基于轉錄組學的釀酒酵母耐酸機制解析
2020-04-13 13:33:28田甜甜孫軍勇蔡國林楊華吳殿輝陸健
食品與發酵工業 2020年6期
關鍵詞:差異
田甜甜,孫軍勇,蔡國林,楊華,吳殿輝*,陸健*
1(工業生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)2(糧食發酵工藝與技術國家工程實驗室(江南大學),江蘇 無錫,214122)3(江南大學 生物工程學院,江蘇 無錫,214122)
釀酒酵母(Saccharomycescerevisiae)作為傳統生產果酒的主要菌種,在群體基因組水平經歷了大量的系統進化、種群分化和適應性馴化等過程,具有清晰的遺傳背景[1]。在葡萄酒工業生產中,S.cerevisiae最適pH值通常在3.0~3.6[2],與葡萄相比,青梅和山楂等水果的有機酸含量偏高(15~20 g/L-1),導致發酵pH偏低(2.7~2.9),而目前生產高酸度果酒(青梅酒、山楂酒等)使用的酵母仍是葡萄酒釀酒酵母,最終影響了此類果酒的發酵周期和酒體風味。因此,選育耐酸性能良好的S.cerevisiae成為釀造高酸度果酒育種的新方向。
隨著釀酒工業需求的增加和菌種選育技術的提高,S.cerevisiae的耐酸機制引起了人們的廣泛關注[3]。目前,關于S.cerevisiae耐酸機制的研究主要集中在燃料乙醇和/或有機酸的工業生產上[4],而關于果酒釀造中與耐酸性相關的研究仍然較少。于玲對篩選出耐酸石榴酒酵母的發酵性能進行初探,但未對耐酸性能進行深入研究[5]。本實驗室前期從青梅酒發酵液中分離出1株S.cerevisiaeET008,在此基礎上通過等離子誘變技術(atmospheric and room temperature plasma,ARTP)、高通量篩選(high-throughput screening,HTS)和適應性實驗室進化(adaptive laboratory evolution,ALE)獲得1株耐酸性較好的S.cerevisiaeET008-c54,該菌種發酵生產的青梅酒具有典型性風味。因此,闡明ET008-c54在酸性環境中的生物學機制為提高高酸度果酒品質奠定基礎。隨著高通量測序技術的發展,基因組學、轉錄組學等技術為研究S.cerevisiae細胞生理代謝提供了重要手段,使人們對S.cerevisiae的基因背景有了更深入的了解,其中轉錄組對于解釋基因組的功能元件、揭示細胞和組織的分子成分以及對發育和進化的理解具有重要作用[6]。
本研究基于轉錄組分析S.cerevisiaeET008-c54和ET008的差異表達基因,同時結合生物信息學手段對差異基因進行注釋,最終篩選出與耐酸相關的關鍵基因并進行驗證。在此基礎上對ET008-c54的發酵特性進行進一步分析,從而為S.cerevisiae的耐酸調控機制的研究提供理論基礎,為后續高酸度果酒的生產提供科學依據。
1 材料與方法
1.1 菌株和培養基
S.cerevisiaeET008,從青梅酒發酵過程中分離并保存于本實驗室;ET008-c54,是ET008經ARTP、HTS和ALE獲得的耐酸菌株。
發酵培養基(g/L):葡萄糖200.0、酵母提取物12.0、(NH4)2SO43.0、KH2PO42.0、MgSO4·7H2O 1.0、CaCl2·2H2O 0.1、NaCl 0.1,用HCl溶液將pH調節至2.5[7]。
1.2 試劑和儀器
3,5-二硝基水楊酸、Tris-HCl、EDTA、二硫蘇糖醇等,國藥集團化學試劑有限公司;標準品乙酸、乙醇、甘油和麥角甾醇,Sigma公司。
RNA提取試劑盒(FastPure Cell/Tissue Total RNA Isolation Kit RC101)、反轉錄試劑盒(Hiscript Ⅱ Q RT SuperMix for qpcr)和AceQ RT-PCR試劑盒(Universial SYBR qPCR Master Mix),南京諾唯贊生物科技有限公司;H+-ATPase活性測定試劑盒,Sigma公司。
Biostat Q發酵罐,德國Sartorius公司;實時熒光定量PCR儀,德國 Eppendorf 公司;Shimadzu UV-1800分光光度計,德國Shimadzu Europe GmbH公司;6890 N氣相色譜,美國Agilent公司。
1.3 總RNA提取及測序
在pH 2.5的條件下,培養S.cerevisiaeET008-c54和ET008生長至對數中期,分別取10 mL菌液于5 000×g離心5 min收集菌體,用無菌水洗滌3次,在液氮中速凍10 min,于-80 ℃保存備用。總RNA提取方法,按照試劑盒說明書進行。利用Nanodrop檢測RNA濃度及瓊脂糖凝膠檢測RNA完整性以及基因組污染情況。檢測合格的RNA用于后續Illumina測序儀測序。轉錄組測序分析,由上海歐易生物醫學科技有限公司完成。
將高通量測序獲得的每個樣本的轉錄組文庫,經過CASAVA堿基識別分析轉化為原始測序序列(raw data),再經過去除接頭和過濾低質量數據處理,所獲得的RNA序列(clean data)作為本研究的基本數據。利用hisat2將過濾后的RNA序列與S.cerevisiae模式菌株S288的參考基因組進行序列比對,獲取在參考基因組或基因上的位置信息,以及ET008-c54和ET008特有的序列特征信息。已知的參考基因序列以及注釋文件做為數據庫,采取序列相似性比對的方法鑒定出各蛋白編碼基因在各樣本中的表達豐度。使用htseq-count軟件獲取每個樣本中比對到蛋白編碼基因上的reads數,cufflinks軟件來計算蛋白編碼基因的表達量FPKM值。差異表達基因(differentially expressed genes,DEGs)使用DESeq R包功能的EstimateSizeFactors和nbinomTest進行鑒定。將差異表達倍數(fold change,FC)≥2或≤0.5和P<0.05設置為顯著差異表達的閾值。
1.4 基因功能注釋分析
利用Blast2go在線數據庫對ET008-c54和ET008的差異基因分別進行GO(Gene Ontology)數據庫(http://www.gEneontology.org/)和KEGG(Kyoto Encyclopedia of Genes and Genomes)數據庫(http://www.genome.jp/kegg/)注釋分析。利用DESeq2方法對定量結果進行差異表達分析并獲得差異表達基因列表,再通過GOSeq方法采取超幾何分布樣本抽取方法進行差異表達的基因功能的GO富集和KEGG代謝通路富集。
針對現有技術的不足,提供了一種基于企業培訓體系下的專業人才培養的質量分析板,具備更加方便清理的優點,解決了現有專業人才培養用質量分析板大多采用白板和白板筆,當使用一定時間后,由于白板上的字痕未擦干凈而留下的污漬,難以清洗,影響美觀和使用的問題。
1.5 RT-qPCR驗證
利用上述1.3獲得的RNA樣品及PrimeScript RT試劑和SYBR Premix Ex Taq試劑盒在定量實時PCR系統上進行PCR反應,以測定8個靶基因的mRNA水平。以ACT1基因作為內標,使用2-ΔΔCt算法計算相對基因表達[8]。表1列出了所用的引物。
1.6 不同pH條件下2菌株的生長情況
在pH為3.0和2.5基礎培養基條件下,分別對ET008-c54和ET008進行三角瓶培養試驗,每間隔10 h 取樣測定其OD600并利用美蘭染色活細胞計數法計數[9],直至培養結束。
1.7 發酵性能驗證及分析方法
分別將ET008-c54和ET008利用配有連續數據采集功能的5 L Biostat Q發酵罐進行發酵試驗。初始OD600為0.2,發酵溫度25 ℃,攪拌速度為120 r/min,并通過自動添加2 mol/L HCl溶液調節pH 2.5。每間隔24 h取樣,直至發酵結束。
菌體濃度的測定:使用分光光度計測定菌液的OD600。

表1 本研究中用到的引物
葡萄糖含量的測定[10]:參照3,5-二硝基水楊酸法(3,5-dinitrosalicylic acid,DNS)進行。
生物量的測定:將5 mL細胞懸浮液轉移至預先稱重的10 mL離心管中,以5 000×g離心10 min,然后用去離子水洗滌2次并離心,于105 ℃下干燥至恒重。
乙酸、乙醇、甘油含量的檢測:樣品離心后經0.45 μm PTFE過濾器過濾,并保存于-20 ℃直至分析。使用氣相色譜儀通過外標法進行測定。升溫程序為進樣器和檢測器的溫度均為260 ℃;柱溫在40 ℃保持8 min,然后以10 ℃/min升至260℃并保持5 min;載體為He,流速1.2 mL/min。與標準品的保留時間進行比較以確定該物質,并通過標準曲線進行定量。
麥角甾醇含量的測定:菌體經無菌水洗滌后,在液氮中冷凍并保存在-80 ℃直至分析。將1 g干細胞重懸于100 mL 25%KOH中,加入5 mL甲醇,然后在95 ℃回流2.5 h進行皂化反應。冷卻至室溫,用70 mL乙醚萃取,并振搖15 min。將混合物轉移至分配器裝置靜置2 h后分離,280 nm處測定上清液在的吸光度,并將其與麥角固醇的標準曲線進行比較。
H+-ATPase活性的測定:通過超聲法裂解處于對數期的細胞用5 mL溶液(0.1 mol/L Tris-HCl、0.33 mol/L蔗糖、5 mmol/L EDTA和2 mmol/L二硫蘇糖醇)稀釋并離心。將沉淀重懸于100 μL溶液(10 mmol/L Tris-HCl、20%甘油、0.1 mmol/L EDTA和0.1 mmol/L二硫蘇糖醇)中,并于-80 ℃保存。根據H+-ATPase活性測定試劑盒測定H+-ATPase的活性。
1.8 數據分析
采用OriginPro 7.5進行單因素方差分析(one-way analysis of variance,ANOVA)及Tukey檢驗來確定數據的顯著差異(P<0.05),并利用GraphPad Prism 7.04(GraphPad Software Inc,San Diego,CA)進行繪圖。所有實驗均重復3次,結果均以x±SD表示。
2 結果與分析
2.1 測序數據的統計與質控報告
測序數據質量的水平直接決定后續測序結果的準確性。采用Illumina HiSeqTM2500高通量測序技術對ET008-c54和ET008進行轉錄組測序,樣品各項檢測參數均達到質量要求,結果如表2所示。經質控過濾后,ET008-c54和ET008后分別得到49.48 Mb和 49.27 Mb原始數據,其中過濾后的RNA序列分別占97.51%和96.28%。ET008獲得7.40 Gb個堿基,有效數據比例為94.50%;而ET008-c54獲得7.41 Gb個堿基,有效數據比例為94.37%。統計顯示,低質量(Q<30)堿基比例極少,表明測序效果較好[11]。ET008測序堿基質量值Q30超過94.56%,平均GC含量為42.15%,而ET008測序堿基質量值Q30超過93.89%,平均GC含量為42.24%。ET008-c54和ET008重測序結果Q30都在90%以上,說明基因組重測序原始數據結果可靠,可進行后續信息分析。

表2 測序數據統計
注:Q30表示原始數據中Qphred>30的堿基占總體堿基的百分比
2.2 差異表達基因(DEGs)篩選
經過數據質控與參考基因組比對,差異表達基因的篩選條件為FC≥2或≤0.5以及P<0.05。差異分析結果如圖1-a所示,共鑒定出688個差異表達基因,其中有364個基因顯著上調,324個基因顯著下調。差異表達上調基因中(圖1-b),FC>10倍的基因占上調差異表達基因總數的4.12%,FC 5~10倍的基因占上調差異表達基因總數的6.33%,而FC 2~5 倍的基因占上調差異表達基因總數的89.08%;差異表達下調基因中,FC 0.25~0.5倍的基因占下調差異表達基因總數的88.79%,而FC 0~0.25倍的基因占上調差異表達基因總數的11.21%。相比較而言,菌株ET008-c54的FC在2倍以內和0.25~0.5倍的上調基因和下調基因的數量相當,而FC > 2和0.25~0.5的基因中上調基因數量明顯高于下調基因數量,說明這些明顯上調的基因可能是潛在的導致ET008-c54具有耐酸性能的關鍵基因。
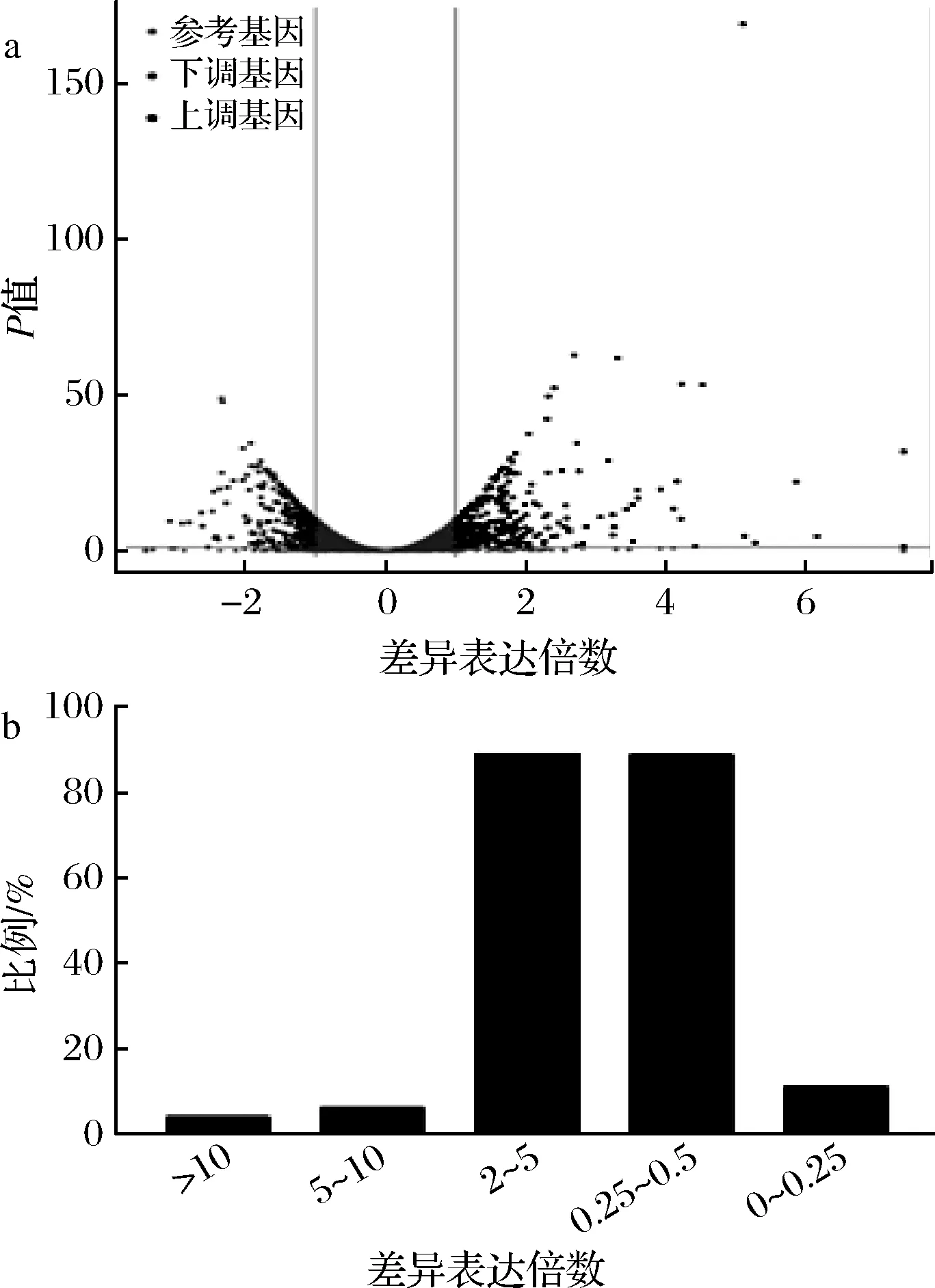
a-上調和下調基因的分布;b-不同差異表達倍數的基因所占的比例
通過代謝通路分析以及相關文獻相結合[12-13],進一步確定了與耐酸性的相關的關鍵差異表達基因,其注釋情況如表3所示。鐵穩態是由復雜的代謝途徑的協調運輸來調節的,它直接參與關鍵的生理過程,例如,細胞存活、耐酸和抗氧化應激[14]。在本研究中,與鐵吸收有關的基因顯著上調,尤其是FET3(167.87)和IZH4(18.89)。INO1(23.02)的顯著上調加速了細胞膜的重構,以減少氧化損傷,表明此基因的表達與細胞內的氧化應激密切相關[15]。另外,與麥角甾醇合成有關的基因ERG5(12.21)也顯著性上調。總之,確定了8種與耐酸性相關的關鍵基因。
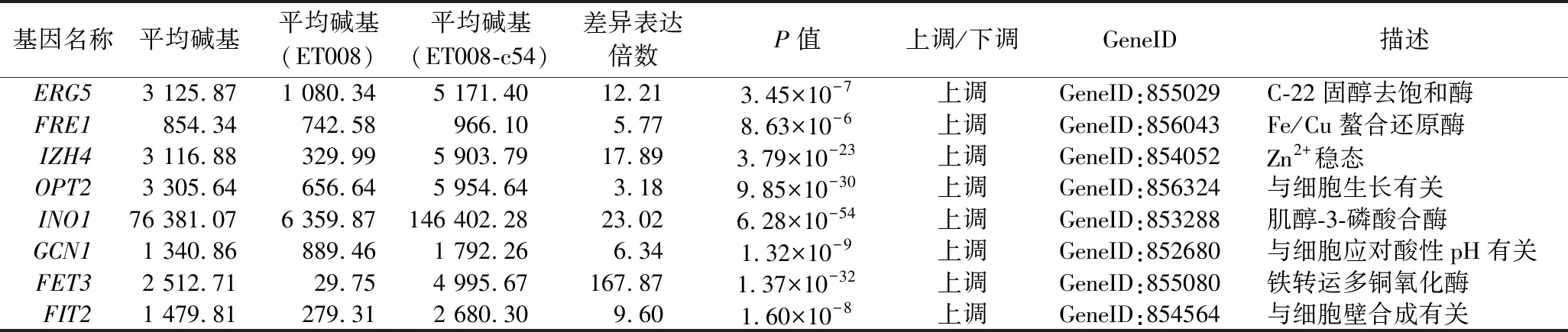
表3 關鍵耐酸差異表達基因的注釋
2.3 差異表達基因功能的GO富集分析
獲得差異表達基因之后,與GO數據庫進行比對并對差異基因進行GO富集分析,如圖2所示。GO數據庫把基因的本體分為三類:生物過程(biological process, BP)、細胞組成(cellular component, CC)和分子功能(molecular function, MF)。由圖2可知,ET008-c54和ET008的差異基因主要富集在BP過程的細胞質翻譯(GO∶0002181)、鐵離子穩態(GO∶0055072)和麥角甾醇的生物合成過程(GO∶0006696);差異基因主要集中CC過程的膜的組成部分(GO∶0016021)、質膜的組成部分(GO∶0005887)和真菌型細胞壁(GO∶0009277);差異基因主要富集在MF上的主要是亞鐵跨膜轉運蛋白活性(GO∶0015093)、鐵離子跨膜轉運蛋白活性(GO∶0005381)和錳離子跨膜轉運蛋白活性(GO∶0005384)。上述結果與關鍵耐酸基因的結果是一致的,進而說明酸含量過高會影響細胞膜的生理功能進而影響微生物的生長及代謝[16]。
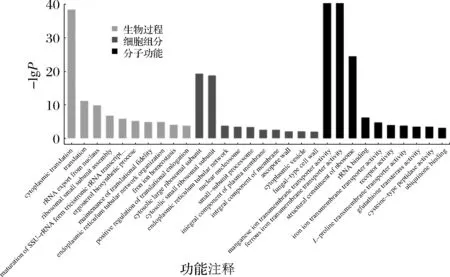
圖2 差異基因的GO功能注釋
2.4 KEGG富集的差異表達基因的功能
對生物體內差異表達基因的進一步通過KEGG數據庫進行功能分析,有助于了解基因的生物學功能[17]。如圖3所示,圖中橫坐標為注釋到KEGG通路上的差異基因數與差異基因總數的比值,縱坐標為KEGG通路,點的大小代表注釋到KEGG通路上的基因數,顏色從紅到紫代表富集的顯著性大小。圖3富集分析表明,DEGs共涉及30條通路,數量較多的幾個被注釋的信號通路為核糖體(ko03010)、類固醇生物合成(ko00100)和氨基苯甲酸酯降解(ko00627)等。綜上可知,此結果與前期結果是一致的。
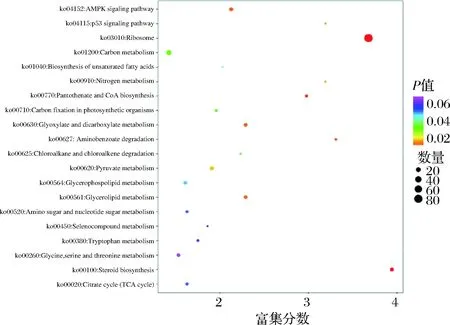
圖3 差異基因的KEGG通路分析
2.5 差異表達基因的RT-qPCR驗證
基于轉錄組數據,確定ET008-c54中與耐酸性相關基因的表達水平發生了顯著性變化。為進一步驗證ET008-c54中的基因差異表達情況,選取8個與耐酸性相關的關鍵基因(ERG5、FRE1、IZH4、OPT2、INO1、GCN1、FET3和FIT2),以驗證轉錄組測序結果的可靠性。如圖4所示,與ET008相比,ET008-c54中與耐酸性有關的基因的表達水平增強,其中ERG5、FRE1、IZH4,OPT2、INO1、GCN1、FET3和FIT2基因的表達水平是3.20、6.27、5.24、3.79、5.83、7.84、9.55和4.07倍。RT-qPCR驗證結果與轉錄組測序結果中差異表達基因的變化趨勢一致,表明本研究結果的有效性和可靠性。

圖4 差異表達基因的RT-qPCR驗證
2.6 不同pH條件下2菌株的生長情況
ET008-c54和ET008在pH 3.0和2.5條件下的細胞生長情況如圖5所示。由圖5可知,在pH值為3.0時,整個培養階段ET008-c54和ET008的OD600差異都不明顯;而pH值為2.5時,整個過程ET008-c54和ET008的OD600差異較大,且在穩定期時ET008-c54的OD600是ET008的4.18倍。對于活細胞個數來說,在pH值為3.0時,ET008-c54和ET008差異依然不明顯;而pH值為2.5時,在穩定期時ET008-c54的活細胞數是ET008的8.07倍。這些結果表明,在pH 2.5時,隨著培養時間的增加,ET008的活細胞大量死亡,進而說明ET008-c54具有很強的耐酸性。

a-pH 3.0;b-pH 2.5
2.7 發酵實驗
ET008-c54和ET008在pH 2.5時的細胞發酵特性如圖6所示。由圖6可知,在pH值為2.5時,ET008-c54的生長速度更快,最大比生長速率(μmax)是ET008的6.38倍[ET008,μmax=(0.062±0.021) h-1);ET008-c54,μmax=(0.396±0.074) h-1](圖6-a)。此外,與ET008相比,ET008-c54最大生物量提高了2.1倍(圖6-a)。在發酵結束時,ET008-c54中的葡萄糖幾乎耗盡,而在相同的發酵時間內,ET008中只有30.5%的葡萄糖被利用(圖6-b)。正如預期,ET008-c54的乙醇產量明顯增加,是ET008的150.7%。此外,甘油作為高品質葡萄酒的重要成分[18],與ET008相比,ET008-c54的甘油含量增加218%(圖6-c)。乙酸作為不受歡迎的發酵的副產物之一[19],與ET008相比,ET008-c54中乙酸含量降低45.7%。此外,ET008-c54中的麥角甾醇的含量和H+-ATPase活性顯著增加,分別比ET008高435%和586%(圖6-d)。麥角甾醇是存在于酵母細胞中的主要甾醇,在維持細胞膜的正常功能中起著重要作用[20],例如,較高的麥角甾醇含量可有效保持最佳細胞膜厚度[21]。因此,麥角甾醇的積累可以保護細胞膜免受酸脅迫,并有助于酵母細胞的酸耐受性[22]。低pH和高葡萄糖含量對酵母細胞代謝產生不利的影響,可能導致細胞活力和發酵能力下降,這是由于在極端pH(pH<2.5)下細胞外質子濃縮,負責細胞內質子運輸H+-ATPase的壓力驟增[23]。這些研究結果表明,具有高活力H+-ATPase的酵母可在低pH和高葡萄糖條件下進行酒精發酵,這與嗜酸酵母的研究一致[24]。總之,ET008-c54的生長速率,葡萄糖消耗速率,乙醇產量和H+-ATPase活性均得到明顯改善,表明ET008-c54的發酵能力可有效利用葡萄糖并將其在低pH值下轉化為乙醇。
3 結論
本研究通過 Illumina測序平臺對耐酸釀酒酵母進行轉錄組測序分析,共獲得688個差異表達基因,其中364個差異基因上調,324個差異基因下調。通過對差異基因的分析,確定8個基因(ERG5、FET3、FIT2、FRE1、GCN1、INO1、IZH4和OPT2)是ET008-c54具有耐酸特性的關鍵基因。基于差異基因的GO富集和KEGG通路富集結果表明,關鍵基因與細胞膜的組成及生理功能、麥角固醇合成及亞鐵吸收等代謝途徑有關。進一步的細胞活力及發酵實驗也證實ET008-c54可以耐受較低的pH。本研究結果為進一步篩選高酸度工業果酒釀造菌株提供相關技術參考。
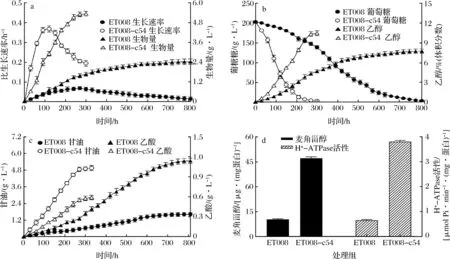
a-生長速率和生物量的變化;b-葡萄糖和乙醇含量的變化;c-甘油和乙酸含量的變化;d-麥角甾醇含量和H+-ATPase活性的測定
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
