EGFL8基因沉默對肝癌細胞侵襲轉移能力的影響
2020-04-20 13:07:00
實用醫學雜志 2020年4期
暨南大學醫學院附屬廣州紅十字會醫院肝膽外科(廣州510220)
肝細胞癌(hepatocellular carcinoma,HCC)是臨床上第6位最常見的惡性腫瘤,盡管近年來針對肝癌的手術切除、經皮射頻消融術及肝移植等治療手段和分子靶向藥物的應用取得了一定的進步[1],使肝癌患者的5年生存率較此前提高了10%以上[2],但是肝癌的復發和轉移仍是肝癌患者致死的主要原因[3]。因此,探索肝癌轉移的分子機制,對于研發新的肝癌治療措施及提高其總體治療效果均具有重大意義[4]。筆者前期研究已發現EGFL8 在結直腸癌及胃癌中表達下調,且其低表達與這兩種惡性腫瘤的轉移、TMN 分期及預后密切相關[5-6],提示其可能參與多種消化道惡性腫瘤的調控,但目前尚未見EGFL8 在肝癌中的研究報道。為此,本研究檢測了EGFL8 在3種不同轉移潛能的肝癌細胞系的表達水平,并采用慢病毒介導的siRNA技術靶向沉默Hep3B 肝癌細胞系中EGFL8的表達,觀察EGFL8基因沉默對肝癌細胞侵襲遷移能力的影響。
1 材料與方法
1.1 細胞系及質粒Hep3B及SMMC-7721 肝癌細胞系均購自中國科學院上海生命科學研究院生物化學與細胞生物學研究所,HCCLM3 肝癌細胞系、293T 細胞及慢病毒質粒GV248,包裝質粒pGCLV、pHelper1.0和pHelper2.0 均購自上海吉凱基因化學技術有限公司。
1.2 實時定量PCR檢測提取細胞總RNA并逆轉錄,在熒光定量PCR 儀(TP800,日本TAKARA 公司)上行PCR 檢測,反應條件為:95℃預變性15 s,然后95℃變性5 s,60℃退火延伸30 s,共進行45個循環。EGFL8 引物為:上游,5′-GTCCCGCTCC ACTACAACG-3′;下游,5′-GCGGTACATGGTCCTGTAAGT-3′。內參GAPDH 引物為:上游,5′-TGACTTCAACAGCGACACCCA-3′;下游,5′- CACCCTGTTGCTGTA- GCCAAA-3′。以2-ΔCT法分析EGFL8基因的表達水平。
1.3 慢病毒介導的siRNA設計針對EGFL8基因的shRNA 序列如下:正義鏈:5′-CCGGCAACCAGTGCCAGCATACTCACTCGAGTGAGTATGCTGGCACTGGTTGTTTTTG-3′;反義鏈:5′-AATTCAAAAAGAGTTGGTACTGGATCACATTCTCGAGAATGTGATCCAGTACCAACTC-3′。陰性對照序列為:正義鏈:5′-CCGGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTG-3′;反義鏈:5′-AATTCAAAAATTCTCCGAACGTGTCACGTAAGTTCTCTACGTGACACGTTCGGAAA-3′。上述序列由上海吉凱基因化學技術有限公司合成,并將其所形成的DNA 雙鏈連接到siRNA慢病毒質粒GV248,再將上述質粒分別與pGC-LV、pHelper1.0和pHelper2.0 3個包裝質粒共轉染至293T 細胞,制備EGFL8-siRNA及陰性對照慢病毒。上述兩種慢病毒各自感染的Hep3B 肝癌細胞系即被分別命名為shEGFL8組(實驗組)和shCtrl組(對照組)Hep3B 細胞。采用熒光倒置顯微鏡(micropublisher 3.3RTV,日本奧林帕斯公司)觀察兩組細胞的慢病毒感染效率,并采用實時定量PCR 法檢測兩組細胞中EGFL8基因的表達水平以檢測EGFL8基因的沉默效率。
1.4 Transwell小室侵襲實驗檢測細胞侵襲能力將Transwell 小室(購自美國Corning 公司)放置于一個潔凈的24 孔板中,取100 μL 無血清培養基加入Transwell 小室,放置培養箱中1~2 h。在24 孔板下室中加入30% FBS 培養基600 μL,用無血清培養基稀釋細胞,在每個小室中加入1 × 105個細胞,把小室轉移入含30%FBS 培養基的下室中,在組織培養箱中常規培養4~24 h。移除培養基,用棉簽仔細除去非轉移細胞,在24 板的空孔中加入400 μL Giemsa 染色液,把小室浸泡在染色液中20 min,在聚碳酸酯膜的下方對轉移細胞進行染色,完成后棄去孔中培養液,沖洗數次,將小室適當風干。顯微鏡拍照計數穿過膜的細胞數即為侵襲細胞數。
1.5 劃痕實驗檢測細胞遷移能力將所有能滅菌的器械及劃痕儀在操作前使用紫外照射30 min(超凈臺內)。在96 孔板中按約3×104個細胞/孔,加入各組Hep3B 細胞系。待細胞生長鋪滿96 孔板底表面時,使用劃痕儀對準96 孔板的下端中央部位,向上平移形成劃痕。使用無血清培養基輕輕漂洗2 遍,加入低濃度血清培養基,隨后將96 孔板放入37℃、5%CO2培養箱培養。分別于劃痕后0、24和72 h 顯微鏡拍攝照片并測量劃痕寬度,并計算各組細胞24 h和72 h的細胞遷移率,計算公式為細胞遷移率=(劃痕后0 h 劃痕寬度-劃痕后24 h或72 h 劃痕寬度)/劃痕后0 h 劃痕寬度×100%。
1.6 統計學方法應用SPSS 19.0 統計軟件進行統計學分析。計量資料以均數±標準差表示,對EGFL8 在肝癌細胞系中的表達水平采用單因素方差分析檢驗,多重比較采用Tukey 檢驗;其余采用Student′st檢驗。所有分析均為雙側檢驗,P<0.05為差異具有統計學意義。統計圖采用Graphpad Prism 5.0 軟件(美國Graphpad 軟件公司)繪制。
2 結果
2.1 EGFL8在肝癌細胞系中的表達水平多重比較的方差分析顯示EGFL8 在侵襲轉移能力最強的HCCLM3細胞系中的表達水平低于侵襲轉移能力中等的SMMC-7721 細胞系(0.000 261 3 ± 0.000 019vs.0.000 602 8±0.000 031 28,P=0.007 1),后者又顯著低于侵襲轉移能力最弱的Hep3B 細胞系(0.000 602 8±0.000 031 28vs.0.002 106±0.000 782,P<0.000 1)見圖1。
2.2 慢病毒的感染效率及EGFL8基因沉默效率熒光顯微鏡觀察shCtrl組和shEGFL8組Hep3B細胞的結果顯示,兩組細胞中可發出綠色熒光的細胞比例均超過80%,顯示慢病毒的感染效率達到80%以上(圖2A)。實時定量PCR 結果顯示shEGFL8組Hep3B 細胞中的EGFL8基因表達水平僅為shCtrl組的23.3%(0.233 ± 0.007vs.1.013 ± 0.118,t= 6.582,P= 0.002 8),即EGFL8基因在Hep3B 細胞系中的沉默效率達到了76.7%(圖2B)。
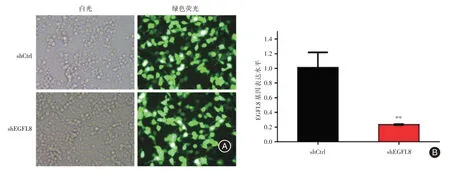
圖2 慢病毒的感染效率及沉默效率Fig.2 Infection efficiency and silencing efficiency of lentivirus
2.3 EGFL8基因沉默對肝癌細胞侵襲能力的影響Transwell 小室侵襲實驗的結果示,在同樣時間內,shEGFL8組Hep3B 細胞的侵襲細胞數量明顯高于shCtrl組(105.3±6.983vs.52.33±4.256,t=6.48,P= 0.002 9),證明shEGFL8組Hep3B 細胞的侵襲能力顯著高于shCtrl組,提示EGFL8基因沉默可增強肝癌細胞的侵襲能力(圖3)。
2.4 EGFL8基因沉默對肝癌細胞遷移能力的影響劃痕愈合實驗的結果顯示,shEGFL8組Hep3B細胞的24及72 h 遷移均高于shCtrl組(24 h 遷移率:10.67% ± 1.20%vs.5.67% ± 0.33%,t= 4.009,P= 0.016;72 h 遷移率:22.67% ± 1.20%vs.12.67%± 1.20%,t= 5.883,P= 0.004),提示EGFL8 沉默可明顯增強肝癌細胞的遷移能力(圖4)。
3 討論
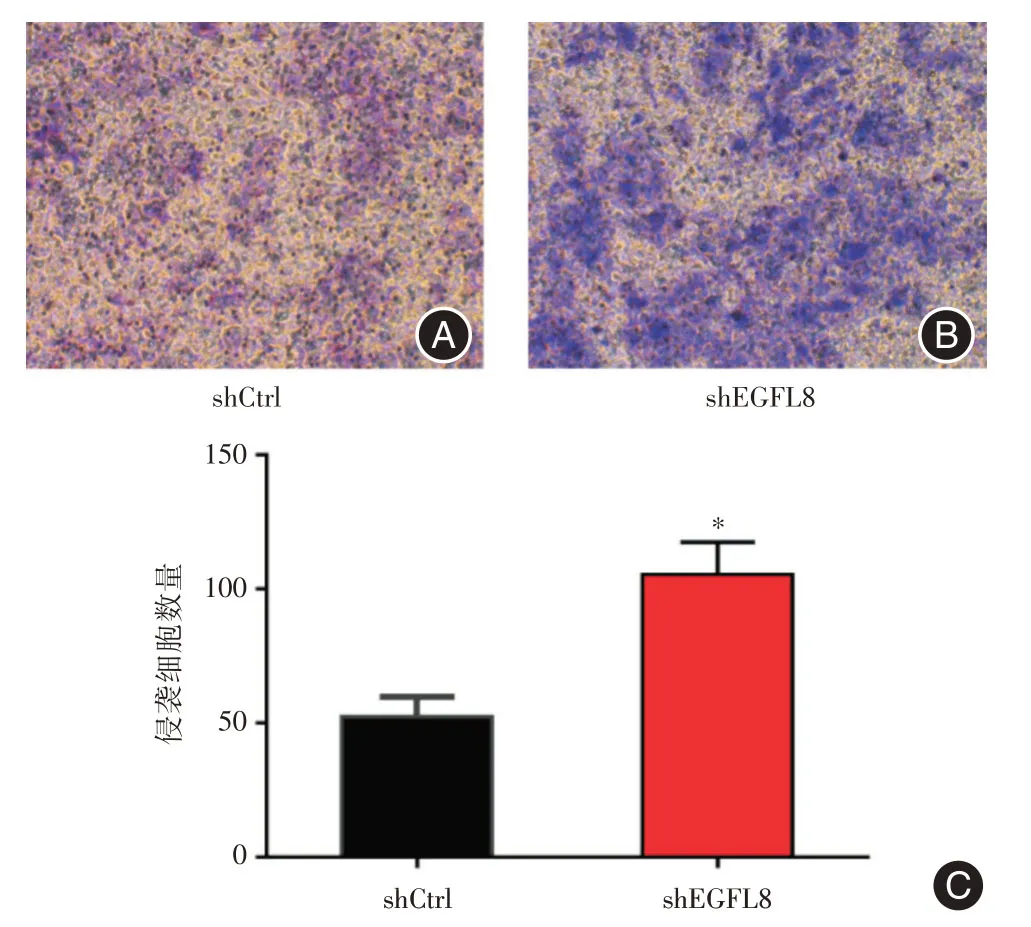
圖3 Transwell 小室侵襲實驗結果Fig.3 Results of transwell chamber invasion assay
原發性肝癌是全球第5 大常見的惡性腫瘤,占腫瘤致死原因的第3位,全球每年有超過50萬人罹患肝癌,其中一半以上在中國[7]。盡管針對肝癌的治療手段眾多,如手術切除、介入治療、傳統放化療、基因靶向治療、生物治療和中醫藥治療等,但根治性肝切除術仍然是治療肝癌的最主要的手段,然而,即便在實施了肝癌肝切除術的患者中,仍然有超過50%以上的患者出現術后的復發和轉移[1],這顯著限制了肝癌總體治療效果的進一步提高。因此,肝癌侵襲轉移的機制已成為目前肝癌研究中的重點問題。
在前期研究中,筆者闡述了EGFL7 基因可通過EGFL7/EGFR/FAK 這一信號通路影響肝癌的侵襲轉移,顯示EGFL7可成為一個新的潛在肝癌治療靶點[8]。EGFL8基因是EGFL7的同源基因,兩者均為EGFL 基因家族成員[9]。EGFL8 蛋白與EGFL7蛋白具有極為相似的分子結構,兩者均屬于分泌型蛋白,都含有一個EGF 樣結構域,一個鈣離子結合型EGF 樣結構域和N 尾端的信號肽[9-10],且二者在成年小鼠組織中的表達水平也很相似,均在腎、腦、胸腺及肺組織中高表達[10],這種結構和表達譜的高度相似性提示EGFL8 可能和EGFL7 一樣影響人類惡性腫瘤的發生發展。而筆者前期研究也確實證實EGFL8 在人結直腸癌及胃癌組織中的表達水平較正常組織明顯下調,且其低表達與結直腸癌及胃癌的轉移、疾病進展及不良預后密切相關。然而,目前尚無EGFL8 在肝癌中表達及功能的研究報道。因此,本研究檢測了EGFL8 在Hep3B 等三種肝癌細胞系中的表達水平,結果發現轉移潛能最強的HCCLM3 肝癌細胞系[11]中的EGFL8表達水平低于中等轉移潛能的SMMC-7721肝癌細胞系[12],后者中的EGFL8表達水平又低于轉移潛能最弱的Hep3B 肝癌細胞系[13],提示EGFL8表達水平的下調可能與肝癌細胞轉移潛能的增強相關,然而EGFL8 在肝癌中發揮的功能尚不清楚。
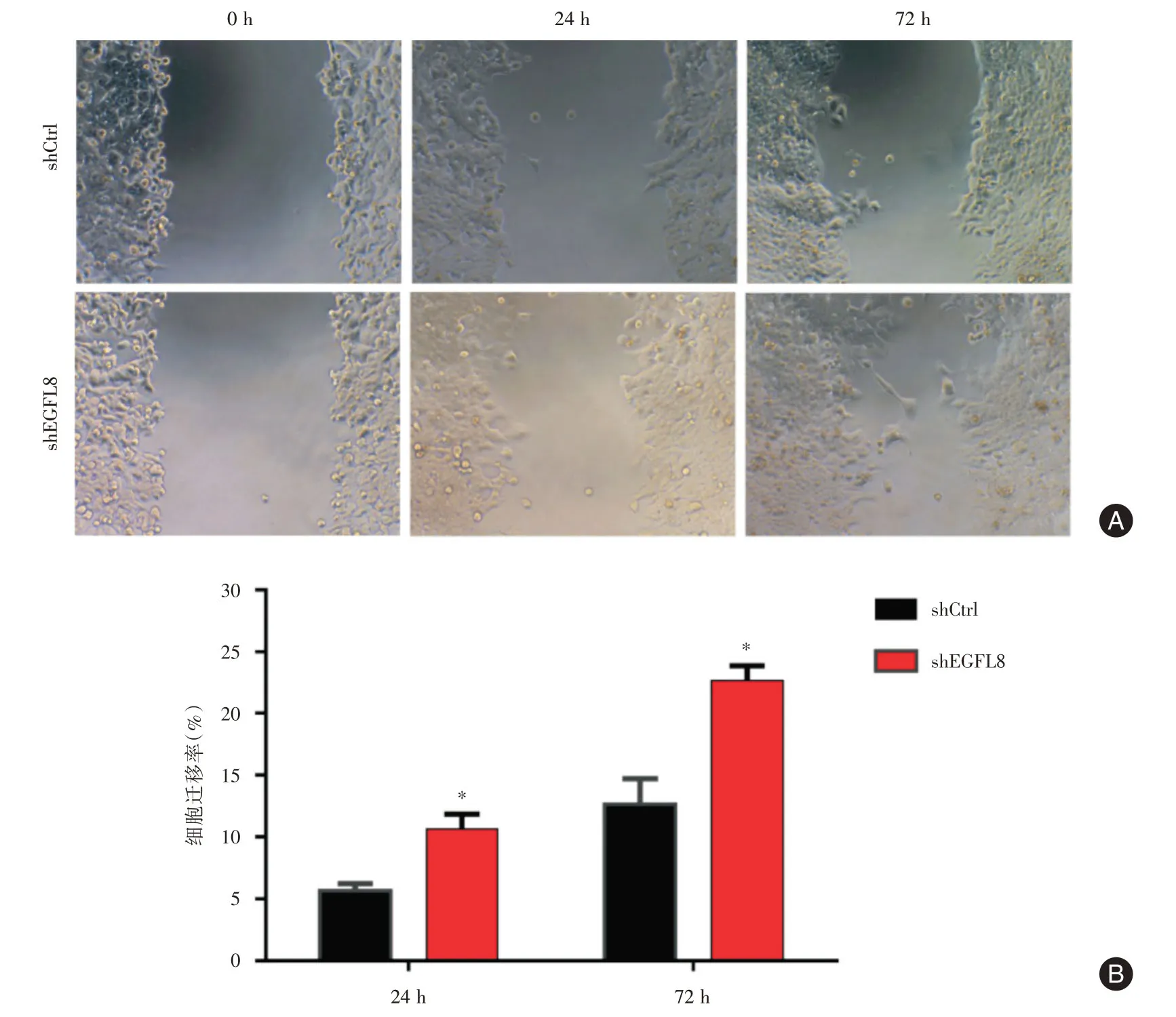
圖4 劃痕愈合實驗的結果Fig.4 Results of wound healing assay
為進一步研究EGFL8在肝癌中的生物學功能,本研究采用小干擾RNA技術靶向抑制了3種肝癌細胞系中EGFL8表達水平最高的Hep3B 肝癌細胞系中EGFL8基因的表達,從而模擬EGFL8基因在肝癌組織中表達水平下調的狀態,以此觀察EGFL8 下調對肝癌侵襲轉移能力的影響。相較于直接轉染的siRNA 片段易降解而導致基因沉默不持久,慢病毒介導的siRNA技術具有可感染分裂期和非分裂期細胞、轉移基因片段容量大、目的基因表達時間長、不易誘發宿主免疫反應等優點[14],這使其成為目前主要的基因抑制技術[15]。本研究結果顯示慢病毒的感染效率達到80%以上,而EGFL8基因的沉默效率達到了76.7%,顯示了慢病毒介導的siRNA技術在基因靶向沉默中的優勢。進而,通過Transwell 小室侵襲實驗及劃痕愈合實驗來觀察EGFL8基因表達抑制對肝癌細胞侵襲遷移能力的影響。實驗結果顯示,EGFL8 沉默組Hep3B 肝癌細胞的侵襲能力和遷移運動能力較之對照組細胞都有明顯的增強,這不僅驗證了筆者之前的推測,即EGFL8基因可能參與了肝癌的發生發展過程,而且與前期研究發現的EGFL8表達下調與結直腸癌及胃癌的腹膜播散及肝轉移等密切相關的結果相吻合[5-6],提示EGFL8基因可能發揮了抑制肝癌細胞侵襲遷移的作用。
有研究[16]顯示,EGFL8 對小鼠胸腺上皮細胞中細胞間粘附因子(ICAM-1)的表達具有負向調控的作用:即EGFL8 過表達可抑制小鼠胸腺上皮細胞中ICAM-1的表達,而EGFL8基因沉默則可促進小鼠胸腺上皮細胞中ICAM-1的表達。因為ICAM-1在現有的研究中被公認為可促進肝癌等多種惡性腫瘤的侵襲轉移[17-18],而胸腺上皮細胞和肝癌細胞都屬于腺上皮細胞,而筆者推測EGFL8 在肝癌中可能也存在對ICAM-1 表達的負向調控,即沉默EGFL8基因表達時可能上調肝癌細胞中ICAM-1的表達水平,進而增強肝癌的侵襲轉移。此外,重組小鼠EGFL8 蛋白可抑制小鼠胸腺細胞中Notch信號通路下游作用基因Hes1和Hey1的表達[19],而后者已被證實可促進肝癌的轉移[12-20],提示EGFL8也有可能通過抑制Notch 信號通路發揮其抑制肝癌侵襲轉移的功能。由于EGFL8 蛋白和EGFL7蛋白有著相同的蛋白質結構,而EGFL7 蛋白又稱VE-statin,是一種血管內皮抑制素,通過抑制平滑肌細胞的運動來抑制血管的新生和成熟[21-22]。筆者由此推測,肝癌細胞中的EGFL8表達下調也有可能通過提高平滑肌細胞的運動能力來促進腫瘤內血管的生成,從而提高肝癌細胞的增殖、侵襲及轉移能力。顯然,EGFL8 調控肝癌細胞侵襲轉移能力的具體分子機制和EGFL8 在肝癌中的更多生物學功能目前尚不清楚,還有待于進一步研究闡明。
綜上所述,本研究EGFL8的表達下調與肝癌細胞系的轉移潛能相關,EGFL8基因沉默則可顯著提高Hep3B 肝癌細胞系的侵襲遷移能力,提示EGFL8 可能是一個肝癌轉移抑制基因,有望成為降低肝癌復發轉移的治療靶點。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
人大建設(2019年12期)2019-05-21 02:55:32
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
