RCP8.5情景下氣候變化對四川省獼猴桃潰瘍病病菌地理分布的影響
2020-04-10 06:57:04林姍陸興利王茹琳
江蘇農業科學 2020年3期
關鍵詞:模型
林姍 陸興利 王茹琳

摘要:基于當前和RCP8.5情景,選用最大熵(MaxEnt)模型對獼猴桃潰瘍病病菌(Pseudomonas syringae pv.actinidiae)在四川省的潛在分布區進行預測,并分析21世紀30年代、50年代、70年代和80年代的適生區變化。結果表明,利用ROC曲線對模型模擬的準確度進行評價,訓練數據和測試數據AUC分別介于0.915~0.970、0.924~0.956之間,預測結果準確。當前氣候條件下,獼猴桃潰瘍病病菌在四川省的高適生區主要位于成都市、德陽市、綿陽市、廣元市、巴中市、達州市和雅安市,中適生區在四川省21地市(州)均有分布。RCP8.5情景下,與當前情景相比,高適生區和低適生區面積均顯著增加,中適生區面積先增加后減少,不同適生區幾何中心位置和遷移規律均有所不同,但總體上均向北移動。
關鍵詞:獼猴桃潰瘍病病菌(Pseudomonas syringae pv.actinidiae);MaxEnt模型;氣候變化;適生分析;地理分析
中圖分類號: S436.634.1+9 ?文獻標志碼: A ?文章編號:1002-1302(2020)03-0124-06
四川省是獼猴桃種植大省,主栽品種以紅陽、東紅、紅什2號、金紅50等紅心獼猴桃為主,栽培面積為46.6萬hm2,占全國紅心獼猴桃栽培面積的90%以上[1]。目前,四川省獼猴桃種植區域包含17個市(州),年產量約39.8萬t,居全國第2位。獼猴桃產業是四川省脫貧攻堅工作的重要手段,因此種植規模仍呈迅速擴大之勢[2]。在發展過程中,由于缺乏合理的種植區劃,全省大面積推廣紅心獼猴桃,出現品種單一化和重大病蟲害突發等問題,特別是獼猴桃潰瘍病的大面積暴發和迅速蔓延,對獼猴桃產業造成嚴重威脅[3]。
獼猴桃潰瘍病危害植株的主干、枝條、花和葉片等部位,病原為丁香假單胞桿菌獼猴桃致病變種(Pseudomonas syringae pv.actinidiae),可通過農事操作、苗木、花粉、風雨和昆蟲等傳播[4-5]。獼猴桃潰瘍病的發生和危害與氣候關系密切,氣候適宜時蔓延擴散極快,且難以根治,處理稍不及時極易造成毀園[6-7]。據統計,2017年獼猴桃潰瘍病在四川省發病面積達1.04萬hm2,約為全省獼猴桃種植面積的26%,對產業發展造成極大威脅。
氣象因素是影響病蟲害發生、消長和蔓延流行的重要環節變量,全球乃至區域性氣候的改變,對病蟲害有重要的影響[8-9]。氣候變化會導致新病菌的出現,可改變病害的危害程度,促使次要病害的危害風險和分布范圍增大,上升為主要病害。氣候變暖通過改變病蟲害的生活習性,如繁殖、生長和越冬等,進而影響其地理分布格局。獼猴桃潰瘍病的發生與氣象關系密切,因此探明氣候變化對四川省獼猴桃潰瘍病病菌分布的影響及對該病害的預測預報和風險區劃具有重要意義[10-11]。氣候變化可改變原有正常氣候條件下生存的寄主植物的生存范圍,進而引起病害微生物和害蟲的生境變化。氣候變化引起的異常天氣,客觀上助長了病蟲害的地理擴張[12-13]。利用已有的氣候數據評價已有病蟲害的適生性,依據可能出現的氣候情景評價分析未來的危害風險是極其必要的。最大熵(MaxEnt)模型是目前應用最多且評價很高的生態位模型軟件,近年來在植保方面的應用包括重大病蟲害氣候適宜性的分析、檢疫性病蟲害入侵可能性的預測和氣候變化對病蟲害分布區影響的預估等[14-16]。MaxEnt模型通過已知的物種地理分布信息,結合環境變量,根據氣候相似原理,計算指定生態位約束下物種分布規律的最理想狀態,即熵最大時目標物種在預測地區的可能分布情況[17]。研究表明,MaxEnt模型較其他模型運行更穩定、操作簡單、運算速度快,且在數據較少時準確性更高,已成為物種地理分布預測的首選模型[18]。
本研究通過實地調查和查閱文獻等方式獲得獼猴桃潰瘍病病菌的分布數據,結合當前和未來的氣候數據,應用MaxEnt模型模擬并預測氣候變化情景下獼猴桃潰瘍病病菌在四川省的潛在分布,分析氣候變化對其分布的影響,旨在為四川省獼猴桃潰瘍病風險分析、預測預報和有效防控等提供理論支撐。
1 材料與方法
1.1 氣候數據的來源與處理
當前和RCP8.5氣候變化情景的數據分別從Worldclim和CCAFS數據庫免費下載,選擇柵格分辨率為2.5 arc-minutes(約4.5 km2)的數據集。選用最常用的19個生物氣候變量作為初始環境變量導入MaxEnt軟件,計算百分比貢獻率。利用Pearson相關系數法消除共線性的影響,依據獼猴桃潰瘍病的生物學特性,最終從19個生物氣候變量中獲得7個變量進行建模[19](表1)。
1.2 分布數據的來源與處理
筆者所在課題組于2015—2018年對四川省獼猴桃主栽區的潰瘍病發生情況進行全面地系統調查,記錄了發病地的經度、緯度。獼猴桃潰瘍病在新西蘭、西班牙、法國、智利、日本、韓國等國家均有發生[20-21],因此采用查詢物種分布數據庫和檢索相關文獻2種方式統計該病菌在全球其他地區的分布信息。查詢的數據庫為歐洲和地中海植物保護組織數據庫(EPPO,https://www.eppo.int/)和全球物種多樣性信息庫(GBIF,http://www.gbif.org/)。上述分布點中有具體經緯度的直接應用,無經緯度的則查詢全球地理信息集成數據庫GeoName獲得位置的經緯度。通過上述方式共獲得286個分布點。參考張東方等的方法[22]對分布數據進行有效篩選以避免空間自相關導致的誤差,使用ArcGIS 10.0軟件的統計分析功能計算并保留1條與網格中心最近的分布記錄,最終獲得148個分布點。分布記錄導入Excel 2010中,格式為物種名+經度+緯度。
1.3 模型的構建和適生等級劃分
將獼猴桃潰瘍病病菌分布點數據導入MaxEnt模型時,設置75%的分布點數據用于建模,25%的分布點數據用于驗證模型;選擇模型自帶的刀切法(Jackknife)測定7個環境變量的重要性;環境變量與獼猴桃潰瘍病病菌存在概率之間的Logistic關系圖由MaxEnt的繪制響應曲線功能完成;其余參數均選擇模型的默認值,重復運行10次進行建模[23-24]。
MaxEnt模型默認適生等級為10級,根據獼猴桃潰瘍病在四川省的實際發生情況并結合相關文獻,利用ArcGIS的Reclassify功能對連續分布概率(P)進行重分類,劃分方法如下:白色代表不適生區,P<0.2;黃色代表低適生區,0.2≤P<0.4;橙色代表中適生區,0.4≤P<0.8;紅色代表高適生區,P≥0.8[25-26]。
1.4 模型模擬結果評價
本研究選用ROC(receiver operating characteristic,ROC)曲線法評價MaxEnt模擬結果的準確性。ROC曲線其曲線下面積(area under curve,AUC)的大小是評估模型預測是否準確的指標[22]。MaxEnt模型固有模塊可繪制ROC曲線,并計算AUC。評估標準定義如下:AUC取值范圍為[0.5,0.6)時,定義模型模擬結果為失敗;AUC取值范圍為[0.6,0.7)時,定義模型模擬結果為較差;AUC取值范圍為[0.7,0.8)時,定義模型模擬結果為一般;AUC取值范圍為[0.8,0.9)時,定義模型模擬結果為好;AUC取值范圍為[0.9,1.0]時,定義模型模擬結果為很好[27-28]。
2 結果與分析
2.1 模擬準確性檢驗
由表2可知,訓練數據的AUC介于0.915~0.970 之間,測試數據的AUC介于0.924~0.956之間,表明構建的5個模型的模擬結果為很好,可用于后續分析。
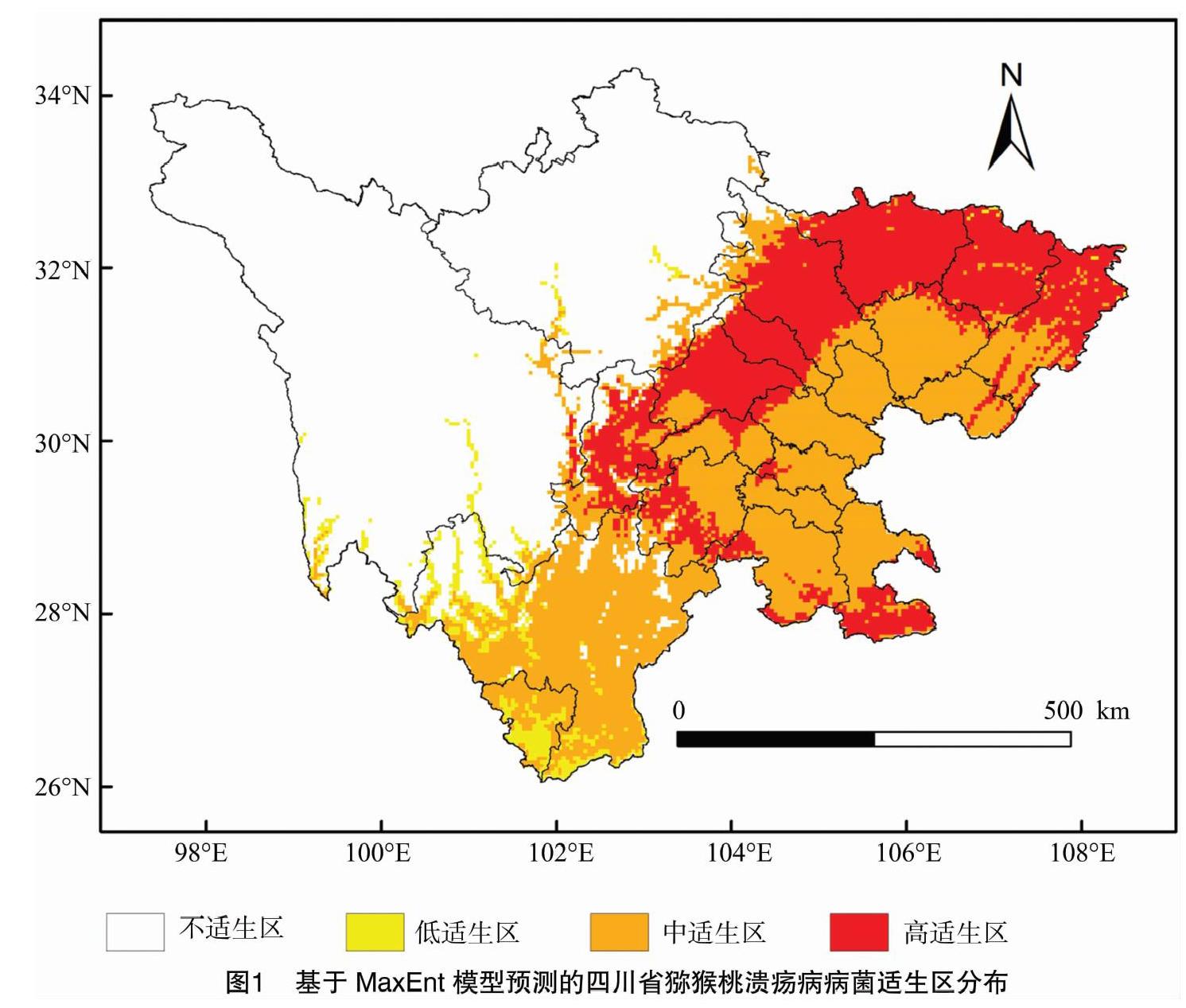
2.2 當前氣候情景下四川省獼猴桃潰瘍病病菌地理分布預測
由圖1可以看出,四川省獼猴桃潰瘍病病菌高適生區位于成都市、德陽市、綿陽市、廣元市、巴中市、達州市、雅安、樂山市、瀘州市、宜賓市、廣安市等,面積達8.41萬km2;中適生區在四川省21個地市(州)均有分布,面積為13.32萬km2;低適生區位于甘孜州、阿壩州、涼山州、攀枝花市、達州市等,面積為1.18萬km2。
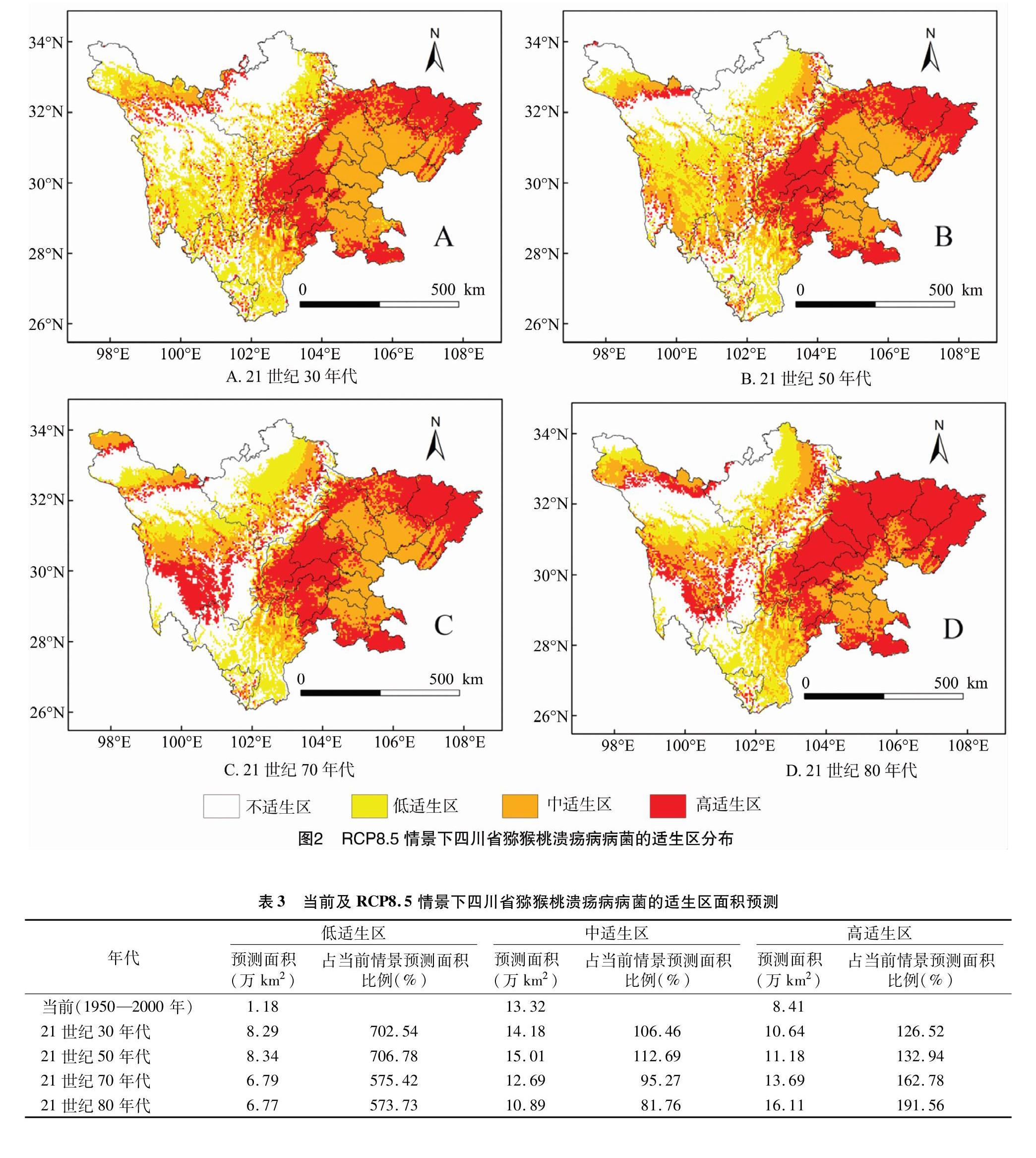
2.3 RCP8.5情景下四川省獼猴桃潰瘍病病菌地理分布預測
RCP8.5氣候變化情景下,21世紀30年代、50年代、70年代和80年代獼猴桃潰瘍病病菌在四川
省的潛在分布如圖2、表3所示。高適生區面積變化趨勢如下:由當前的8.41萬km2分別增加到21世紀30年代、50年代、70年代和80年代的 10.64萬、11.18萬、13.69萬、16.11萬km2。中適生區面積變化趨勢如下:首先由當前的 13.32萬km2 分別增加至21世紀30年代、50年代的14.18萬、15.01萬km2,然后分別減少至70年代和80年代的12.69萬、10.89萬km2。低適生區面積未來增幅最大,由當前的1.18萬km2分別增加至21世紀30年代、50年代、70年代和80年代的 8.29萬、8.34萬、6.79萬、6.77萬km2。
2.4 四川省獼猴桃潰瘍病病菌適生區質心位移軌跡
本研究計算了RCP8.5情景下四川省獼猴桃潰瘍病病菌適生區的質心位移軌跡。由表4可以看出,高適生區質心由當前位置至21世紀30年代、21世紀30年代至50年代、21世紀50年代至70年代、21世紀70年代至80年代依次沿西南57.05 km、東南 8.00 km、西南59.13 km和東北57.11 km移動,至21世紀80年代總體上向西北方向移動 66.59 km。中適生區質心由當前位置至21世紀30年代、21世紀30年代至50年代、21世紀50年代至70年代、21世紀70年代至80年代依次沿西北62.51 km、西北25.32 km、西北17.83 km和東北11.77 km移動,至21世紀80年代總體上向西北方向移動103.13 km。由此可見,未來不同適生區幾何中心位置和遷移規律均有所不同。
3 討論與結論
3.1 獼猴桃潰瘍病病菌在四川省的潛在分布
本研究中在當前氣候情景下,四川省獼猴桃潰瘍病病菌高適生區主要位于成都市、德陽市、綿陽市、廣元市、巴中市、達州市、雅安市等。適生區域(中適生區和高適生區)除在甘孜州和阿壩州外,在其他19個市(州)均有分布。2017年,獼猴桃潰瘍病在四川省14個市(州)發生,其中雅安市、成都市和廣元市發病面積最大,分別占全省發生面積的43%、37%、7%。對比此次預測結果和獼猴桃潰瘍病病菌在四川省的發生現狀可知,除目前已知該病菌廣泛分布在成都、雅安、廣元等地以外,巴中、達州、廣安等地也為該病菌適生等級較高的地區。由此推測,四川省獼猴桃潰瘍病仍存在繼續擴散的可能。因此上述高適生區中,已發現該病菌的地區必須及時采取相應措施進行防治,阻止該病菌向其他地區擴散蔓延;對于具備該病菌適生的寄主植物和氣候條件的潛在適生分布區,應高度重視,加強檢驗檢疫工作,防止該病菌的傳入;獼猴桃潰瘍病自1986年在我國湖南省被發現以來,在長期的擴散傳播過程中產生了較高的種內遺傳多樣性[29-30],且隨著全球氣候的變暖,當前的低適生區或不適生區有可能變為該病原菌的適生區,因此對于非適生區,仍應保持高度警惕。
聯合國政府間氣候變化專門委員會(Intergovernmental Panel on Climate Change,IPCC)第五次評估報告公布了4種氣候變化情景,即RCP2.6情景、RCP4.5情景、RCP6.0情景和RCP8.5情景。RCP8.5情景下,由于缺乏應對氣候變暖的有效措施,導致全球能源需求最高,溫室氣體排放量大,生態環境未得到有效改善,全球平均溫度上升最多[31]。本研究利用MaxEnt模型模擬RCP8.5情景下獼猴桃潰瘍病病菌在四川省適生區的分布變化,與當前情景相對比發現,高適生區和低適生區面積均明顯增加,中、高適生區總體上均有向西北方向移動的趨勢。說明未來氣候變暖將使病菌適生范圍擴大,且向高緯度移動明顯,更利于病菌越冬和生長繁殖。因此未來四川盆地具備獼猴桃潰瘍病大暴發的氣候條件,防控任務依然艱巨。
3.2 模型的選擇
MaxEnt模型基于最大熵理論,利用物種分布數據和環境數據分析熵最大時物種的分布狀態,得到國內外學者的認可。如Elith等對比了多種生態位模型的模擬性能,結果表明MaxEnt模型在16種模型中的預測精度最高[32];Petitpierre等應用生態位模型驗證入侵生物的生態位保守性,表明MaxEnt模型為此項研究的有效工具,適合分析物種地理分布與氣候之間的關系[33];張海濤等應用MaxEnt、GARP、BIOCLIM和DOMAIN等4種模型預測福壽螺在我國的潛在適生區,結果表明,MaxEnt模型的模擬效果顯著優于其他模型[34]。因此本研究選取MaxEnt作為模擬軟件。
3.3 不足和局限性
研究表明,獼猴桃潰瘍病發生流行的非生物因素除氣候外,還與地形特征、土壤類型、土壤理化性狀、獼猴桃栽培密度等息息相關[35-38],本研究僅選擇了氣候因素進行模擬,可能對預測效果有一定影響;其次,獼猴桃潰瘍病病菌的分布數據主要來自實地調查、檢索數據庫和查閱文獻,共獲得148個全球分布點。對四川省全省獼猴桃潰瘍病菌的調查雖較為系統,但也不能確保毫無遺漏。檢索數據庫和查閱文獻獲得的數據中,有的分布點無明確經緯度,通過坐標定位軟件搜索地名確定,不可避免地存在一定的地理誤差。因此下一步工作中,應著重解決上述問題,獲取盡可能全面且準確的分布數據和氣候數據,以為獼猴桃潰瘍病的預測預報和科學防治提供參考。
參考文獻:
[1]四川省“十三五”果樹育種攻關項目組. 產學研協同創新為四川水果產業供給側結構性改革發力[J]. 四川農業科技,2018(4):56-58.
[2]顧 穎,楊錦秀,傅雪梅. 果農選擇經營組織模式的影響因素分析——基于四川省蒼溪縣獼猴桃產業調研[J]. 江蘇農業科學,2017,45(8):342-345.
[3]涂美艷,黃昌學,陳 棟,等. 四川獼猴桃產區潰瘍病綜合防治月歷表[J]. 四川農業科技,2018(1):31-33.
[4]劉 瑤,朱天輝,樊芳冰,等. 四川獼猴桃潰瘍病的發生與病原研究[J]. 湖北農業科學,2013,52(20):4937-4942.
[5]任茂瓊,余 敖,李家慧,等. 四川省北川縣獼猴桃潰瘍病發生特點與綜合防控技術示范[J]. 中國植保導刊,2018,38(11):55-57.
[6]王 麗,周增強,侯 琿,等. 我國獼猴桃細菌性潰瘍病研究分析及防控[J]. 中國南方果樹,2017,46(2):178-182.
[7]潘 慧,胡秋舲,張勝菊,等. 貴州六盤水市獼猴桃病害調查及病原鑒定[J]. 植物保護,2018,44(4):125-131,137.
[8]張 蕾,霍治國,王 麗,等. 氣候變化對中國農作物蟲害發生的影響[J]. 生態學雜志,2012,31(6):1499-1507.
[9]王 麗,霍治國,張 蕾,等. 氣候變化對中國農作物病害發生的影響[J]. 生態學雜志,2012,31(7):1673-1684.
[10]郭建平. 氣候變化對中國農業生產的影響研究進展[J]. 應用氣象學報,2015,26(1):1-11.
[11]周廣勝. 氣候變化對中國農業生產影響研究展望[J]. 氣象與環境科學,2015,38(1):80-94.
[12]霍治國,李茂松,王 麗,等. 氣候變暖對中國農作物病蟲害的影響[J]. 中國農業科學,2012,45(10):1926-1934.
[13]葉彩玲,霍治國,丁勝利,等. 農作物病蟲害氣象環境成因研究進展[J]. 自然災害學報,2005,14(1):90-97.
[14]史 麗,申榮榮,白 娟. 昆蟲適生性分析的研究進展[J]. 內蒙古農業大學學報(自然科學版),2017,38(1):119-126.
[15]趙 力,李慧琪,朱耿平,等. 入侵害蟲椰子木蛾(鱗翅目:木蛾科)在我國的潛在分布[J]. 林業科學,2015,51(10):93-100.
[16]張 超,陳 磊,田呈明,等. 基于GARP和MaxEnt的云杉矮槲寄生分布區的預測[J]. 北京林業大學學報,2016,38(5):23-32.
[17]朱耿平,劉國卿,卜文俊,等. 生態位模型的基本原理及其在生物多樣性保護中的應用[J]. 生物多樣性,2013,21(1):90-98.
[18]許仲林,彭煥華,彭守璋. 物種分布模型的發展及評價方法[J]. 生態學報,2015,35(2):557-567.
[19]張天蛟,劉 剛. 提高生態位模型時間轉移能力的方法研究[J]. 中國農業大學學報,2017,22(2):98-105.
[20]Wilstermann A,Schrader G,Kehlenbeck H,et al. Potential spread of kiwifruit bacterial canker (Pseudomonas syringae pv. actinidiae) in Europe[J]. EPPO Bulletin,2017,47(2):255-262.
[21]Ferrante P,Scortichini M. Redefining the global populations of Pseudomonas syringae pv. actinidiae based on pathogenic,molecular and phenotypic characteristics[J]. Plant Pathology,2015,64(1):51-62.
[22]張東方,張 琴,郭 杰,等. 基于MaxEnt模型的當歸全球生態適宜區和生態特征研究[J]. 生態學報,2017,37(15):5111-5120.
[23]葉永昌,周廣勝,殷曉潔. 1961—2010年內蒙古草原植被分布和生產力變化——基于MaxEnt模型和綜合模型的模擬分析[J]. 生態學報,2016,36(15):4718-4728.
[24]萬基中,王春晶,韓士杰,等. 應用GIS和MaxEnt預測南方紅豆杉潛在地理分布及其保護建議[J]. 江蘇農業科學,2014,42(7):349-352.
[25]趙文娟,陳 林,丁克堅,等. 利用MAXENT預測玉米霜霉病在中國的適生區[J]. 植物保護,2009,35(2):32-38.
[26]孫 穎,秦大河,劉洪濱. IPCC第五次評估報告不確定性處理方法的介紹[J]. 氣候變化研究進展,2012,8(2):150-153.
[27]趙晶晶,高 丹,馮紀年. 基于MaxEnt模型的葡萄根瘤蚜在中國的適生性分析[J]. 西北農林科技大學學報(自然科學版),2015,43(11):99-104,112.
[28]王雷宏,楊俊仙,徐小牛. 基于MaxEnt分析金錢松適生的生物氣候特征[J]. 林業科學,2015,51(1):127-131.
[29]高小寧,鄭 州,趙志博,等. 陜西省獼猴桃細菌性潰瘍病菌(Pseudomonas syringae pv. actinidiae)Rep-PCR的遺傳多樣性分析[J]. 果樹學報,2016,33(3):340-349.
[30]朱海云,李 勃,李 燕,等. 丁香假單胞菌獼猴桃致病變種的遺傳多樣性及進化關系[J]. 微生物學雜志,2013,33(4):66-71.
[31]秦大河,Stocker T.IPCC第五次評估報告第一工作組報告的亮點結論[J]. 氣候變化研究進展,2014,10(1):1-6.
[32]Elith J,Graham C H,Anderson R P,et al. Novel methods improve prediction of species distributions from occurrence data[J]. Ecography,2006,29(2):129-151.
[33]Petitpierre B,Kueffer C,Broennimann O,et al. Climatic niche shifts are rare among terrestrial plant invaders[J]. Science,2012,335(6074):1344-1348.
[34]張海濤,羅 渡,牟希東,等. 應用多個生態位模型預測福壽螺在中國的潛在適生區[J]. 應用生態學報,2016,27(4):1277-1284.
[35]馮 華,李海洲. 周至縣獼猴桃潰瘍病流行規律及其綜合防治措施[J]. 中國植保導刊,2009,29(11):29-30,28.
[36]李有忠,宋曉斌,張學武. 獼猴桃細菌性潰瘍病發生規律研究[J]. 西北林學院學報,2000,15(2):53-56.
[37]李 瑤,承河元,方書苗,等. 獼猴桃細菌性潰瘍病流行預測初探[J]. 應用生態學報,2001,12(3):355-358.
[38]任 平,阮祥穩,趙文娟,等. 獼猴桃潰瘍病P-L菌株誘導植株系統的抗性[J]. 江蘇農業科學,2017,45(21):109-111.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
