,不能夠全面地認識苔蘚植物在生態系統中的作用和價值.因此,在國內開展苔蘚植物功能多樣性研究對于國內苔蘚植物多樣性的保護意義重大.
本文作者認為,在進行苔蘚植物功能多樣性研究時,可以根據苔蘚植物的特點,將其功能特征分為以下4個方面:
(1)苔蘚植物的生活型與水分、營養及光合生理關系密切,也影響著環境的濕度等,因此是重要的功能性狀.常見苔蘚的生活型主要有叢集型(Turf)(如金發蘚、真蘚等)、墊狀(Cushion)(如紫萼蘚等)、交織型(Weft)(如青蘚、灰蘚、羽蘚等)、樹狀(Dendroids)(如樹蘚、萬年蘚等)、懸掛型(Pendant)等.另外,由于水分條件變異較大,有些適應能力較強、分布較廣的種類(如木蘚)在不同環境中發育生長成為不同的生活型;還需考慮植株密度、枝葉排列等指標.
(2)苔蘚植物的繁殖特征,包括:有性繁殖(有無孢蒴)、無性繁殖(有無芽胞等營養繁殖結構)、有無蒴齒、蒴齒長短(與孢子傳播有關)、孢子大小、蒴柄長短、苞葉大小、是否隱蒴等.
(3)樣地中苔蘚植物群落的蓋度、高度是與生物量相關的指標,也具有重要的生態功能和生物多樣性價值.
(4)苔蘚植物的形態特征,主要包括:莖中央是否有導水細胞分化,是否有儲水細胞,葉片先端是否有透明毛尖,葉片細胞是否具疣及發育程度,葉片細胞大小,葉片細胞是否多層,葉片干燥時是否強烈背卷,葉片干燥時是否強烈扭折卷縮,葉表是否有櫛片,有無葉片中肋,莖上是否有附屬物等,這些性狀是否與環境水分利用、截留和保持有關.
在亞高山苔蘚林、云冷杉林等寒溫帶森林生態系統,泥炭蘚沼澤濕地等生態系統中,苔蘚植物是重要的組成部分,今后應該選擇具有代表性的生態系統開展苔蘚植物功能多樣性、物種多樣性、系統發育多樣性間關系的研究,還應該研究苔蘚植物功能多樣性對維持健康的生態系統的作用.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
