魚類水霉病病原菌LY04對環境及營養的適應性
2020-04-10 06:57:04段魏魏申慧魏冬梅
江蘇農業科學 2020年3期
段魏魏 申慧 魏冬梅
摘要:以標準菌株ATCC 26116為對照組,采用單因素試驗方法分別探討菌株LY04對溫度、pH值、鹽度的環境適應性以及對碳源、氮源的營養適應性。結果顯示,菌株LY04與對照株在10~30 ℃的溫度范圍、5~9的pH值范圍內均可生長,但二者在相同環境條件下生長速率差異顯著,LY04較對照株對溫度、pH值的適應性差,鹽度適應性優于對照株;二株菌對營養的要求差異顯著,在相同營養條件下,LY04菌株生長速率顯著快于對照株,試驗株對寡營養適應性優于對照株;NH+4-N在質量濃度0.1%~0.3%范圍內對二株菌生長均有促進作用,NO3-N對菌株LY04的生長有抑制作用,在0.1%質量濃度下可以促進對照株的生長,其余濃度也表現出明顯的抑制作用。綜上,菌株LY04具有很好的環境適應性和營養適應性,這些適應性都與新疆獨特的水域環境相適應。
關鍵詞:水霉;病原菌;環境;營養;適應性
中圖分類號: S941.43+1 ?文獻標志碼: A ?文章編號:1002-1302(2020)03-0190-04
水霉病是各種水產養殖中廣泛存在的一種寄生性疾病,是防治難度最大的魚類病害之一,每年給養殖業帶來巨大的經濟損失[1-2]。由于其特效防治藥物孔雀石綠的禁用,其危害更加普遍和嚴重[3-4]。目前,硫酸銅、臭氧、福爾馬林、重氫硫酸鹽等各種化學試劑及各種中草藥、微生物抗菌劑廣泛用于水霉病防治,但大多只起到預防作用[5-11]。在尋找水霉病防治的有效途徑時,了解其病原是必須的。研究認為,水霉病的病原常見的有水霉、綿霉和腐霉[12-13],具有多樣性,對寄主無嚴格選擇性。
國內外大量學者對水霉病病原菌生理特性的研究表明,其環境適應范圍廣,5~32 ℃溫度范圍[14-16]、4~11的pH值范圍[17-18]、3.5%以內的氯化鈉[14,19-21]含量條件下均可生長。新疆獨特的氣候特點使魚類在歷經嚴冬后的初春易患水霉病,魚類水霉病的病情較我國其他地區更為嚴峻,而目前未見相關研究見諸于眾。本研究對新疆五家渠地區水霉病病原菌的環境適應性進行探討,以期為了解病原進而找到合適的防治水霉病藥物提供數據。
1 材料與方法
1.1 試驗材料
1.1.1 菌種 菌種:LY04,從五家渠患水霉病鰱魚肌肉及體表分離獲得,根據形態學及生理學特征鑒定為水霉目、水霉科、水霉屬[22];標準菌株,多子水霉(saprolegnia fex)ATCC 26116,購自美國典型微生物菌種保藏中心(ATCC)。
1.1.2 培養基 馬鈴薯葡萄糖培養基(pH值 7.0),用于水霉的培養及環境適應性試驗;沙堡氏瓊脂培養基(葡萄糖40 g,蛋白胨10 g,瓊脂15 g,純凈水1 000 mL,pH值5.6±0.2),用于營養適應性試驗。
1.2 試驗方法
1.2.1 環境適應性試驗方法
1.2.1.1 溫度對水霉菌生長的影響 用微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺的水霉菌菌落后重新接種在新的PDA培養基中央。然后分別置于10、15、20、25、30 ℃下恒溫培養72 h,用十字交叉法測量菌落直徑(mm),計算生長率(%)。每組重復3次,用GraphPad Prism 5軟件進行統計分析。
1.2.1.2 pH值對水霉菌生長的影響 配制PDA培養基,常規滅菌后,用3 mol/L HCl和3 mol/L NaOH調節pH值分別為5、6、7、8、9。采用微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺的水霉菌菌落后重新接種在滅過菌的不同pH值的PDA培養基中央。然后置于25 ℃下恒溫培養72 h。用十字交叉法測量菌落直徑(mm),計算生長率(%)。每組重復3次。用GraphPad Prism軟件進行統計分析。
1.2.1.3 鹽度對水霉菌生長的影響 用微生物平板打孔器在培養有水霉菌PDA培養基上,輕輕打取長勢較旺的水霉菌菌落后,分別接種于含NaCl質量分數為0.0%、0.5%、1.0%、1.5%、2.0%、2.5%的PDA平板中央,然后置于25 ℃恒溫培養48 h后,用十字交叉法測量菌落直徑(mm),計算生長率(%)。每組重復三次,用GraphPad Prism軟件進行統計分析。
1.2.2 營養條件適應性試驗方法
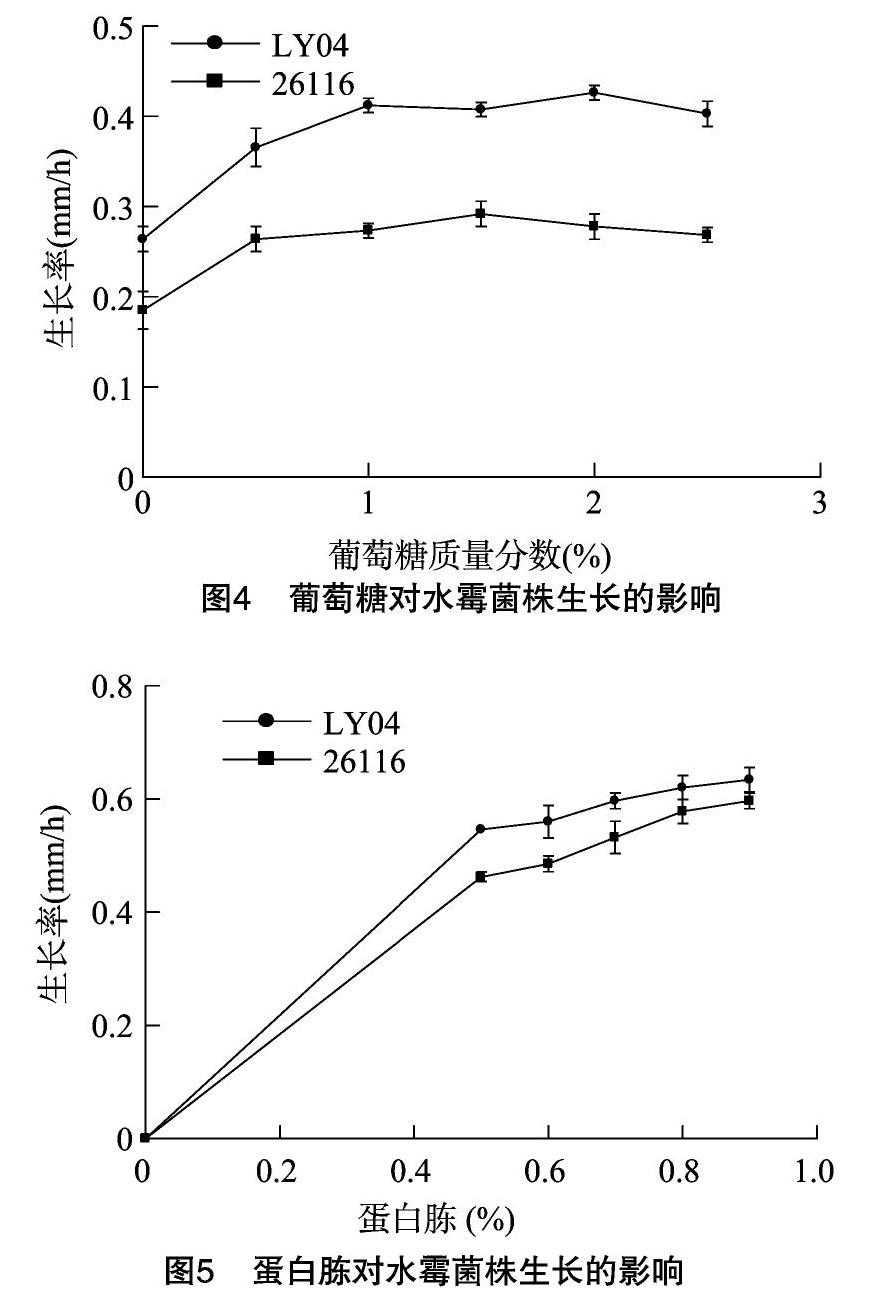
1.2.2.1 葡萄糖對水霉菌株生長的影響 采用滅過菌的微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺較一致的水霉菌菌落后,接種在葡萄糖質量濃度為0.0%、0.5%、1.0%、1.5%、2.0%、2.5%的沙堡氏瓊脂培養基(去掉了葡萄糖組分)中央。然后置于25 ℃溫度條件下恒溫培養,分別觀察24、48、72 h的生長情況并用十字交叉法測量菌落直徑,每組3個平行。
1.2.2.2 蛋白胨對水霉菌株生長的影響 采用滅過菌的微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺較一致的水霉菌菌落后,接種在蛋白胨質量濃度為0.0%、0.5%、0.6%、0.7%、0.8%、0.9%的沙堡氏瓊脂培養基(去掉了蛋白胨組分)中央。然后置于25 ℃下恒溫培養,分別觀察24、48、72 h的生長情況并用十字交叉法測量菌落直徑,每組3個平行。
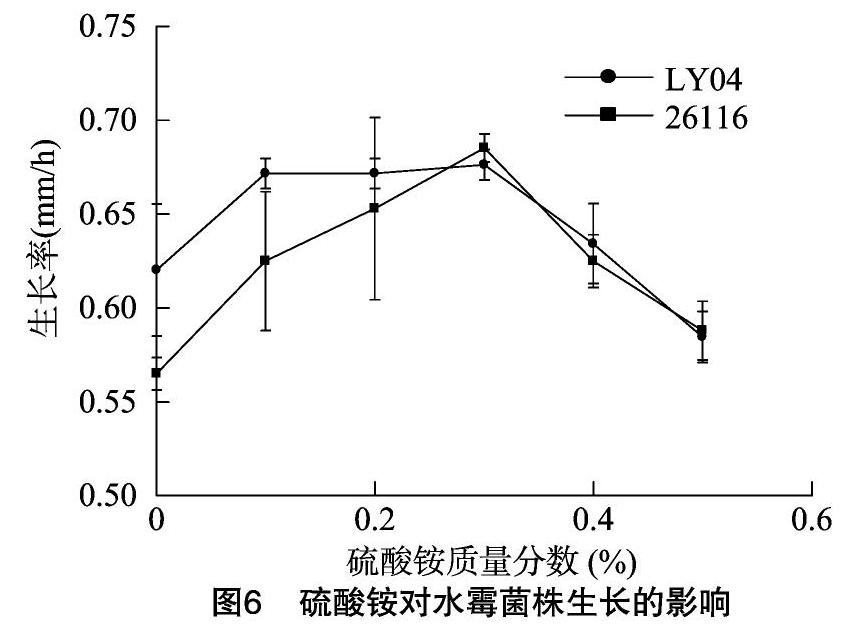
1.2.2.3 硫酸銨對水霉菌株生長的影響 采用滅過菌的微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺較一致的水霉菌菌落后,接種在含硫酸銨質量濃度為0.0%、0.1%、0.2%、0.3%、0.4%、0.5%的沙堡氏瓊脂培養基中央,后置于25 ℃下恒溫培養,分別觀察24、48、72 h的生長情況并用十字交叉法測量菌落直徑,每組3個平行。
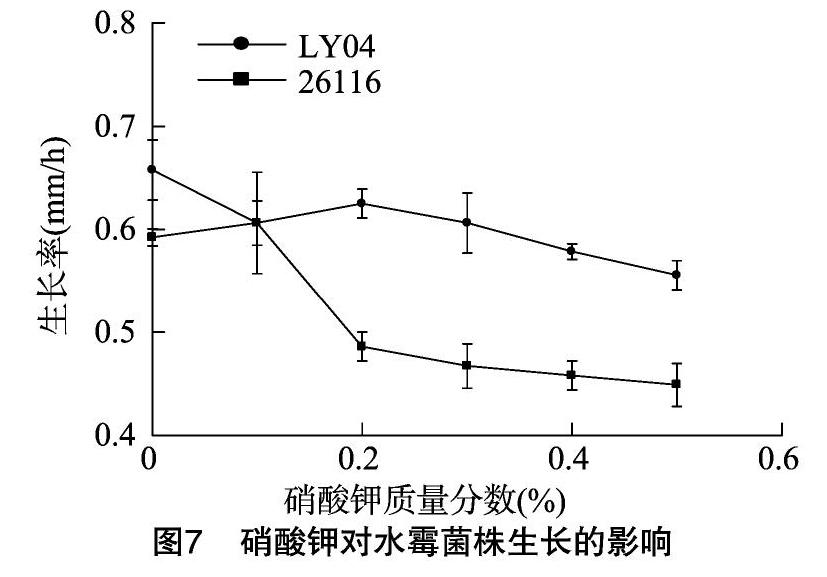
1.2.2.4 硝酸鉀對水霉菌株生長的影響 采用滅過菌的微生物平板打孔器在培養有水霉菌的PDA培養基上,輕輕打取長勢較旺較一致的水霉菌菌落后,接種在含硝酸鉀質量濃度為0.0%、0.1%、0.2%、0.3%、0.4%、0.5%的沙堡氏瓊脂培養基中央。然后置于25 ℃下恒溫培養,分別觀察24、48、72 h的生長情況并用十字交叉法測量菌落直徑,每組3個平行。
2 結果與分析
2.1 環境適應性結果
2.1.1 溫度對水霉菌生長的影響 微生物的生長繁殖有其適宜的溫度范圍,培養溫度過高或過低,都會造成微生物生長緩慢,甚至導致微生物的死亡。溫度對2株水霉菌株生長的影響見圖1。由圖1可知,菌株LY04及標準菌在10~30 ℃范圍內均可生長。菌株LY04在為25 ℃達到最大生長速率,標準菌株在20 ℃和25 ℃時生長最好且其生長速率在同溫度下高于菌株LY04。
2.1.2 pH值對水霉菌生長的影響 pH值對微生物的影響是多方面的,它不僅可以影響微生物對物質的吸收能力,還可以影響酶的活性。過酸或過堿的環境會導致微生物生長緩慢甚至導致微生物死亡。pH值對2株水霉菌株生長的影響見圖2。由圖2可知,菌株LY04和標準菌在pH值5~9范圍內均可生長,對于酸堿環境的適應性較強,菌株LY04在pH值=7時生長最好,標準菌在pH值=6時生長最好。在相同pH值條件下,標準菌株的生長速率高于菌株LY04;但試驗菌株對酸堿變化更敏感。
2.1.3 鹽度對水霉菌生長的影響 海水中魚類水霉病的發生率很低,如受水霉菌感染的鱒魚,當回到海水時,水霉病癥狀逐漸消失,說明水霉病原菌對鹽度敏感。鹽濃度對2株水霉菌株生長的影響,由圖3可知,菌株LY04在NaCl質量分數為0.5%、1.0%、1.5%、2.0%時生長被促進,而在NaCl質量分數為2.5%時生長被抑制,最適NaCl質量分數為1.5%; 標準菌株在NaCl質量分數為0.5%~2.0%范圍內生長幾乎不受影響,當鹽度達2.5%時生長顯著被抑制。在相同鹽度條件下,試驗株的生長速率顯著高于對照株。
2.2 營養條件適應性試驗結果
2.2.1 葡萄糖對水霉菌生長的影響 葡萄糖不僅為生物的生長提供細胞的碳架,還提供細胞生命活動所需要的能量。在微生物的培養基中一般都過量添加葡萄糖,但過量的葡萄糖對微生物的生長是有影響。葡萄糖對2株水霉菌株生長的影響見圖4。由圖4可知,在不添加葡萄糖時蛋白胨中的碳源就可使2個株菌生長,在有蛋白胨的培養基中只需添加少量葡萄糖即可使水霉菌生長旺盛(0.5%與對照相比 P<0.05)。在相同條件的同一葡萄糖濃度下,菌株LY04的生長較標準菌株旺盛。
2.2.2 蛋白胨對水霉菌生長的影響 氮是微生物必需的營養物質之一,蛋白胨主要作為氮源用于微生物的培養。蛋白胨對2株水霉菌生長的影響見圖5。由圖5可知,2株菌在不添加蛋白胨時均不生長;菌株LY04在0.5%的蛋白胨下,生長速率顯著提高,0.8% 時達到飽和;標準菌在0.7%的蛋白胨下,生長速率顯著提高,0.8%時達到飽和。在相同條件的同一低濃度蛋白胨下,菌株LY04生長較標準菌旺盛。
2.2.3 硫酸銨對水霉菌生長的影響 眾多研究表明,真菌生長時對不同形態的氮鹽利用效率不同。NH+4-N以還原態形式存在,添加在培養基中主要增加還原態的氮。硫酸銨對2株水霉菌生長的影響見圖6。由圖6可知,0.1%~0.3%濃度范圍的硫酸銨對2株菌的生長起促進作用,硫酸銨濃度高于 0.5% 時開始抑制菌株生長,即2株菌在一定濃度范圍內對銨態氮有偏好。相同條件同一濃度硫酸銨條件下,2株菌生長差異不明顯。
2.2.4 硝酸鉀對水霉菌生長的影響 硝態氮是氮元素的另外一種存在形式,為氧化態氮。在培養基中添加硝酸鉀,主要增加氧化態的氮。硝酸鉀對2株水霉菌生長的影響見圖7。由圖7可知,添加硝酸鉀菌株LY04的生長受到抑制;標準菌株僅在0.1%的硝酸鉀條件下生長稍有促進,增加濃度其生長被抑制菌。
3 討論
3.1 水霉病原菌的環境適應性
試驗菌株LY04在10~30 ℃溫度范圍、5~9的pH值范圍內均可生長,對溫度和pH值的適應性較廣。與標準菌株相比,溫度、pH值對試驗菌株LY04影響更大,在相同的溫度、pH值條件下,試驗菌株的生長速率要慢于標準株,當溫度達到適溫時,試驗菌株生長迅速增快,這與魚類水霉病的發生有對應關系,說明試驗菌株對新疆獨特的水域環境有高度的適應性。在初春時節,魚類由于越冬時的凍傷易感水霉病,但病情發展緩慢,這與此時溫度較低,水霉菌生長緩慢相關;初夏時節,在魚類繁殖期,水霉菌大量侵襲魚卵,造成水霉病暴發,此時池塘水溫恰與水霉菌最適生長溫度一致。在相同的外界條件下,菌株LY04對鹽度較標準菌有更好的忍耐性,生長速率均好于標準株,說明試驗菌株對鹽度也有更好的適應性。新疆的氣候條件導致新疆水域環境較高的鹽堿度,從分離獲得的菌株來看,新疆水霉病原菌都具有良好的鹽堿抵抗力,因而在新疆魚類水霉病的防治難度更大。
3.2 水霉病原菌的營養適應性
水域環境條件對于微生物生長來講應該屬于貧營養,通過營養條件適應性試驗發現,水霉病原菌對營養條件的要求確實較低,2株菌在不存在任何形式的糖時仍可生長;對氮源的要求也很低,低濃度的蛋白胨就可以使2株菌迅速生長,說明2株菌對貧營養的適應性都較好,相對而言,試驗菌株LY04在相同營養條件下生長速率明顯高于標準菌株,說明試驗菌株在貧營養環境中更易生長。新疆水域由于鹽堿度較大,生物豐富度相對小,因此水域的營養條件相對貧瘠,試驗菌株對貧營養的耐受恰好與此吻合。
3.3 不同形式氮源的存在對水霉病原菌的影響
氮作為地球上存在最多的元素,可以氧化態、還原態、氧化-還原態多種形式存在。許多研究都表明,生物可以利用不同形式的氮源。對于水霉病病原菌而言,尚未見此方面的研究報道。本研究在添加了蛋白胨的培養基中分別添加硫酸銨、硝酸鉀,以便探討試驗菌株對不同形式的氮的利用情況。由結果可知,2株菌都表現出了對銨態氮的更好利用,硝態氮則表現出了抑制作用。水域環境由于溶解氧的限制,一般處于還原狀態,2株菌都能更好地利用還原態氮——銨態氮,也說明水霉病原菌對水環境的良好適應性。
綜上所述,發現試驗菌株具有很好的環境適應性和營養適應性,這些適應性都與新疆獨特的水域環境相適應;對照菌株也表現出了較強的適應性,這也許是魚類水霉病廣泛發生的主要原因。
參考文獻:
[1]Tampieri M P,Galuppi R,Carelle M S,et al. Effect of selected essential oils and pure compounds on Saprolegnia parasitica[J]. Pharmaceutical Biology,2003,41(8):584-591.
[2]孟思妤,孟長明,陳昌福. 魚類水霉病的發生原因與預防措施[J]. 漁業致富指南,2016(15):60-61.
[3]王發榮. 水庫魚種水霉病綜合防治[J]. 農業與技術,2017,37(14):112.
[4]黎姍梅. 水產養殖中水霉病的防治[J]. 科學養魚,2017(2):91.
[5]Forneris G,Bellardi S,Palmegiano G B,et al. The use of ozone in trout hatchery to reduce saprolegniasis incidence[J]. Aquaculture,2003,221(1/2/3/4):157-166.
[6]Straus D L,Farmer B D,Ledbetter C K,et al. Use of copper sulfate to control egg saprolegniasis at a commercial sunshine bass hatchery[J]. North American Journal of Aquaculture,2016,78(3):243-250.
[7]Gieseker C M,Serfling S G,Reimschuessel R. Formalin treatment to reduce mortality associated with Saprolegnia parasitica in rainbow trout,Oncorhynchus mykiss[J]. Aquaculture,2006,253(1/2/3/4):120-129.
[8]Ali E H. Morphological and biochemical alterations of oomycete fish pathogen Saprolegnia parasitica as affected by salinity,ascorbic acid and their synergistic action[J]. Mycopathologia,2005,159(2):231-243.
[9]宋學宏,陳 葵,王永玲,等. EM及重氫硫酸鹽對黃顙魚卵孵化中水霉的抑制作用[J]. 淡水漁業,2007,37(1):9-12,18.
[10]劉永濤,艾曉輝,楊秋紅,等. 15種中草藥超臨界CO2流體萃取物對2種水霉菌的抑制作用研究[J]. 淡水漁業,2015,45(3):40-45.
[11]宋增福,范 斌,佘林榮,等. 一株具水霉抑制特性的芽孢桿菌篩選及其分子生物學鑒定[J]. 淡水漁業,2012(3):28-31.
[12]Hсаева Н М,李永芳.魚類的霉菌病[J]. 國外水產,1993(3):13-16.
[13]王立寶,劉文霞,左萬星,等. 松蘿酸對3種水霉病致病菌的抑菌作用[J]. 河北師范大學學報(自然科學版),2017,41(4):354-357.
[14]Hatai K,Willoughby L G,Beakes G W. Some characteristics of saprolegnia obtained from fish hatcheries in Japan[J]. Mycological Research,1990,94(2):182-190.
[15]Koeypudsa W,Phadee P,Tangtrongpiros J,et al. Influence of pH,temperature and sodium chloride concentration on growth rate of Saprolegnia sp.[J]. Journal of Scientific Research at Chulalongkorn University,2005,30(2):123-130.
[16]甄 珍,王 荻,劉紅柏,等. 山女鱒水霉病病原的分離鑒定及其生物學特性[J]. 江西農業大學學報,2015,37(2):333-338.
[17]熊 剛,尹思璐,梁永增,等. 河鱸水霉病病原菌的研究[J]. 安徽農業科學,2013,41(8):3411-3413,3439.
[18]夏文偉,曹海鵬,王 浩,等. 彭澤鯽卵源致病性水霉的鑒定及其生物學特性[J]. 微生物學通報,2011,38(1):57-62.
[19]楊偉杰,薛鵬偉,邵 偉,等. 大西洋鮭水霉病鹽浴治療初步研究,河北漁業2017(2),37-39
[20]Hussein M M A,Hatai K.Pathogenicity of Saprolegnia species associated with outbreak of salmonid saprolegniosis in Japan[J]. Fisheries Science,2002,68:1067-1072.
[21]張書俊,楊先樂,李 聃,等. 施氏鱘水霉病病原的初步研究[J]. 中國水產科學,2009,16(1):89-96.
[22]熊 剛,黃奕童,王詠星. 新疆五家渠水霉病原菌多樣性的初步研究[C]//第九屆全國微生物學青年學者學術研討會論文集. 武漢,2012.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
今日農業(2021年11期)2021-08-13 08:53:34
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
中國生殖健康(2018年6期)2018-11-06 07:09:28
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
