SB431542對人羊膜上皮細胞上皮間質轉化的影響
2020-04-16 02:32:38王靜戴成祥李萍沈美萍李蘇克劉必佐
中國醫藥生物技術 2020年2期
王靜,戴成祥,李萍,沈美萍,李蘇克,劉必佐
·論著·
SB431542對人羊膜上皮細胞上皮間質轉化的影響
王靜*,戴成祥*,李萍,沈美萍,李蘇克,劉必佐
201210 上海賽比曼生物科技有限公司
探討通過添加 TGF-β 抑制劑 SB431542控制人羊膜上皮細胞的干性、表型和間質特性,為SB431542 在預防人羊膜上皮細胞上皮間質轉化(EMT)中的作用提供依據。
使用不同濃度 SB431542 培養 P2 代人羊膜上皮細胞,通過細胞的增殖曲線和基因水平(CK8、ACTA2、OCT4 和 KLF4)表達的差異,篩選出最適濃度。以不加 SB431542 的細胞為對照組,以加最適濃度的 SB431542 為實驗組,比較不同代次的對照組和實驗組的細胞倍增時間,基因表達水平(CK8、CK19、OCT4、KLF4、Vimentin、ACTA2),流式檢測細胞表面標志物(CD146、CD105、SSEA4)和免疫熒光檢測細胞蛋白表達情況(CK8、ACTA2、EpCAM)的差異。
使用不同濃度 SB431542 培養 P2 代人羊膜上皮細胞,5 μmol/L SB431542 培養條件下的細胞增殖曲線最優,且基因水平(CK8、ACTA2、OCT4 和 KLF4)表達最優,最終篩選最適藥物濃度為 5 μmol/L SB431542 完成后續實驗。在 5 μmol/L SB431542 培養條件下,P7 代實驗組的細胞倍增時間優于對照組;P7 代實驗組細胞的 CK8、CK19、KLF4、OCT4 的表達水平明顯高于對照組細胞的表達水平,實驗組的 ACTA2(α-SMA)和 Vimentin 表達水平明顯低于對照組細胞的表達水平;P3 和 P5 代細胞的實驗組的間充質標志物 CD146、CD105 的表達量均比對照組的低,實驗組干細胞標志物 SSEA4 的表達量均比對照組的高;P7 代實驗組的 CK8 和 EpCAM 表達效果強于 P7 代對照組細胞,且 P7 代實驗組的 α-SMA 基本不表達。
5 μmol/L SB431542 在人羊膜上皮細胞體外培養7 個代次以內可以有效地抑制其上皮間質轉化的發生。
人羊膜上皮細胞; TGF-β; SB431542; 上皮間質轉化; 干細胞
羊膜是位于胎兒胎盤內側的一層薄膜。人羊膜上皮細胞(human amniotic epithelial cells,hAECs)表達多種胚胎干細胞表面標志物:腫瘤排斥抗原 TRA1-60、TRA1-81,階段性特異性抗原 SSEA3、SSEA4,多潛能轉錄因子 NANOG、OCT4、SOX2[1]。這些因子相互協調,在維持胚胎干細胞的自我更新和多向分化潛能方面發揮著關鍵作用[2]。目前,人羊膜上皮細胞已在結膜重建、神經系統疾病、肝臟疾病等動物疾病模型中取得初步成效[3-6]。然而,要實現人羊膜上皮細胞的臨床應用,仍有許多待解決的問題,特別是上皮間質轉化的發生。人羊膜上皮細胞在體外擴增過程中由于自發經歷了上皮間質轉化(EMT),其自身會發生性狀的改變[7]。EMT是上皮細胞獲得間充質表型的一個轉分化過程。上皮細胞通常呈頂基極性,通過黏附連接和緊密連接表現出較強的細胞-細胞黏附性,基底基質主要由 IV 型膠原和層粘連蛋白組成[8]。EMT 誘導發生后,上皮細胞失去黏附性,細胞形態發生改變,并獲得了頂端到尾端的極性[8-9]。發生 EMT 的上皮細胞分裂并侵入基底膜,沿著纖維連接蛋白和 I 型膠原新形成的基質遷移[9]。上皮細胞標志物的豐度開始降低,而間質標志物的豐度開始增加。細胞通路信號,如 TGF-β、Wnt 和 FGF 可以抑制 EMT 以及促進它的逆向過程 MET 的發生[10]。EMT 過程將會導致上皮細胞相關蛋白(E-Cadherin、Cytokeratin-8)表達的丟失,上調間充質相關蛋白[如波形蛋白(Vimentin)、α 平滑肌肌動蛋白(α-SMA)]的表達[10-11],特別是 EMT 顯著改變了人羊膜上皮細胞的生物學特性和再生潛力[12-13]。人羊膜上皮細胞 EMT 是一種依賴于 TGF-β 的現象,使用一種 TGF-β 受體抑制劑 SB431542 可以阻止 EMT的發生并保留人羊膜上皮細胞的上皮表型。SB431542 是一種 TGF-β I 型受體激活素受體樣激酶(ALK)5、ALK 4 和 ALK 7 的抑制劑,可以誘導 Smad2/Smad4 和 Smad3/Smad4 依賴的轉錄,但對 BMP 信號通路沒有影響[14]。在 A498 細胞中,SB431542 抑制TGF-β I 誘導的膠原 Iα1 和 PAI-1 mRNA。此外,SB431542 抑制 TGF-β I 誘導的纖連蛋白 mRNA 和蛋白的表達[15]。SB431542 抑制 TGF-β 調節的生長因子,包括 PDGF-A、FGF-2 和 HB-EGF[16]。在 NMuMG 和 PANC-1 細胞中,SB431542 也抑制 TGF-β 調節的上皮細胞向間質轉型[17]。
1 材料和方法
1.1 材料
1.1.1 試劑與耗材 人足月胎盤經知情同意情況下獲得,符合倫理要求。上皮細胞培養基(EpiCM)購于美國 Sciencell 公司;10 × TrypLE、RevertAid? First Strand cDNA Synthesis Kit、TaqMan?Gene Expression assays、α-SMA 多克隆抗體、CK8 單克隆抗體均購于美國 Thermos 公司;SB431542 購于美國 Selleck 公司;RNAeasy? Plus 動物 RNA 抽提試劑盒(離心柱式)、CCK8 均購于上海碧云天生物技術有限公司;CD146-PE、CD324-FITC、SSEA4-FITC、CD105-APC 均購于美國 Biolegend 公司;DMSO 購于德國 WAK 公司;青鏈霉素購于德國 PAN Biotech 公司。
1.2 方法
1.2.1 人羊膜上皮細胞的分離 將胎盤的胎兒面朝上放置于紫外消毒生物安全柜,使用手術刀劃“十”字,四等分羊膜,使用無菌手術鑷、手術剪剝取胎盤表面的羊膜。第一次消化:①將羊膜置于 10 cm 直徑培養皿,培養皿中裝有 1/2 體積的含有抗生素(1% 青鏈霉素)的PBS。②用手術鑷夾持羊膜進行漂洗,重復漂洗 2 ~ 3 遍,去除血細胞。③轉移羊膜至新的 10 cm 直徑培養皿中,添加 10 × TrypLE(10 ml/g 組織),將組織放置于恒溫搖床中,37 ℃,100 r/min 消化0.5 h。④消化完畢,將組織及消化液轉入 15 cm 直徑的培養皿中,使用細胞刮鏟,輕刮細胞。⑤將羊膜組織轉入新的 10 cm 直徑培養皿中,用于第二次消化。細胞懸液轉入50 ml 離心管中,以 4 ℃、300 ×條件離心10 min。⑥棄去上清液,使用 20 ml PBS 重懸沉淀,4 ℃保存。第二次消化:①在 10 cm 直徑培養皿的羊膜組織中,添加 10 × TrypLE 20 ml,將組織放置于恒溫搖床中,37 ℃、100 r/min 消化0.5 h。②消化完畢,將組織及消化液轉入 15 cm 直徑的培養皿中,使用細胞刮鏟,輕刮細胞。③細胞懸液轉入 50 ml 離心管中,以 4 ℃、300 ×條件離心10 min。④棄去上清液,使用 20 ml PBS 重懸沉淀,4 ℃保存。⑤將第一次和第二次消化的細胞懸液混勻,臺盼藍計數,按照 2 × 104/cm2接種 T75 培養瓶,37 ℃體積分數 5% CO2培養。⑥ 48 h 換液,以后每 2 天換液 1 次,直至細胞可以進行傳代。
1.2.2 人羊膜上皮細胞的原代培養 細胞接種密度為 2 × 104/cm2,37 ℃體積分數 5% CO2培養。使用未添加 SB431542 的上皮完全培養基進行培養。上皮完全培養基的配制按照EpiCM 的使用說明進行。人羊膜上皮細胞的傳代培養:人羊膜上皮細胞生長至 90% 時,每個 T75 培養瓶加入 2 ml TrypLE 進行消化傳代。消化 2 min,加未加藥物的上皮完全培養基終止消化,將細胞懸液轉移至50 ml 離心管中,1200 r/min 離心 5 min,棄上清,計數,按照 1 × 104/cm2接種,置于 37 ℃體積分數 5% CO2培養。傳代培養基:上皮完全培養基中添加不同濃度的 SB431542。
1.2.3 篩選最適濃度 SB431542 ①將 SB431542 溶于 DMSO,濃度配制成 10 mmol/L。②將 10 mmol/L SB431542 加入至上皮細胞完全培養基中,分別配制成 0、1、5、10、50 μmol/L 濃度。③取 P2 代人羊膜上皮細胞,使用不同藥物濃度的培養基進行培養,每 2 天換一次加藥培養基。④使用 CCK8,對培養 1、2、3、4 d 的 P2 代細胞進行生長曲線測定。⑤使用 qPCR 檢測培養 3 d 的 P2 代細胞的 CDH1、CK8、OCT4、KLF4、ACTA2(α-SMA)的基因表達情況。⑥篩選出最適的藥物濃度完成后續實驗。
1.2.4 細胞生長曲線測定 取不添加 SB431542 的上皮完全培養基培養的 P1 代人羊膜上皮細胞,經 TrypLE 消化,以 1 × 104/cm2的密度接種于96 孔板,每個藥物濃度的細胞接種 2 孔。以對應的藥物濃度的完全培養基作為空白對照,接種2 孔,SB431542 的濃度分別為 0、1、5、10、50 μmol/L。使用 CCK8 試劑盒測試在不同濃度 SB431542 的培養條件下,人羊膜上皮細胞在 1、2、3、4 d 的代謝值,并篩選出細胞代謝最高值對應的藥物濃度。
1.2.5 細胞倍增時間測定 取不添加 SB431542 的上皮完全培養基培養的 P1 代人羊膜上皮細胞,經 TrypLE 消化,以 1 × 104/cm2的密度接種于T25 培養瓶。對照組細胞使用不加 SB431542 的上皮完全培養基,實驗組細胞使用最適濃度的 SB421542 的上皮完全培養基進行培養,等細胞90% 融合度,進行傳代,標記為 P2 代細胞,以此類推,計算出 P1 ~ P7 代每個代次的人羊膜上皮細胞的倍增時間。細胞倍增時間計算:DT = t×[lg2/(lgNt – lgN0)](t:培養時間;N0 為接種細胞數,Nt 為 t 時間后的細胞數)。
1.2.6 熒光定量 PCR(qPCR)檢測 使用 RNA 抽提試劑盒抽提 RNA,使用 RevertAid? First Strand cDNA Synthesis Kit 反轉錄 1 μg RNA。根據實驗要求,使用 TaqMan?Gene Expression assays 中的探針為引物(表 1),熒光定量 PCR 檢測相關轉錄因子 mRNA 表達量。并以 GAPDH 為內參,實驗重復 3 次。PCR 結束后收集 Ct 值,使用 2-??CT法對實驗數據進行分析。

表 1 熒光定量 PCR 引物設計
注:使用TaqMan Gene Expression assays進行 qPCR 分析,評估相關轉錄因子的 mRNA 表達水平情況。
Note: TaqMan Gene Expression assays were used for qPCR analysis to evaluate mRNA expression levels of related transcription factors.
1.2.7 免疫熒光鑒定羊膜上皮細胞 ①取出12 孔板,將涂有多聚賴氨酸的細胞爬片置于孔內;②以不加藥物培養的 P1、P3、P7 人羊膜上皮細胞作為陰性對照,以最適濃度 SB431542 培養的 P1、P3、P7 人羊膜上皮細胞作為實驗組,以 1 × 104/cm2密度接種;③細胞爬片按以下步驟進行染色:PBS 漂洗 3 次,細胞經 4% 多聚甲醛溶液室溫固定20 min;PBS 漂洗 3 次,使用 0.5% Triton X-100 進行破膜;加入 500 μl 封閉液(PBS 含0.1% 的BSA 稀釋)室溫封閉30 min;分別加入一抗 CK8、EpCAM、α-SMA 在 4 ℃條件下孵育過夜;PBS 漂洗 3 次,加入二抗,室溫避光孵育 1 h;PBS 漂洗 3 次,加入1 μg/ml 的DAPI 500 μl,孵育10 min。漂洗、晾干、封固后熒光顯微鏡下觀察。
1.2.8 流式檢測人羊膜上皮細胞 ①以不加藥物培養的 P3、P5、P7 人羊膜上皮細胞作為陰性對照,以最適濃度 SB431542 培養的 P3、P5、P7 人羊膜上皮細胞作為實驗組。收集細胞懸液,1500 r/min 離心 5 min,棄上清(留 100 μl 左右液體)。②加抗體(CD324、CD146、CD105、SSEA4),避光 20 min孵育。③加 1 ml/管 PBS,1500 r/min 離心 5 min,棄上清。④加 500 μl PBS,樣品用于上機。
1.3 統計學處理
使用 GraphPad Prism 5.0 對數據進行分析。
2 結果
2.1 5 μmol/L SB431542 為最適藥物濃度
使用 CCK8 檢測 P2 代人羊膜上皮細胞在不同濃度藥物影響下每天的代謝450值。從實驗數據來看(圖 1A),50 μmol/L SB431542 對細胞的增殖具有顯著抑制作用,低劑量 SB431542(0、1、5、10 μmol/L)對于細胞的增殖沒有抑制作用。
使用 qPCR 檢測不同濃度藥物影響的 P2 代人羊膜上皮細胞的基因表達水平情況。使用的Marker 為上皮標志物CK8、間充質標志物 ACTA2(α-SMA)、干細胞特性標志物 OCT4 和 KLF4。從實驗數據來看(圖 1B),5 μmol/L SB431542 和對照組 0 μmol/L SB431542 在 CK8 和 KLF4 的基因表達水平上具有顯著性差異(< 0.05)。
綜上,挑選最適藥物濃度為 5 μmol/L SB431542完成后續對不同代次人羊膜上皮細胞的上皮表型影響的實驗。
2.2 5 μmol/L SB431542 對人羊膜上皮細胞的增殖影響
記錄每個代次 90% 融合度的細胞數和時間(h),計算每個代次細胞倍增所需要的時間。對照組為不加藥的細胞,實驗組為 5 μmol/L SB431542 處理的細胞。
實驗數據(圖 2)表明,P1 ~ P6 人羊膜上皮細胞加藥和不加藥的細胞倍增時間基本一致。在 P7 代,加藥的細胞倍增時間為 43.5 h,不加藥的細胞倍增時間為 61.4 h。5 μmol/L SB431542 在7 個代次內可維持羊膜上皮細胞的增殖。
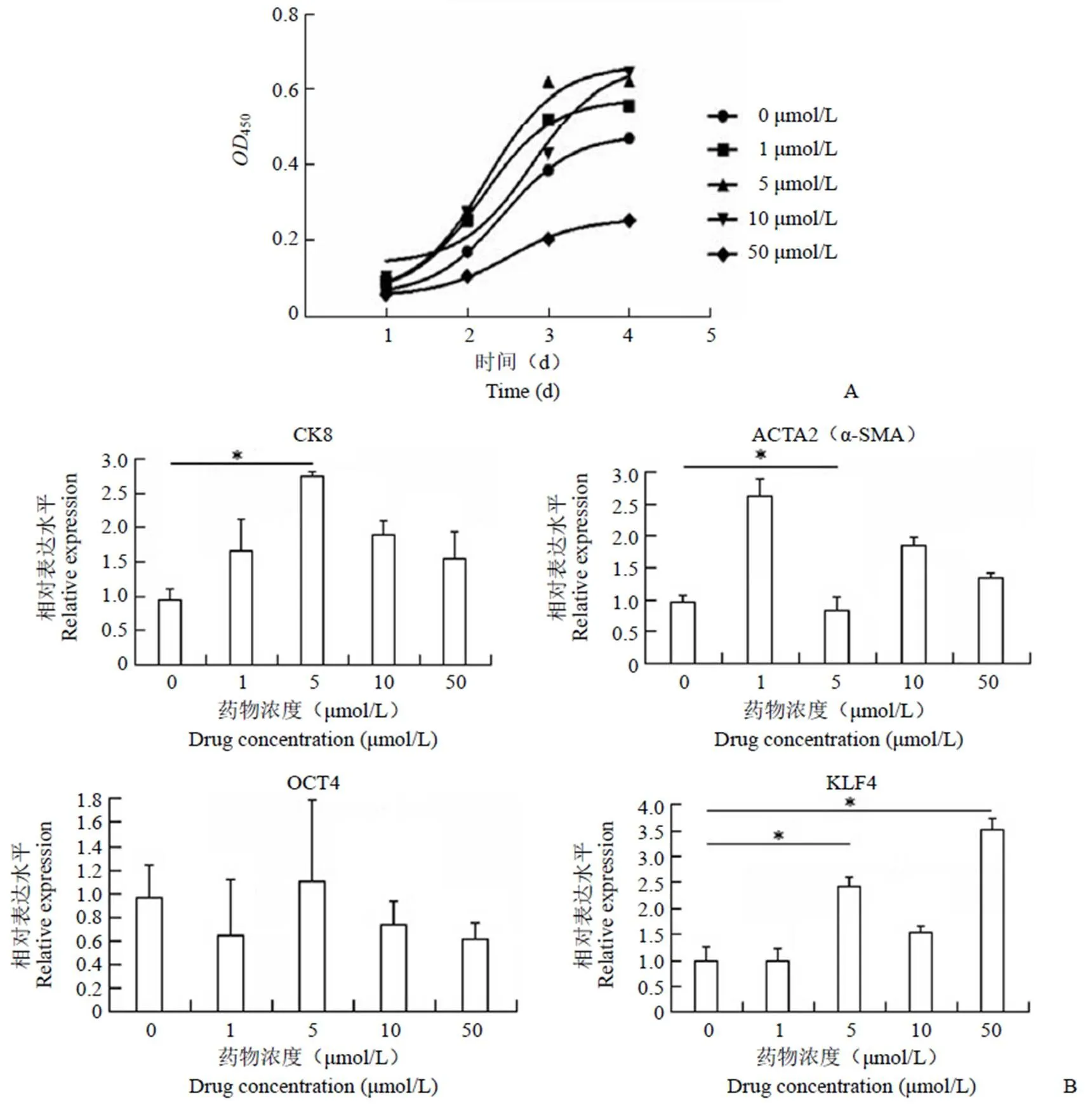
圖 1 5 μmol/L SB431542 為 hAECs 生長的最佳的濃度(A:每天將 CCK8 加入不同濃度 SB431542 培養的細胞中,根據OD450讀數繪制生長曲線;B:使用 qPCR 檢測不同濃度 SB431542 培養的 P2 hAECs 的生物標志物的表達水平情況;*P < 0.05)
Figure 1 5 μmol/L of SB431542 was considered as the optimal concentration [A: The growth curve was drawn based on the450read after CCK8 was added to cells cultured with different concentration of SB431542 each day; B: The expression of biological markers including cytokeratin 8 (CK8), mesenchymal marker ACTA2 (α-SMA), and stem cell markers OCT4 and KLF4 were assessed by qPCR analysis in P2 hAECs treated with SB431542 at different concentrations;*< 0.05]
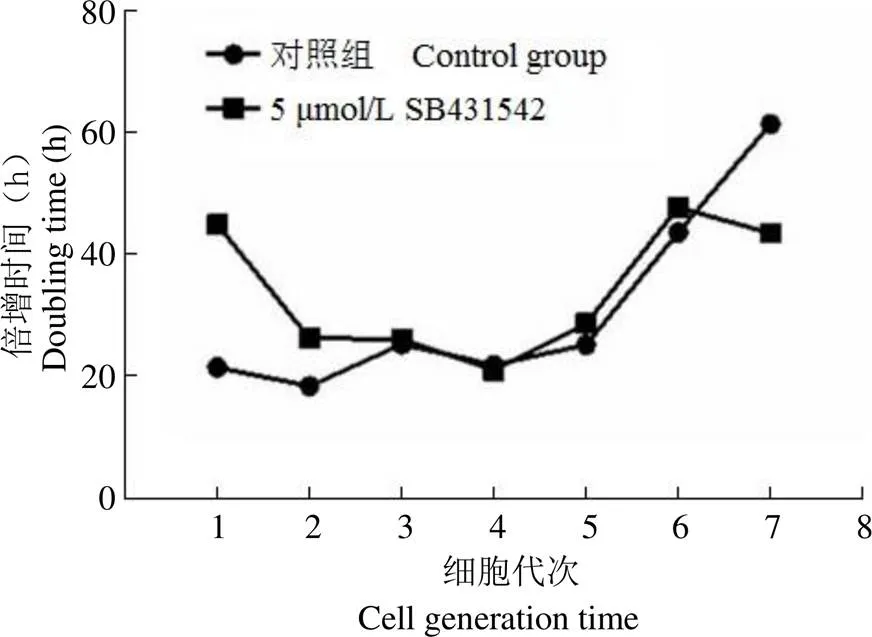
圖 2 5 μmol/L SB431542 對不同代次的人羊膜上皮細胞的倍增時間影響
Figure 2 Effect of SB431542 on doubling time of hAECs at different generation
2.3 5 μmol/L SB431542 對人羊膜上皮細胞的基因表達水平的影響
使用 qPCR 檢測對照組和實驗組 P1、P3、P7 代人羊膜上皮細胞的基因表達水平情況。使用的Marker 為上皮標志物角蛋白 8(CK8)和 19(CK19)、干細胞標志物 OCT4 和 KLF4、間充質干細胞標志物波形蛋白和 ACTA2(α-SMA)。從實驗數據(圖 3)來看,對照組和實驗組在 CK8、CK19、KLF4、OCT4、波形蛋白和 ACTA2(α-SMA)的基因表達水平上都有顯著性差異(< 0.05)。且在 P7 代時,實驗組細胞的 CK8、CK19、KLF4、OCT4 的表達水平明顯高于對照組細胞的表達水平;實驗組細胞的 ACTA2(α-SMA)和波形蛋白明顯低于對照組的表達水平。

圖 3 使用 qPCR 方法檢測對照組和實驗組的 P1、P3、P7 hAECs 基因表達情況(*P < 0.05,**P < 0.01,***P < 0.001)
Figure 3 The qPCR analysis was performed to assess gene expression in the P1, P3 and P7 hAECs of the control and experimental groups (*< 0.05,**< 0.01,***< 0.001)
2.4 5 μmol/L SB431542 對人羊膜上皮細胞的表面標志物的影響
流式檢測技術檢測對照組和實驗組 P3、P5、P7 代人羊膜上皮細胞的表面標志物的變化情況。使用的 Marker 為間充質標志物 CD146、CD105 和干細胞標志物 SSEA4。從實驗數據(表 2)來看,P3 和 P5 代細胞的對照組的間充質標志物 CD146、CD105 的表達量都要比實驗組的高,對照組 SSEA4 的表達量都要比實驗組的低。
2.5 5 μmol/L SB431542 對人羊膜上皮細胞的蛋白表達的影響
免疫熒光檢測 P1 和 P7 代人羊膜上皮細胞的表面標志物的變化情況。使用的 Marker 為上皮細胞標志物CK8 和 EpCAM、間充質干細胞標志物 α-SMA。從實驗結果來看(圖 4),P1 代實驗組的 CK8 和 EpCAM 表達效果強于 P1 代對照組細胞,α-SMA 都為弱表達;P7 代實驗組的 CK8 和 EpCAM 表達效果強于 P7 代對照組細胞,且 P7 代實驗組的 α-SMA 基本不表達。
3 討論
本研究證實了 TGF-β 抑制劑 SB431542 在人羊膜上皮細胞體外擴增過程中維持其上皮表型、保持干性的作用。新鮮分離的人羊膜上皮細胞表達高水平的上皮標志物 CK8 和低水平的間充質標志物 α-SMA。然而,隨著傳代次數的增加,CK8 的表達量減少,α-SMA 的表達量增加。α-SMA 是EMT 過程中蛋白組成的一部分[18]。由于 CK8 和 α-SMA 在 EMT 和 MET 過程中具有快速的調節作用,因此被選為首選的 EMT 標記物[19]。KLF4 在 EMT 過程中也被鑒定為 TGF-β 調控基因[20-21]。人羊膜上皮細胞具有較強的KLF4 表達,隨著傳代次數的增加,KLF4 表達逐漸減少,最終消失。波形蛋白的存在也被認為是細胞發生 EMT 的一個重要標志[22]。此外,在上皮細胞向間質轉化的過程中,波形蛋白的表達已被證明會誘導細胞的表型、活力和細胞黏附能力發生改變[23]。

表 2 5 μmol/L SB431542 對于不同代數人羊膜上皮細胞的表面標志物影響
注:使用流式檢測方法,觀察對照組和實驗組中 P3、P5、P7 hAECs 相關表面標志物的變化情況。
Note: The changes of surface markers of P3, P5 and P7 of hAECs between control group and experimental group were observed by flow cytometry.
在 P7 代時,使用 SB431542 處理過的人羊膜上皮細胞的 CK8、CK19、KLF4、OCT4 的基因表達水平高于對照組細胞的表達水平;實驗組細胞的 ACTA2(α-SMA)和波形蛋白的基因表達水平低于對照組的表達水平。從實驗數據可以看出,SB431542 在基因水平上有抑制人羊膜上皮細胞的 EMT 的發生,同時不影響人羊膜上皮細胞的干細胞標志物的表達。使用 SB431542 處理過的P3、P5 代人羊膜上皮細胞的間充質表面標志物CD146、CD105 的表達量都要比對照組的低,而 SSEA4 的表達量都要比對照組的高。實驗結果表明,SB431542 在蛋白表達水平上能抑制人羊膜上皮細胞的 EMT 的發生,但同時促進干細胞特性相關蛋白 SSEA4 的表達。P1 代實驗組的 CK8 和 EpCAM 表達效果強于 P1 代對照組細胞,α-SMA 都為弱表達;P7 代實驗組的 CK8 和 EpCAM 表達效果強于 P7 代對照組細胞,且 P7 代實驗組的 α-SMA 基本不表達。使用 SB431542 處理后的人羊膜上皮細胞在 P7 代仍能夠形成緊密的“鋪路石”上皮樣,但是對照組的細胞呈間充質特點的“長梭形”。

圖 4 5 μmol/L SB431542 對于不同代次羊膜上皮細胞的蛋白表達情況影響(對照組:未經 SB431542 處理的 hAECs;實驗組:使用 5 μmol/L SB431542 處理過的 hAECs;熒光標記:綠色;DAPI:藍色;標尺:50 μm)
Figure 4 Effect of 5 μmol/L SB431542 on protein expression in different generations of hAECs (Control group: hAECs without SB431542 treatment; Experimental group: hAECs treated by 5 μmol/L SB431542; Markers: Green; DAPI: Blue; Scale bar: 50 μm)
TGF-β 對細胞的生長、分化和免疫功能都有重要的調節作用。TGF-β 能夠促進成纖維細胞的增殖[24],抑制上皮細胞、內皮細胞、免疫細胞的增殖[25]。同時 TGF-β 在治療傷口愈合[26],促進軟骨和骨修復[27]以及通過免疫抑制治療自身免疫性疾病[28]等方面有潛在的應用前景。在本研究中使用了 TGF-β 抑制劑 SB431542 處理了人羊膜上皮細胞,但其在體內的生物學安全性仍有待后續研究。
[1] Miki T, Lehmann T, Cai H, et al. Stem cell characteristics of amniotic epithelial cells. Stem Cells, 2005, 23(10):1549-1559.
[2] Kellner S, Kikyo N. Transcriptional regulation of the Oct4 gene, a master gene for pluripotency. Histol Histopathol, 2010, 25(3):405- 412.
[3] Tan JL, Lau SN, Leaw B, et al. Amnion epithelial cell-derived exosomes restrict lung injury and enhance endogenous lung repair. Stem Cells Transl Med, 2018, 7(2):180-196.
[4] Rodriguez NS, Yanuaria L, Parducho KMR, et al. Liver-directed human amniotic epithelial cell transplantation improves systemic disease phenotype in hurler syndrome mouse model. Stem Cells Transl Med, 2017, 6(7):1583-1594.
[5] Wu Z, Hui G, Lu Y, et al. Human amniotic epithelial cells express specific markers of nerve cells and migrate along the nerve fibers in the corpus callosum. Neural Regen Res, 2012, 7(1):41-45.
[6] Yang SP, Yang XZ, Cao GP. Conjunctiva reconstruction by induced differentiation of human amniotic epithelial cells. Genet Mol Res, 2015, 14(4):13823-13834.
[7] Lange-Consiglio A, Accogli G, Cremonesi F, et al. Cell surface glycan changes in the spontaneous epithelial-mesenchymal transition of equine amniotic multipotent progenitor cells. Cells Tissues Organs, 2014, 200(3-4):212-226.
[8] Gonzalez DM, Medici D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci Signal, 2014, 7(344):re8.
[9] Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial- mesenchymal transition. Nat Rev Mol Cell Biol, 2014, 15(3):178-196.
[10] Zhang J, Tian XJ, Xing J. Signal transduction pathways of EMT induced by TGF-β, SHH, and WNT and their crosstalks. J Clin Med, 2016, 5(4):E41.
[11] Medici D, Hay ED, Olsen BR. Snail and Slug promote epithelial-mesenchymal transition through beta-catenin-T-cell factor-4-dependent expression of transforming growth factor-beta3. Mol Biol Cell, 2008, 19(11):4875-4887.
[12] Barboni B, Russo V, Curini V, et al. Gestational stage affects amniotic epithelial cells phenotype, methylation status, immunomodulatory and stemness properties. Stem Cell Rev Rep, 2014, 10(5):725-741.
[13] Stadler G, Hennerbichler S, Lindenmair A, et al. Phenotypic shift of human amniotic epithelial cells in culture is associated with reduced osteogenic differentiation in vitro. Cytotherapy, 2008, 10(7):743-752.
[14] Inman GJ, Nicolás FJ, Callahan JF, et al. SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol Pharmacol, 2002, 62(1):65-74.
[15] Laping NJ, Grygielko E, Mathur A, et al. Inhibition of transforming growth factor (TGF)-beta1-induced extracellular matrix with a novel inhibitor of the TGF-beta type I receptor kinase activity: SB-431542. Mol Pharmacol, 2002, 62(1):58-64.
[16] Matsuyama S, Iwadate M, Kondo M, et al. SB-431542 and Gleevec inhibit transforming growth factor-beta-induced proliferation of human osteosarcoma cells. Cancer Res, 2003, 63(22):7791-7798.
[17] Halder SK, Beauchamp RD, Datta PK. A specific inhibitor of TGF-beta receptor kinase, SB-431542, as a potent antitumor agent for human cancers. Neoplasia, 2005, 7(5):509-521.
[18] Kalluri R, Neilson EG. Epithelial-mesenchymal transition and its implications for fibrosis. J Clin Invest, 2003, 112(12):1776-1784.
[19] Martinovic V, Vukusic Pusic T, Restovic I, et al. Expression of epithelial and mesenchymal differentiation markers in the early human gonadal development. Anat Rec (Hoboken), 2017, 300(7):1315-1326.
[20] Valcourt U, Kowanetz M, Niimi H, et al. TGF-beta and the Smad signaling pathway support transcriptomic reprogramming during epithelial-mesenchymal cell transition. Mol Biol Cell, 2005, 16(4): 1987-2002.
[21] Li XM, Kim SJ, Hong DK, et al. KLF4 suppresses the tumor activity of cutaneous squamous cell carcinoma (SCC) cells via the regulation of SMAD signaling and SOX2 expression. Biochem Biophys Res Commun, 2019, 516(4):1110-1115.
[22] Janda E, Lehmann K, Killisch I, et al. Ras and TGF[beta] cooperatively regulate epithelial cell plasticity and metastasis: dissection of Ras signaling pathways. J Cell Biol, 2002, 156(2):299-313.
[23] Mendez MG, Kojima S, Goldman RD. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J, 2010, 24(6):1838-1851.
[24] de Larco JE, Todaro GJ. Growth factors from murine sarcoma virus-transformed cells. Proc Natl Acad Sci U S A, 1978, 75(8):4001- 4005.
[25] Moses HL, Roberts AB, Derynck R. The discovery and early days of TGF-β: a historical perspective. Cold Spring Harb Perspect Biol, 2016, 8(7):a021865.
[26] Roberts AB. Transforming growth factor-beta: activity and efficacy in animal models of wound healing. Wound Repair Regen, 1995, 3(4): 408-418.
[27] Poniatowski ?A, Wojdasiewicz P, Gasik R, et al. Transforming growth factor Beta family: insight into the role of growth factors in regulation of fracture healing biology and potential clinical applications. Mediators Inflamm, 2015, 2015:137823.
[28] Letterio JJ, Roberts AB. Regulation of immune responses by TGF-beta. Annu Rev Immunol, 1998, 16:137-161.
Effect of SB431542 on epithelial mesenchymal transformation of human amniotic epithelial cells
WANG Jing, DAI Cheng-xiang, LI Ping, SHEN Mei-ping, LI Su-ke, LIU Bi-zuo
Cellular Biomedicine Group, Shanghai 201210, China
SB431542 was added to control the stemness, phenotype and mesenchymal characteristics of human amniotic epithelial cells. The objective of this study is to provide evidence for the role of TGF-β inhibitor SB431542 in preventing epithelial mesenchymal transformation of human amniotic epithelial cells.
Different concentrations of SB431542 was used to culture P2 human amniotic epithelial cells, and the optimal concentration was selected through cell proliferation curve and gene expression (CK8, ACTA2, OCT4 and KLF4). Cells without SB431542 were used as the control group, while cells with SB431542 were experimental group. Cells doubling time, gene expression levels (CK8, CK19, OCT4, KLF4, Vimentin, ACTA2), the mesenchymal biomarkers (CD146, CD105, SSEA4) and epithelial biomarker levels (CK8, ACTA2, EpCAM) were detected by cellular number accounting, qPCR, flow cytometry and immunofluorescence, respectively, and compared between the control group and the experimental group.
When cultured P2 human amniotic epithelial cells were treated with different concentration of SB431542, the cell proliferation and the gene expression (CK8, ACTA2, OCT4 and KLF4) were the best after treatment with 5 μmol/L SB431542. Therefore, 5 μmol/L SB431542 was used in the following experiments. Under the culture condition of 5 μmol/L SB431542, the doubling time of P7 cell group was better than that of control group. Also, the gene expression levels of CK8, CK19, KLF4, OCT4 in P7 cell group with 5 μmol/L SB431542 were significantly higher than that in control group, while ACTA2 (α-SMA) and Vimentin in P7 cell group with 5 μmol/L SB431542 were significantly lower than that in control group. Expression levels of mesenchymal markers CD146, CD105 in P3 and P5 cells group with 5 μmol/L SB431542 were lower, while SSEA4 was higher, than that in the control group. The expressions of epithelial markers CK8 and EpCAM in the P7 cell group with 5 μmol/L SB431542 were higher than that in the P7 control group, while the α-SMA in the P7 experimental group was undetectable.
5 μmol/L SB431542 can inhibit the occurrence of epithelial mesenchymal transformation effectively in human amniotic epithelial cells culturedwithin 7 generations.
Human amniotic epithelial cells; TGF-β; SB431542; Epithelial mesenchymal transformation; Stem cells
·協會之窗·
中國醫藥生物技術協會積極配合多部委開展新冠肺炎干細胞治療應急科研課題備案
新型冠狀病毒感染作為一個全新的疾病,目前對其診治還有很多處于摸索階段,為此部分醫療機構提出了開展干細胞治療新冠肺炎臨床研究的課題,科技部也啟動了相關的應急科研立項。為了加快相關干細胞臨床研究的備案,中國醫藥生物技術協會作為干細胞臨床研究備案技術審核單位,積極配合國家衛生健康委、國家藥監局、科技部,開通干細胞治療新冠肺炎臨床研究備案審核的快速通道,組織專家提前介入,完善研究方案,協助相關課題的備案工作。
中國醫藥生物技術協會關于設立新分支機構的公告
根據《民政部關于貫徹落實國務院取消全國性社會團體分支機構、代表機構登記行政審批項目的決定有關問題的通知》(民發[2014]38 號)和《中國醫藥生物技術協會分支機構管理辦法》等有關規定,由有關專家和機構發起,經協會第六屆理事會第二次理事會議審議通過,決定設立“中國醫藥生物技術協會臨床前評價技術專業委員會”、“中國醫藥生物技術協會臨床研究專業委員會”、“中國醫藥生物技術協會神經修復與再生分會”、“中國醫藥生物技術協會醫工結合分會”等4 個新分支機構。
LI Ping, Email: freezonli@aliyun.com
10.3969/j.issn.1673-713X.2020.02.013
李萍,Email:freezonli@aliyun.com
2019-09-17
*同為第一作者
