滇龍膽8-羥香葉醇氧化還原酶基因的克隆與表達分析
2020-04-17 08:58:20王偉妍李彩霞李湫鄢秋紫張曉東
廣西植物 2020年2期
關鍵詞:分析
王偉妍 李彩霞 李湫 鄢秋紫 張曉東

摘要:? 滇龍膽主要藥效成分為龍膽苦苷,而8-羥香葉醇氧化還原酶基因Gr8HGO是龍膽苦苷生物合成途徑的結構基因。為了研究Gr8HGO基因的功能,該文克隆了滇龍膽Gr8HGO基因,并進行表達分析。結果表明:(1)共克隆到5個Gr8HGO基因,其GenBank登錄號分別為KP722029.1(Gr8HGO-1)、KP722030.1(Gr8HGO-2)、KP722031.1(Gr8HGO-3)、KP722032.1(Gr8HGO-4)、KP723852.1(Gr8HGO-5)。(2)Gr8HGO-1基因全長1 062 bp,編碼353個氨基酸,其他4個基因全長1 131 bp,編碼376個氨基酸;理化性質分析結果表明5個蛋白單體相對分子質量約40 kD,理論等電點在5.47~5.95之間,均為疏水穩定蛋白。(3)信號序列分析結果表明5個蛋白均不含信號肽、跨膜螺旋和葉綠體轉運肽;亞細胞定位分析結果表明5個蛋白可能定位于細胞質;結構域預測結果表明除Gr8HGO-1蛋白僅包含乙醇脫氫酶N端結構域(IPR013154)和C端結構域(IPR013149)外,其他4個蛋白還包含聚酮合酶、烯酰還原酶結構域(IPR020843)。(4)系統發育分析結果表明這些Gr8HGO蛋白與長春花Cr8HGO蛋白親緣關系最近。(5)qPCR結果表明Gr8HGO基因主要在葉中表達,在根和莖中表達量很低。該研究為后續龍膽苦苷生物合成途徑的解析奠定基礎。
關鍵詞: 滇龍膽, 8-羥香葉醇氧化還原酶, 克隆, 生物信息學, 基因表達
中圖分類號:? Q786文獻標識碼:? A
文章編號:? 1000-3142(2020)02-0200-10
Abstract:? Gentiopicroside is the main active ingredient in Gentiana rigescens, while 8-hydroxygeraniol oxidoreductase gene Gr8HGO is a structural gene involved in gentiopicroside biosynthesis. In order to study the function of Gr8HGO gene, the Gr8HGO gene in G. rigescens was cloned and its expression analysis was conducted in this study. The results were as follows: (1) Five Gr8HGO genes were cloned and their GenBANK accession numbers were KP722029.1 (Gr8HGO-1), KP722030.1 (Gr8HGO-2), KP722031.1 (Gr8HGO-3), KP722032.1 (Gr8HGO-4) and KP723852.1 (Gr8HGO-5), separately. (2) The length of Gr8HGO-1 gene was 1 062 bp encoding 353 amino acids, while the other four genes were 1 131 bp encoding 376 amino acids; The results from physicochemical analysis showed that the relative molecular weight of these five Gr8HGO proteins were approximately 40 kD, and their theoretical pI ranged from 5.47 to 5.95, which were all hydrophobic stable proteins. (3) Signal sequence analysis showed that five proteins did not contain signal peptides, transmembrane helixes and chloroplast transit peptides; Subcellular localization analysis indicated that these five proteins might be localized in cytoplasm; Domain prediction results showed that beside the Gr8HGO-1 contained only alcohol dehydrogenase N-terminal (IPR013154) and C-terminal (IPR013149) conserved domains, the other four also contained polyketide synthase, enoylreductase domain (IPR020843). (4) Phylogenic analysis showed that these five Gr8HGO proteins had the closest relationship with Cr8HGO in Catharanthus roseus. (5) The results of qPCR suggested that Gr8HGO gene was mainly expressed in leaves, but very low in roots and stems. This study will lay a foundation for further analysis of the biosynthesis pathway of gentiopicroside.
Key words: Gentiana rigescens, 8-hydroxygeraniol oxidoreductase, cloning, bioinformatics, gene expression
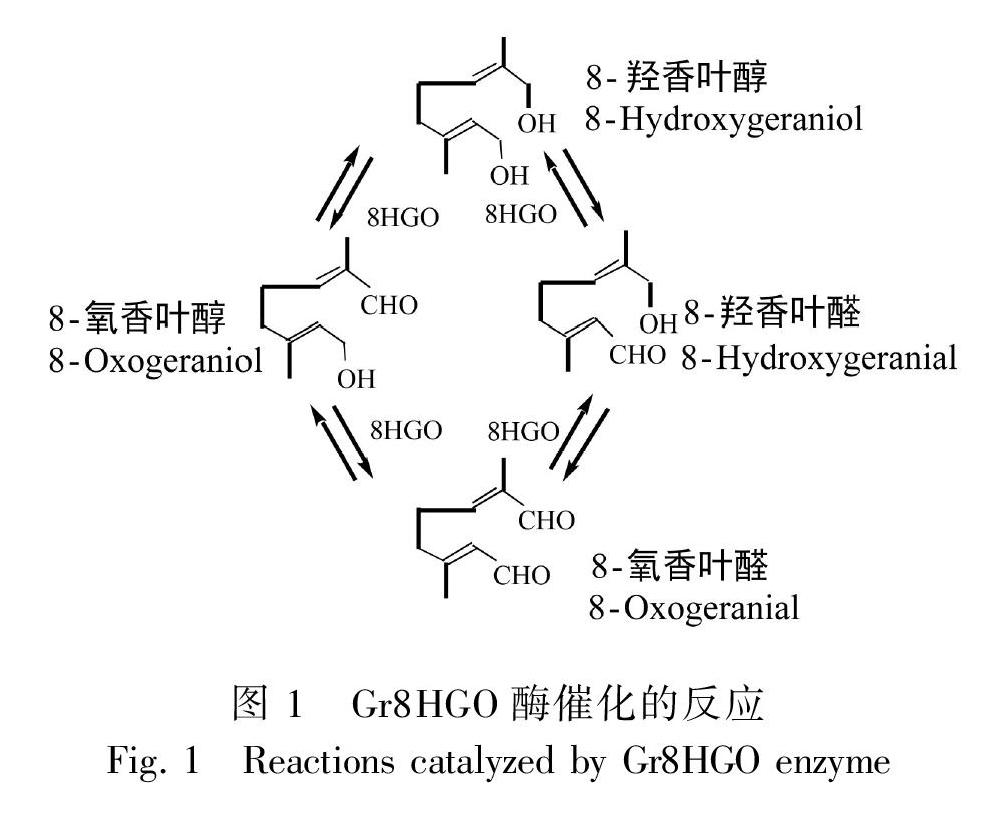
滇龍膽(Gentiana rigescens)為龍膽科(Gen-tianaceae)龍膽屬(Gentiana)植物,主要分布于我國西南地區的云南、四川、貴州等地(中國植物志編委會, 1988)。在傳統中藥中,滇龍膽藥用部位為根,具有清熱燥濕、瀉肝膽火的作用(國家藥典委員會, 2015)。現代研究表明滇龍膽根部主要藥用成分為一種單萜類化合物龍膽苦苷(Zhang et al., 2015; 張曉東等, 2016)。在植物中,龍膽苦苷的生物合成可分為三個階段。第一階段是由質體2-C-甲基-D-赤蘚糖醇-4-磷酸(MEP)途徑和胞質甲羥戊酸(MVA)途徑合成原料異戊二烯焦磷酸(IPP)和二甲基丙烯基焦磷酸(DMAPP)(Hua et al., 2014);第二階段是以IPP和DMAPP為原料,經過縮合、氧化、還原、糖基化、甲基化等步驟,生成裂環番木鱉苷(Miettinen et al., 2014; Munkert et al., 2015);第三階段是是萜類化合物經過修飾形成龍膽苦苷(黃璐琦和劉昌孝, 2015)。8-羥香葉醇氧化還原酶(8-hydroxygeraniol oxidoreductase,EC 1.1.1.324,在已報道文獻中簡寫為8HGO、10HGO)是龍膽苦苷生物合成途徑第二階段中的第四個催化酶,能夠催化8-羥香葉醇、8-羥香葉醇、8-氧香葉醛和8-氧香葉醛之間的轉化(Ikeda et al., 1991; Salim et al., 2013; De Luca et al., 2014; Miettinen et al., 2014; Dugé De Bernonville et al., 2015; Liu et al., 2015; Padhan et al., 2015; Cao et al., 2016; Rai et al., 2017)。
鑒于8HGO在單萜生物合成中的重要性,8HGO基因已從長春花(Miettinen et al., 2014)、喜樹(Valletta et al., 2010)、川西獐牙菜(Liu et al., 2017)、秦艽(Cao et al., 2016)、大花胡麻草等多種植物中分離。但是,目前8HGO基因僅在長春花中的研究最為清楚。在長春花中,Cr8HGO蛋白在8-羥香葉醇、8-羥香葉醛、8-氧香葉醇、NAD+存在的條件下,根據組合和培養時間的不同,產生以上三種復合物和8-氧香葉醛的混合物的相對比例不同,因此,該酶被命名為8-羥香葉醇氧化還原酶(圖1)(Miettinen et al., 2014)。在NAD+輔因子存在下,Cr8HGO蛋白不能轉換8-氧香葉醛,在輔因子NADP+/NADPH存在下,Cr8HGO蛋白不能催化8-羥香葉醇、8-羥香葉醛、8-氧香葉醇三個底物(Miettinen et al., 2014)。有趣的是,Cr8HGO蛋白對香葉醇、反式-2-己烯醇、法尼醇、橙花醇、異苯基芐醇、辛醇等初級醇也具有相對較高的活性(Miettinen et al., 2014)。由于存在4種相互轉化的化合物和8種可能的反應,反應動力學相當復雜,因此反應常數無法確定(Miettinen et al., 2014)。在基因表達方面,長春花Cr8HGO基因的表達受非生物因素的影響。在長春花懸浮培養細胞中,Cr8HGO基因能夠被茉莉酸甲酯(MeJA)強烈誘導表達;而在幼苗中,Cr8HGO基因在MeJA處理6 h后被輕微誘導表達;在懸浮細胞中過表達ORCA2(硬脂酸應答的長春花花分生組織決定基因2/乙烯響應因子)并不能誘導Cr8HGO的表達,過表達ORCA3反而抑制Cr8HGO基因的表達(Miettinen et al., 2014)。在UV-B脅迫15 min和60 min下,長春花幼苗葉片中Cr10HGO基因表達上調0.75倍;在UV-B脅迫后,黑暗培養72 h,Cr10HGO蛋白含量是對照組的1.98倍,其下游產物異胡豆苷增加約5倍(Zhu et al., 2015)。
為了探究滇龍膽Gr8HGO基因是否參與龍膽苦苷生物合成、其他單萜的生物合成等代謝過程,為龍膽苦苷和單萜類生物合成提供優質基因資源,本研究根據前期本課題組的滇龍膽葉和根的轉錄組數據庫,設計引物,對滇龍膽Gr8HGO基因進行克隆,并進行序列分析、原核表達載體構建和組織器官特異性表達分析,為今后闡明Gr8HGO基因在龍膽苦苷合成中的作用奠定基礎。
1材料與方法
1.1 材料
滇龍膽(Gentiana rigescens)植株采自于臨滄耀陽生物醫藥有限公司后箐基地,種植于玉溪師范學院后勤基地。使用滇龍膽幼葉進行RNA提取,使用土壤栽培兩年生滇龍膽的根、莖和葉進行基因表達分析,取樣日期為2018年4月30日。
1.2 方法
1.2.1 葉片總RNA提取、反轉錄及Gr8HGO基因ORF的克隆使用植物RNA提取試劑盒MiniBEST Plant RNA Extraction Kit(Takara,大連)進行滇龍膽幼葉總RNA的提取;使用逆轉錄試劑盒PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit(Takara,大連)進行cDNA的合成。參照pET32a原核表達載體的多克隆位點和滇龍膽轉錄組數據庫中Gr8HGO基因序列,設計一對特異性引物Gr8HGOBamHI-F:GGATCCATGACTAAAACCATTA
CTCCTGC(下劃線為BamHI酶切位點),Gr8HGOXhoI-R:CTCGAGTTAAAACTTAATTACAA
CCTTAACAC(下劃線為XhoI酶切位點)。以上述cDNA為模板進行PCR擴增,擴增產物經回收后克隆到pMD19-T載體(Takara,大連)。轉化大腸桿菌DH5α感受態細胞(Takara,大連)后。挑取菌落后,提質粒,經雙酶切檢測正確后送生工生物工程上海股份有限公司進行DNA測序,獲得重組質粒pMD19-Gr8HGO。
1.2.2 原核表達載體構建使用雙酶切、連接的方法構建原核表達載體pET32a-Gr8HGO(張曉東等, 2016)。
1.2.3 Gr8HGO基因及其編碼蛋白的序列分析使用NCBI-BLASTp軟件進行蛋白同源性比對,使用離線軟件DNAMAN 7進行多序列比對;使用MEGA X離線軟件的Clustal W進行多序列比對,然后使用NJ法創建系統發育樹,Bootstrap=1 000;利用網絡軟件對基因稀有密碼子進行分析(張曉東等, 2016)。使用ChloroP v1.1預測葉綠體轉運肽;使用在線InterPro軟件預測蛋白保守結構域;使用在線軟件ProtScale分析蛋白的親疏水性;使用在線軟件SPOMA預測蛋白的二級結構;使用在線軟件Phyre2預測蛋白的三級結構;使用SWISS-MODEL進行同源建模分析;利用Expasy中的TMHMM服務器V2.0預測蛋白的跨膜螺旋區;利用Plant-mPLoc、WOLF PSORT和ProtComp 9.0三個軟件對蛋白的亞細胞定位情況進行預測。
1.2.4 Gr8HGO基因的表達分析分別提取兩年生滇龍膽的根、莖和葉的總RNA,使用反轉錄試劑盒PrimeScriptTM RT Master Mix (Perfect Real Time)(Takara,大連)合成第一鏈cDNA。根據Gr8HGO基因序列設計一對特異性引物qGr8HGO-F(5′-AGGACAAACAGTGTACCACC-3′)和qGr8HGO-R(5′-CAACCTCTCTCCAAGCTGCC-3′),用于所有Gr8HGO基因的表達分析。以滇龍膽GrACTIN基因為內參,使用嵌合熒光檢測試劑盒TB GreenTM Premix Ex TaqTM Ⅱ(Takara,大連)進行qPCR。每個反應重復3次。反應在LightCycler 480 Ⅱ熒光定量PCR儀(Roche,瑞士)上進行擴增,擴增結果使用內參基因校準后,采用比較Ct值的“2-△△Ct”的方法自動計算出根、莖、葉中Gr8HGO基因相對表達量。
2結果與分析
2.1 滇龍膽Gr8HGO基因序列的克隆
以逆轉錄獲得的cDNA為模板,使用基因特異黑色表示相似性等于100%;粉紅色表示75%≤相似性<100%;淺藍色表示50%≤相似性<75%。
性引物共克隆到5個Gr8HGO基因,分別為Gr8HGO-1(KP722029.1)、Gr8HGO-2(KP722030.1)、Gr8HGO-3(KP722031.1)、Gr8HGO-4(KP722032.1)、Gr8HGO-5(KP722052.1),其中Gr8HGO-1基因ORF長1 062 bp,編碼353個氨基酸;其余4條基因ORF長1 131 bp,編碼376個氨基酸。
2.2 Gr8HGO基因的生物信息學分析
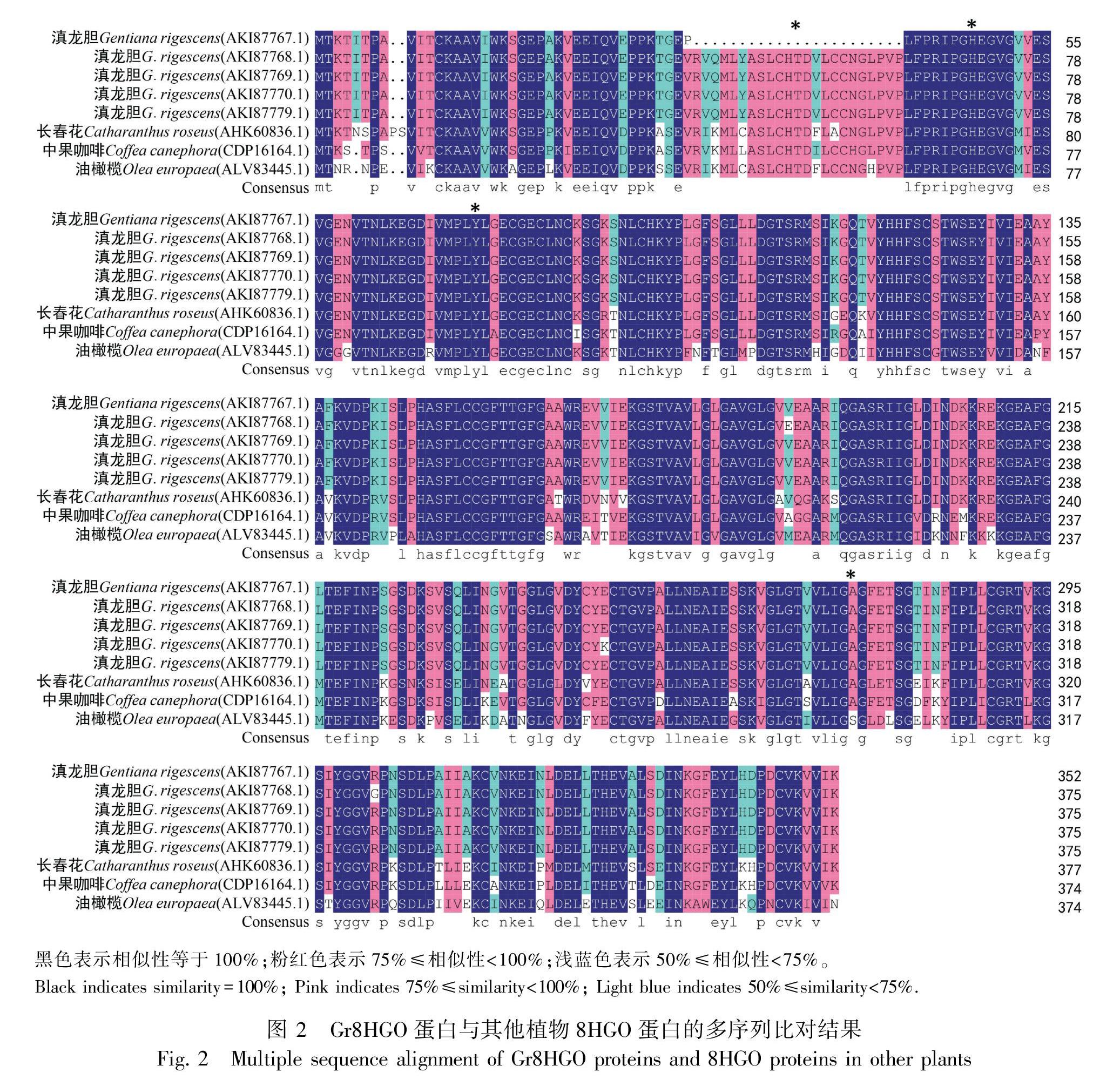
利用GenBank數據庫中的BLASTp程序和DNAMAN 7對Gr8HGO蛋白的氨基酸序列進行相似性分析,結果表明滇龍膽Gr8HGO-1、Gr8HGO-2、Gr8HGO-3、Gr8HGO-4、Gr8HGO-5蛋白與長春花Cr8HGO蛋白序列相似性均最高,分別為84.31%、84.31%、84.84%、84.57%、84.84%,其次是中果咖啡81.25%、81.07%、81.33%、81.07%、81.33%,再次是油橄欖73.86%、73.87%、74.13%、73.87%、74.13%。多序列比對分析結果表明5個Gr8HGO蛋白與已知8HGO蛋白序列相似性很高(圖2),總體相似度高達91.27%。利用MEGA X將Gr8HGO氨基酸序列與BLASTp比對中相似性較高的蛋白氨基酸序列進行系統發育分析,結果顯示5個滇龍膽Gr8HGO蛋白與長春花Cr8HGO蛋白處于同一進化枝(圖3),表明它們的親緣關系較近。
使用Expasy ProtParam工具對5個Gr8HGO蛋白和Cr8HGO蛋白的理化性質進行分析,結果表明在N端部分Gr8HGO-1蛋白比其他4個Gr8HGO蛋白少23個氨基酸,后4個Gr8HGO蛋白與Cr8HGO蛋白大小相似(表1)。與Cr8HGO相比,5個Gr8HGO蛋白帶正負電荷的氨基酸殘基數目相差較大,導致其理論pI更偏酸性(表1)。5個Gr8HGO蛋白與Cr8HGO都為疏水穩定蛋白,但是前者的疏水性更強(表1)。使用ProtScale軟件對5個Gr8HGO蛋白與Cr8HGO蛋白進行疏水性分析,結果與表1一致。5個Gr8HGO蛋白與Cr8HGO蛋白在大腸桿菌、酵母和哺乳動物細胞體內的半衰期都分別大于10、 20和30 h。對5個Gr8HGO基因進行稀有密碼子分析,結果顯示這些基因中稀有密碼子含量均低于0.85%,且無稀有密碼子連續出現的情況,因此可使用大腸桿菌BL21或Rosetta(DE3)進行誘導表達。
利用SPOMA服務器對5個Gr8HGO蛋白與Cr8HGO蛋白的二級結構進行分析,結果表明Gr8HGO-1的α-螺旋(H)含量最高,延伸鏈(E)含量最低,5個Gr8HGO蛋白的β-轉角(T)含量高于Cr8HGO,Gr8HGO-2中無規則卷曲含量最低(表2)。
利用SignalP 4.1服務器對5個Gr8HGO蛋白與Cr8HGO蛋白的信號肽序列進行分析,結果均沒有發現信號肽,表明這些蛋白都是非分泌型蛋白(表2)。使用TMHMM2.0軟件預測5個Gr8HGO蛋白與Cr8HGO蛋白的跨膜螺旋區,結果表明這些蛋白都不包含跨膜螺旋區域,為非膜蛋白(表2)。使用ChloroP v1.1 Server對5個Gr8HGO蛋白與Cr8HGO蛋白的葉綠體轉運肽進行預測,結果表明這些蛋白不包含葉綠體轉運肽(表2)。
使用Plant-mPLoc軟件和ProtComp 9.0軟件進行亞細胞定位分析,結果是5個Gr8HGO蛋白和Cr8HGO蛋白均定位于細胞質 (表3)。而使用WOLF PSORT Prediction軟件進行亞細胞定位分析的結果是這些蛋白最可能為分泌蛋白(表3)。
利用SWISS-MODEL Workspace的自動模式預測5個Gr8HGO蛋白和Cr8HGO蛋白的三級結構,結果如表4,表明滇龍膽5個Gr8HGO蛋白屬于Ⅱ類乙醇脫氫酶,而長春花Cr8HGO屬于Ⅲ類乙醇脫氫酶(圖4)。與其他蛋白相比,Gr8HGO-1蛋白在N末端缺失23個氨基酸,導致其在三維結構模型上表現為蛋白左上側的β片層結構減少(圖4)。
使用InterPro71.0在線軟件對5個Gr8HGO蛋白和Cr8HGO蛋白進行保守結構域分析,結果表明Gr8HGO-1蛋白僅包含乙醇脫氫酶N端結構域(IPR013154)和C端結構域(IPR013149),而后4個Gr8HGO蛋白和Cr8HGO蛋白除具有以上兩個保守結構域外,還具有聚酮合酶、烯酰還原酶結構域(IPR020843)(表5)。在催化Zn結合位點方面,Gr8HGO-1僅具有2個結合位點,而后4個Gr8HGO蛋白和Cr8HGO蛋白具有4個結合位點;在結構Zn結合位點方面,所有蛋白都具有4個結合位點;在NAD結合位點方面,Gr8HGO-1僅具有20個結合位點,而后4個Gr8HGO蛋白和Cr8HGO蛋白則具有22個結合位點;在底物結合位點方面,Gr8HGO-1僅具有3個結合位點,比后4個Gr8HGO蛋白和Cr8HGO蛋白少一個結合位點;在二聚體界面方面,所有蛋白都具有29個界面接觸位點(表5)。GO注釋結果表明所有蛋白均參與氧化還原過程(GO:0055114),分子功能為Zn2+結合(GO:0008270)和氧化還原酶活性(GO:0016491),未預測到其在細胞組分中的作用。
2.3 Gr8HGO-2基因原核表達載體構建
使用限制性內切酶BamHI和XhoI對質粒pET32a-Gr8HGO-2進行雙酶切檢測,結果能夠切出目的片段和載體(圖5), 表明Gr8HGO-2基因已成功插入原核表達載體pET32a中。
2.4 Gr8HGO基因的組織表達分析
根據5個Gr8HGO基因序列,設計一對檢測5個基因表達情況的引物, 通過qRT-PCR技術對Gr8HGO基因在兩年生滇龍膽的根、莖和葉等組織器官中的表達情況進行分析,結果顯示Gr8HGO基因在葉中表達量最高,分別是根和莖中的31.46倍和26.70倍,根和莖中僅有少量表達(圖6)。
3討論
龍膽苦苷是滇龍膽、龍膽、三花龍膽和條葉龍膽、秦艽、川西獐牙菜等中藥材的主要藥效成分(Hua et al., 2014; 國家藥典委員會, 2015; Liu et al., 2017),因此克隆滇龍膽龍膽苦苷生物合成途徑中的基因并對其進行組織特異性表達分析,對于揭示這一類藥材主要藥效成分的生物合成都具有重要意義。在滇龍膽生產中,主要存在龍膽苦苷含量低(Pan et al., 2015)和連作障礙(來源于臨滄耀陽生物藥業科技公司)兩個問題,可通過合成生物學手段工廠化生產龍膽苦苷和培育高龍膽苦苷含量的新品種來解決,但前提是需要首先闡明龍膽苦苷的生物合成途徑。8-羥香葉醇氧化還原酶是龍膽苦苷和其他單萜類生物合成途徑中的重要的催化酶(Cao et al., 2016),因此8HGO基因的表達情況能夠直接影響這些藥材龍膽苦苷的生物合成。本研究基于滇龍膽轉錄組數據庫,在滇龍膽中克隆到5個8HGO基因,表明在滇龍膽中8HGO基因可能存在功能冗余現象。
高效的途徑工程需要不同途徑組分的細胞和亞細胞組織的精確知識(Miettinen et al., 2014)。本研究使用Plant-mPLoc和ProtComp 9.0軟件對5個Gr8HGO蛋白和Cr8HGO蛋白進行亞細胞定位預測,結果為所有蛋白均定位于細胞質,這與通過Cr8HGO與GFP融合表達試驗獲得的長春花Cr8HGO蛋白定位結果一致(Miettinen et al., 2014)。而WOLF PSORT軟件預測的結果卻是Gr8HGO蛋白最可能為分泌蛋白,這是由于不同軟件的預測方法不同所致。ProtComp 9.0軟件綜合了蛋白質定位預測的許多方法,如基于神經網絡的預測方法,與已知定位同源蛋白的直接比較的方法,預測某些功能肽序列如線粒體和葉綠體信號肽、轉運肽和跨膜區段的方法等。上海交通大學沈紅斌教授開發的Plant-mPLoc軟件,是基于已知定位的1 055個蛋白質序列數據庫的預測方法,該軟件解決了單個蛋白存在多種亞細胞定位的問題(Chou & Shen, 2010)。而WOLF軟件是由日本人較早開發的基于分類信號模序和相關序列特征如氨基酸組成等進行預測的軟件。因此,進行蛋白亞細胞定位預測時,需要使用多種軟件進行綜合分析。
在本研究中,三維模型預測結果表明,5個Gr8HGO蛋白與Cr8HGO蛋白的活性形式為二聚體,這與5個Gr8HGO蛋白與Cr8HGO蛋白都具有29個二聚體界面接觸位點相一致,與在印度蘿芙木中,Rs10HGO蛋白活性形式是單體的結果不同(Ikeda et al., 1991)。因此,需要通過實驗來進一步驗證。
組織器官特異性表達分析結果顯示,本研究中的Gr8HGO基因主要在葉中表達,而在根和莖中幾乎不表達。這與滇龍膽的主要藥效成分龍膽苦苷主要在葉中合成、根中累積的結果相一致(楊美權等, 2011; 朱宏濤等, 2011),這表明所克隆的5個Gr8HGO基因在葉中而不是根中促進龍膽苦苷的生物合成。由于5個Gr8HGO基因的高度相似性,很難設計特異性引物將它們的組織特異性表達區分開,因此目前無法判斷究竟是哪一個或哪幾個Gr8HGO基因在葉中高表達,今后可采用高通量測序技術加以區分。我們的結果與在喜樹中Ca8HGO主要在莖和葉中表達,在喜樹堿累積的根中不表達的結果相一致(Valletta et al., 2010)。而與在兩年生花期的麻花艽Gs10HGO基因在花中表達量最高,其次是根,表達量最低的是葉的結果剛好相反(Zhou et al., 2016),與在溫室栽培的印度獐牙菜植株中Sc8HGO基因在葉和根中均表達(Padhan et al., 2015)、在川西獐牙菜中Sm8HGO基因在根、莖、葉和花中的表達量差異不大(Liu et al., 2017)的結果也不同。以上結果表明滇龍膽和喜樹具有相似的活性成分合成與累積模式,而與麻花艽和印度獐牙菜不同。下一步將對Gr8HGO蛋白進行誘導表達、蛋白純化和酶活性分析,為該基因功能的闡明奠定基礎。
參考文獻:
CAO XY, GUO XR, YANG XB, et al., 2016. Transcriptional responses and gentiopicroside biosynthesis in methyl jasmonate-treated Gentiana macrophylla seedlings [J]. PLoS ONE, 11(11): e0166493.
CHOU KC, SHEN HB, 2010. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization [J]. PLoS ONE, 5(6): e11335.
DE LUCA V, SALIM V, THAMM A, et al., 2014. Making iridoids/secoiridoids and monoterpenoid indole alkaloids: progress on pathway elucidation [J]. Curr Opin Plant Biol, 19: 35-42.
DUG DE BERNONVILLE T, FOUREAU E, PARAGE C, et al., 2015. Characterization of a second secologanin synthase isoform producing both secologanin and secoxyloganin allows enhanced de novo assembly of a Catharanthus roseus transcriptome [J]. BMC Genomics, 16(1): 619.
Editorial Board of Flora of China, 1988. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 62: 100.? [中國植物志編委會, 1988. 中國植物志 [M]. 北京: 科學出版社, 62: 100. ]
HUA WP, ZHENG P, HE YH, et al., 2014. An insight into the genes involved in secoiridoid biosynthesis in Gentiana macrophylla by RNA-seq [J]. Mol Biol Rep, 41(7): 4817-4825.
HUANG LQ, LIU CX, 2015. Molecular pharmacognosy [M]. 3rd ed. Beijing: Science Press: 239-243.? [黃璐琦, 劉昌孝, 2015. 分子生藥學 [M]. 3版. 北京: 科學出版社: 239-243. ]
IKEDA H, ESAKI N, NAKAI S, et al., 1991. Acyclic monoterpene primary alcohol: NADP+ oxidoreductase of Rauwolfia serpentina cells: the key enzyme in biosynthesis of monoterpene alcohols [J]. J Biochem, 109(2): 341-347.
JIN H, ZHANG J, ZHANG JY, et al., 2013. Gentiana rigescens [M]. Kunming: Yunnan Science and Technology Press: 1-5.? [金航, 張霽, 張金渝, 等, 2013. 滇龍膽 [M]. 昆明: 云南科技出版社: 1-5. ]
LIU Y, SONG LL, YU WW, et al., 2015. Light quality modifies camptothecin production and gene expression of biosynthesis in Camptotheca acuminata Decne seedlings [J]. Ind Crop Prod, 66: 137-143.
LIU Y, WANG Y, GUO FX, et al., 2017. Deep sequencing and transcriptome analyses to identify genes involved in secoiridoid biosynthesis in the Tibetan medicinal plant Swertia mussotii [J]. Sci Rep, 7: 43108.
MIETTINEN K, DONG L, NAVROT N, et al., 2014. The seco-iridoid pathway from Catharanthus roseus [J]. Nat Comm, 5: 3606-3616.
MUNKERT J, POLLIER J, MIETTINEN K, et al., 2015. Iridoid synthase sctivity is common among the plant progesterone 5β-reductase family [J]. Mol Plant, 8(1): 136-152.
National Pharmacopoeia Committee, 2015. Chinese Pharmacopoeia [M]. Part I. Beijing: China Pharmacopoeia Science and Technology Press: 96.? [國家藥典委員會, 2015. 中國藥典 [M]. 一部. 北京: 中國醫藥科技出版社: 96. ]
PADHAN JK, KUMAR V, SOOD H, et al., 2015. Contents of therapeutic metabolites in Swertia chirayita correlate with the expression profiles of multiple genes in corresponding biosynthesis pathways [J]. Phytochemistry, 116(1): 38-47.
PAN Y, ZHANG J, ZHAO YL, et al., 2015. Investigation of metabolites accumulation in medical plant Gentiana rigescens during different growing stage using LC-MS/MS and FT-IR [J]. Bot Stud, 56(1): 14.
RAI A, KAMOCHI H, SUZUKI H, et al., 2017. De novo transcriptome assembly and characterization of nine tissues of Lonicera japonica to identify potential candidate genes involved in chlorogenic acid, luteolosides, and secoiridoid biosynthesis pathways [J]. J Nat Med, 71(1): 1-15.
SALIM V, YU F, ALTAREJOS J, et al., 2013. Virus-induced gene silencing identifies Catharanthus roseus 7-deoxyloganic acid-7-hydroxylase, a step in iridoid and monoterpene indole alkaloid biosynthesis [J]. Plant J, 76(5): 754-765.
VALLETTA A, TRAINOTTI L, SANTAMARIA AR, et al., 2010. Cell-specific expression of tryptophan decarboxylase and 10-hydroxygeraniol oxidoreductase, key genes involved in camptothecin biosynthesis in Camptotheca acuminata Decne (Nyssaceae) [J]. BMC Plant Biol, 10(1): 69.
YANG MQ, ZHANG JY, SHEN T, et al., 2011. Effects of different cultivation modes on gentiopicroside content in Gentiana rigescens [J]. J Jiangsu Agric Sci, (1): 287-289.? [楊美權, 張金渝, 沈濤, 等, 2011. 不同栽培模式對滇龍膽中龍膽苦苷含量的影響 [J]. 江蘇農業科學, (1): 287-289. ]
ZHANG XD, ALLAN AC, LI CX, et al., 2015. De novo assembly and characterization of the transcriptome of the Chinese medicinal herb, Gentiana rigescens [J]. Int J Mol Sci, 16(5): 11550-11573.
ZHANG XD, LI CX, WANG YZ, 2016. Cloning and expression analysis of GrCMS gene in Gentiana rigescens [J]. Bull Bot Res, 36 (2): 258-265.? [張曉東, 李彩霞, 王元忠, 2016. 滇龍膽GrCMS基因的克隆與表達分析 [J]. 植物研究, 36(2): 258-265. ]
ZHOU DW, GAO S, WANG H, et al., 2016. De novo sequencing transcriptome of endemic Gentiana straminea (Gen-tianaceae) to identify genes involved in the biosynthesis of active ingredients [J]. Gene, 575(1): 160-170.
ZHU HT, ZHENG CW, ZHAO P, et al., 2011. Content analysis of gentiopicroside in wild and tissue culture seedlings of Gentiana rigescens [J]. Nat Prod Res Dev, 23 (3): 482-485.? [朱宏濤, 鄭傳偉, 趙平, 等, 2011. 野生堅龍膽及其組培苗中龍膽苦苷的含量分析 [J]. 天然產物研究與開發, 23(3): 482-485. ]
ZHU W, YANG BX, KOMATSU S, et al., 2015. Binary stress induces an increase in indole alkaloid biosynthesis in Catharanthus roseus [J]. Front Plant Sci, 6: 582.
(責任編輯 周翠鳴)
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
