基于線(xiàn)粒體D-loop區(qū)序列和Cyt b基因的鱘魚(yú)群體遺傳多樣性分析
2020-04-22 06:26:00李世凱
西南農(nóng)業(yè)學(xué)報(bào) 2020年1期
孔 杰,劉 偉,李世凱,周 洲,趙 鳳
(貴州省農(nóng)業(yè)科學(xué)院 水產(chǎn)研究所,貴州 貴陽(yáng) 550025)
【研究意義】西伯利亞鱘(A.baerii)和施氏鱘(Acipenserschrenkii)屬于鱘科(Acipenseridae)鱘屬(Acipenser),淡水定居鱘種,是我國(guó)鱘魚(yú)養(yǎng)殖生產(chǎn)的主要品種。鱘魚(yú)是多倍體起源魚(yú)類(lèi),雜交能力強(qiáng),由于鱘魚(yú)性成熟時(shí)間長(zhǎng),親魚(yú)培育成本高,導(dǎo)致養(yǎng)殖場(chǎng)常忽略鱘魚(yú)繁殖中的遺傳背景問(wèn)題,種質(zhì)混亂現(xiàn)象普遍,而后備親魚(yú)的遺傳多樣性直接影響到苗種質(zhì)量,因此對(duì)后備親魚(yú)的遺傳多樣性研究十分必要。【前人研究進(jìn)展】線(xiàn)粒體DNA(Mitochondrial DNA,mtDNA)與核基因組相比,具有變異率高、進(jìn)化速度快,無(wú)組織特異性、母系遺傳的特點(diǎn),被廣泛應(yīng)用于魚(yú)類(lèi)的群體遺傳學(xué)研究,如鱘鰉魚(yú)[1-2]、鯉[3]分子鑒定,鱘魚(yú)[4-6]、銀鯧[7]、裂腹魚(yú)[8-9]等的遺傳多樣性和遺傳結(jié)構(gòu)研究。mtDNA的不同區(qū)域進(jìn)化速度不同,適合不同水平的進(jìn)化研究。其中,線(xiàn)粒體控制區(qū)(D-loop)位于tRNAPro和rRNAPhe基因之間的控制區(qū),不參與編碼蛋白,是mtDNA序列中變異最大的區(qū)域,進(jìn)化速度快,適用于親緣關(guān)系較近群體間的比較研究和種群水平差異檢測(cè)[10-11];細(xì)胞色素b(Cytb)基因是編碼蛋白基因,進(jìn)化速度適中,適合魚(yú)類(lèi)的種間和種內(nèi)遺傳分化研究[11-12]。結(jié)合D-loop和Cytb分子標(biāo)記可以更準(zhǔn)確地分析魚(yú)類(lèi)種內(nèi)遺傳變異。【本研究切入點(diǎn)】以施氏鱘和西伯利亞鱘親魚(yú)群體為研究對(duì)象,開(kāi)展mtDNA Cytb基因和D-loop的序列對(duì)比分析,探討2標(biāo)記對(duì)鱘魚(yú)遺傳多樣性分析結(jié)果的差異與關(guān)系。【擬解決的關(guān)鍵問(wèn)題】旨在擴(kuò)大研究結(jié)果之間橫向比較,使結(jié)果更客觀(guān),評(píng)估養(yǎng)殖場(chǎng)施氏鱘和西伯利亞鱘后備親魚(yú)群體的遺傳多樣性,為鱘魚(yú)的遺傳選育提供參考。
1 材料與方法
1.1 試驗(yàn)材料
于2017年11月在貴州省惠水鱘魚(yú)養(yǎng)殖廠(chǎng),分別采集西伯利亞鱘親本尾鰭樣本26個(gè),施氏鱘親本尾鰭樣本29個(gè)。尾鰭樣本固定于無(wú)水乙醇中,4 ℃保存?zhèn)溆谩?/p>
1.2 基因組提取與擴(kuò)增
剪取約50 mg鱘魚(yú)鰭條樣本,液氮研磨至粉末,采用磁珠法基因組DNA(動(dòng)物)抽提試劑盒(上海生工)進(jìn)行DNA提取,DNA質(zhì)量與濃度用瓊脂糖凝膠電泳和NanoDrop lite(Thermo Scientific)紫外分光光度計(jì)進(jìn)行測(cè)定。Cytb的擴(kuò)增引物[13],Cytb-F:5′-GACTTGAAAAACCACCGTTG-3′,Cytb-R:5′-CTCCATCTCCGGTTTACAAGAC-3′;D-loop區(qū)序列的擴(kuò)增引物[14],tpro-F1:5′-AACTCCCAAAGCTAAGATTC-3′,phe-R2:5′-ATCCTTTAGTTAAGCTACGC-3′。
PCR反應(yīng)總體積為50 μl,其中:模板DNA 100 ng,PremixTaq25 μl,上下游引物(10 μmol/L)各1 μl,補(bǔ)充無(wú)菌水至50 μl。PCR擴(kuò)增程序:95 ℃預(yù)變性4 min;95 ℃變性45 s,54 ℃/56 ℃(D-loop/Cytb)退火45 s,72 ℃延伸90 s/2 min(D-loop/Cytb),擴(kuò)增35個(gè)循環(huán);最后72 ℃延伸8 min。PCR擴(kuò)增產(chǎn)物經(jīng)1 %瓊脂糖凝膠電泳檢測(cè)后,由上海生工進(jìn)行測(cè)序。
1.3 數(shù)據(jù)處理
采用MEGA 6.0 軟件對(duì)序列進(jìn)行多重比較,分析序列堿基組成、平均轉(zhuǎn)換/顛換率和變異位點(diǎn)。采用DnaSP 5.0 軟件統(tǒng)計(jì)獲取群體序列遺傳多樣性參數(shù),如單倍型個(gè)數(shù)(h)、單倍型多樣性(Hd)、核苷酸多樣性(π)、平均核苷酸差異(K)和Tajima’s D、Fu’s Fs值等。
2 結(jié)果與分析
2.1 西伯利亞鱘與施氏鱘的序列對(duì)比
2.1.1 Cytb序列 對(duì)西伯利亞鱘、施氏鱘Cytb序列測(cè)序并經(jīng)人工序列校正,選擇西伯利亞鱘1075 bp、施氏鱘1068 bp進(jìn)行序列比對(duì)。從表1看出,西伯利亞鱘Cytb基因在1075 bp序列中,變異位點(diǎn)5個(gè),均為單突變位點(diǎn)。A、T、C、G堿基的平均含量分別為26.1 %、27.1 %、31.6 %和15.2 %,A+T含量略高于G+C含量,基于最大似然法估算的轉(zhuǎn)換/顛換比為4.0。
施氏鱘Cytb基因在1068 bp序列中,變異位點(diǎn)4個(gè),約占分析位點(diǎn)的0.4 %,其中簡(jiǎn)約信息位點(diǎn)3個(gè),單突變位點(diǎn)1個(gè)。A、T、C、G堿基的平均含量分別為26.7 %、26.9 %、31.9 %和14.6 %,A+T含量略高于G+C含量,基于最大似然法估算的轉(zhuǎn)換/顛換比為3.0。
2.1.2 D-loop序列 對(duì)西伯利亞鱘、施氏鱘D-loop全序列測(cè)序并經(jīng)人工序列較正,選擇施氏鱘780 bp,西伯利亞鱘994 bp進(jìn)行序列比對(duì)。從表1還可看出,西伯利亞鱘D-loop在994 bp序列中,變異位點(diǎn)2個(gè),約占分析位點(diǎn)的0.2 %,其中簡(jiǎn)約信息位點(diǎn)1個(gè),單突變位點(diǎn)1個(gè)。A、T、C、G堿基的平均含量分別為32.3 %、32.0 %、21.7 %和14.0 %,A+T含量明顯高于G+C含量,基于最大似然法估算的轉(zhuǎn)換/顛換比為1.0。
施氏鱘D-loop在780 bp序列中,共有變異位點(diǎn)55個(gè),約占分析位點(diǎn)的7.0 %,其中簡(jiǎn)約信息位點(diǎn)39個(gè),單突變位點(diǎn)16個(gè)。A、T、C、G堿基的平均含量分別為30.2 %、31.6 %、20.4 %和17.8 %,A+T含量明顯高于G+C含量,基于最大似然法估算的轉(zhuǎn)換/顛換比為4.2。
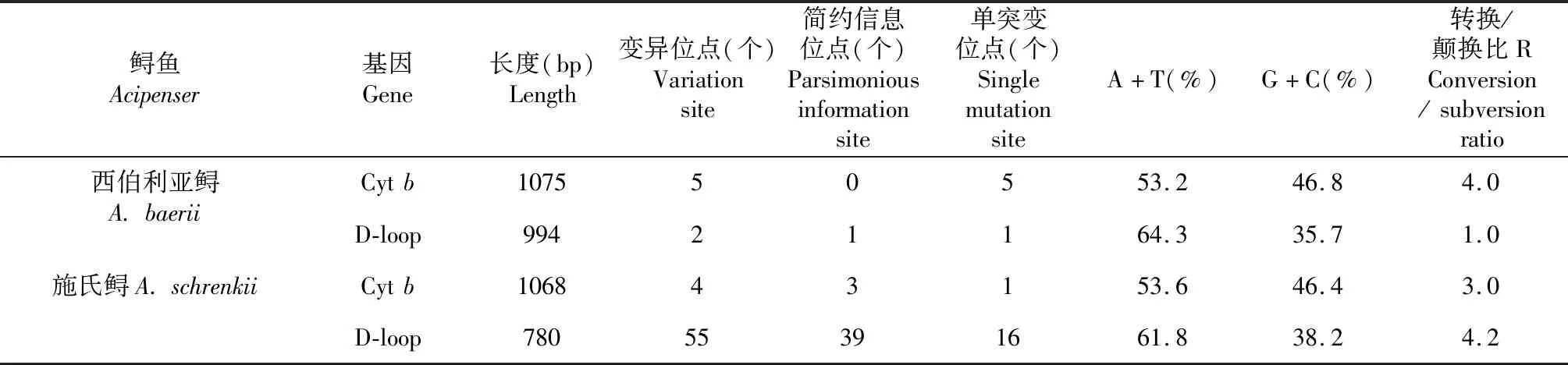
表1 西伯利亞鱘和施氏鱘的序列信息 Table 1 Base sequence information of A. baerii and A. schrenkii population
2.2 親魚(yú)群體遺傳多樣性
2.2.1 施氏鱘 從表2~3看出,基于D-loop基因,施氏鱘親魚(yú)養(yǎng)殖群體多態(tài)性位點(diǎn)數(shù)為55個(gè)、單倍型5個(gè)、單倍型多樣性0.707、核苷酸多樣性0.02242、平均核苷酸差異16.906。共定義了5個(gè)單倍型,其中Hap5為優(yōu)勢(shì)單倍型,代表14個(gè)個(gè)體,占總體個(gè)數(shù)的48.3 %。
基于Cytb基因,施氏鱘親魚(yú)養(yǎng)殖群體多態(tài)性位點(diǎn)數(shù)為4個(gè)、單倍型4個(gè)、單倍型多樣性0.612、核苷酸多樣性0.00114、平均核苷酸差異1.221。共定義了4個(gè)單倍型,其中Hap2為優(yōu)勢(shì)單倍型,代表17個(gè)個(gè)體,占總體個(gè)數(shù)的58.6 %。
2.2.2 西伯利亞鱘 從表2~3還可看出,基于Cytb基因,西伯利亞鱘親魚(yú)養(yǎng)殖群體多態(tài)性位點(diǎn)數(shù)為5個(gè)、單倍型多樣性0.151、核苷酸多樣性0.00036、平均核苷酸差異0.385。共定義了3個(gè)單倍型,其中Hap1為優(yōu)勢(shì)單倍型,代表24個(gè)個(gè)體,占總體個(gè)數(shù)的92.3 %。
基于D-loop基因,西伯利亞鱘親魚(yú)養(yǎng)殖群體多態(tài)性位點(diǎn)數(shù)為2個(gè)、單倍型多樣性0.385、核苷酸多樣性0.00040、平均核苷酸差異0.400。共定義了3個(gè)單倍型,其中Hap1為優(yōu)勢(shì)單倍型,代表20個(gè)個(gè)體,占總體個(gè)數(shù)的76.9 %。
2.3 中性檢驗(yàn)
從表4看出,西伯利亞鱘群體基于Cytb和D-loop的Tajima’s D值分別為-2.0020和-0.50721,F(xiàn)u’s Fs值分別為-0.527和-0.445,可推測(cè)在西伯利亞鱘親魚(yú)群體中低頻率等位基因占主導(dǎo)作用。
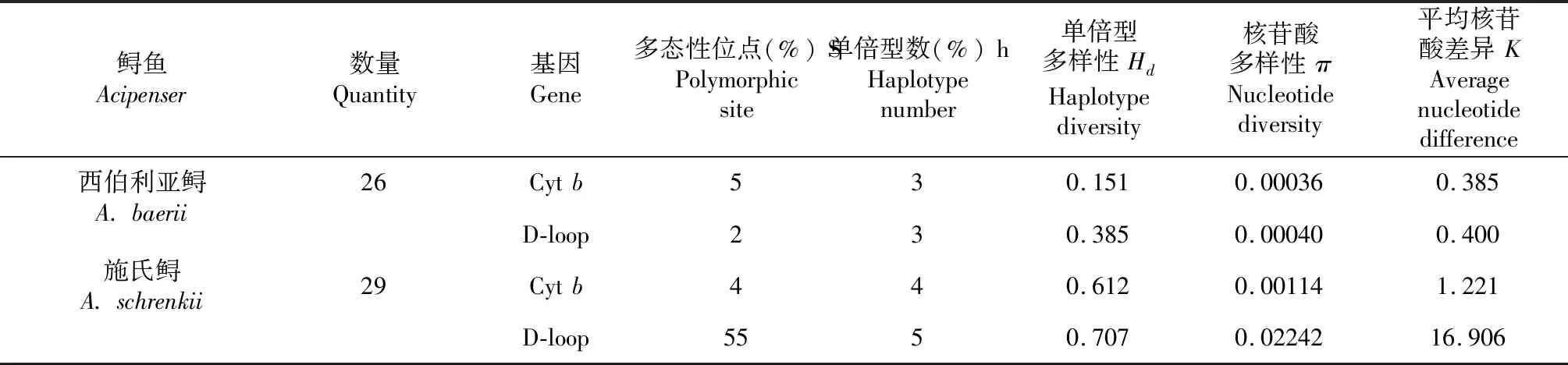
表2 西伯利亞鱘和施氏鱘的遺傳多樣性參數(shù)Table 2 Genetic diversity of A. baerii and A. schrenkii population
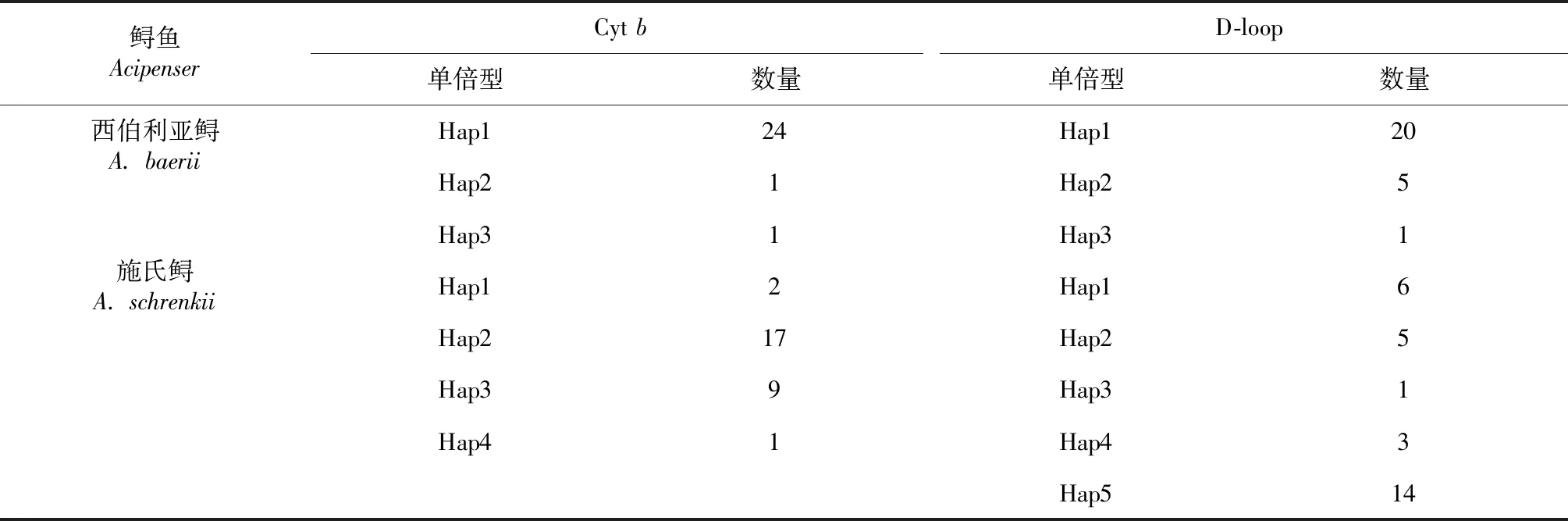
表3 基于Cyt b和D-loop序列的單倍型在西伯利亞鱘和施氏鱘群體中的分布Table 3 Distribution of haplotype in Cyt b and D-loop sequences of A. baerii and A. schrenkii population
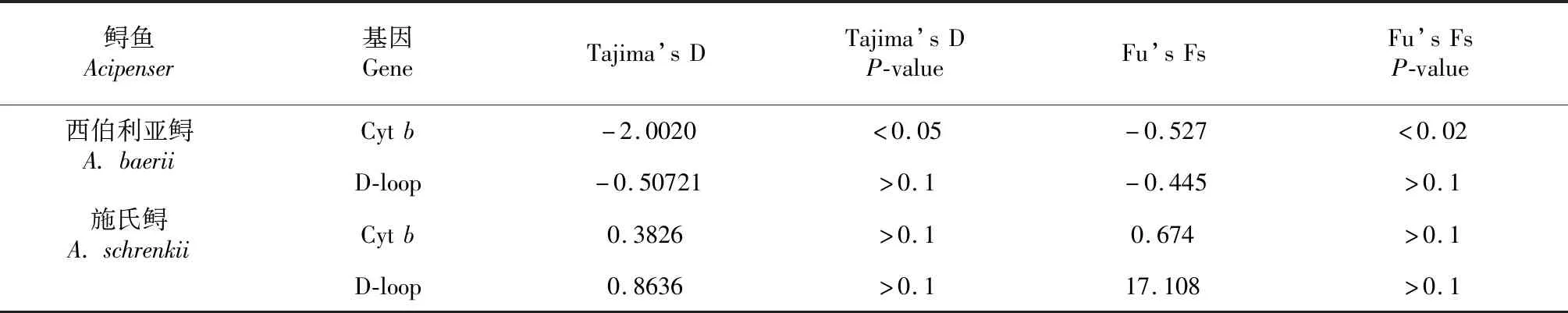
表4 西伯利亞鱘和施氏鱘的中性檢驗(yàn)結(jié)果Table 4 Neutrality test of A. baerii and A. schrenkii population
施氏鱘群體基于Cytb和D-loop的Tajima’s D值分別為0.3826和0.8636,F(xiàn)u’s Fs值分別為0.674和17.108,且其P值不顯著,表明該群體未經(jīng)歷過(guò)種群擴(kuò)張。
3 討 論
3.1 西伯利亞鱘與施氏鱘的Cyt b、D-loop序列信息
線(xiàn)粒體基因組中A、T、G、C的分布呈不均一性[15],而堿基A+T含量高的線(xiàn)粒體基因在進(jìn)化中更具優(yōu)勢(shì)[16]。本研究西伯利亞鱘、施氏鱘D-loop的A+T含量分別為64.3 %和61.8 %,大于其Cytb的A+T含量,表明堿基組成具有偏倚性。施氏鱘群體D-loop的A+T含量高于牛翠娟等[6]研究的施氏鱘群體(57.9 %),與王巍等[5]研究的施氏鱘群體(61.8 %)和西伯利亞群體(62.2 %)的含量相當(dāng)。由此推斷,本研究施氏鱘和西伯利亞鱘群體的D-loop比Cytb基因在進(jìn)化中更具優(yōu)勢(shì)。
在生物體進(jìn)化中顛換不斷積累,因此轉(zhuǎn)換/顛換比不斷減小,一般認(rèn)為轉(zhuǎn)換/顛換比小于2時(shí),基因序列突變已經(jīng)達(dá)到飽和狀態(tài)[17]。本研究西伯利亞鱘D-loop的轉(zhuǎn)換/顛換比小于2,表明突變已達(dá)飽合狀態(tài)。
基于Cytb和D-loop分析施氏鱘群體的Tajima’s D和Fu’s Fs值為正值,表明施氏鱘群體的中等頻率等位基因占主導(dǎo),該種群在經(jīng)歷瓶頸時(shí)使稀有等位基因丟失,且其P值不顯著,表明該群體未經(jīng)歷過(guò)種群擴(kuò)張。基于Cytb和D-loop分析西伯利亞鱘群體的Tajima’s D和Fu’s Fs值均為負(fù)值,表明西伯利亞鱘群體中存在許多稀有等位基因,可能是在定向選擇中削弱了原有等位基因在群體中的頻率。
3.2 西伯利亞鱘與施氏鱘的遺傳多樣性
遺傳多樣性是魚(yú)類(lèi)遺傳育種的一個(gè)重要參考指標(biāo),其高低意味著生物適應(yīng)生存能力的強(qiáng)弱,多樣性越高生物體便有越豐富的育種和遺傳改良潛力[18-19]。衡量一個(gè)種群mtDNA的遺傳多樣性有2個(gè)重要指標(biāo):?jiǎn)伪缎投鄻有?Hd)和核苷酸多態(tài)性(π)[20]。π值考慮了各種mtDNA單倍型在群體中所占的比例,因此反映一個(gè)群體多態(tài)程度往往比單純的遺傳距離平均值更準(zhǔn)確,π值高則說(shuō)明種群的遺傳多樣性較高[21]。
牛翠娟等[6]通過(guò)D-loop序列對(duì)施氏鱘群體的遺傳多樣性分析顯示,核苷酸多樣性(π)為0.011,單倍型多樣性(Hd)為0.768,認(rèn)為后備親魚(yú)有一定的遺傳變異程度。王巍等[5]對(duì)鱘魚(yú)的遺傳多樣性分析顯示,施氏鱘(π=0.002,Hd=0.608)、西伯利亞鱘(π=0.005,Hd=0.706)后備親魚(yú)群體遺傳多樣性偏低。從其他魚(yú)類(lèi)的遺傳多樣性看,香魚(yú)群體[22]D-loop的π=0.00199、Hd=0.81075,Cytb的π=0.00028、Hd=0.323,其群體遺傳水平較低;赤眼鱒野生群體[11]D-loop的π=0.0284、Hd=0.963,Cytb的π=0.0426、Hd=0.9710,具有較高的遺傳多樣性水平。本研究基于mtDNA Cytb基因、D-loop序列研究的遺傳多樣性顯示,西伯利亞鱘Cytb的π=0.00036、Hd=0.151,D-loop的π=0.0040、Hd=0.385;施氏鱘Cytb的π=0.00114、Hd=0.612,D-loop的π=0.02242、Hd=0.707;施氏鱘群體的遺傳多樣性略高于西伯利亞鱘群體,但與上述多樣性豐富的魚(yú)類(lèi)相比,西伯利亞鱘、施氏鱘后備親魚(yú)群體的遺傳多樣性均較匱乏。因此,在繁殖中應(yīng)注意引進(jìn)其他產(chǎn)地的親魚(yú),以增加現(xiàn)有后備親魚(yú)群體的遺傳多樣性。
本研究初步反映了西伯利亞鱘、施氏鱘親魚(yú)群體的遺傳背景、遺傳結(jié)構(gòu)以及遺傳變異水平,但mtDNA標(biāo)記與核基因標(biāo)記并不成正相關(guān)關(guān)系。可以結(jié)合核基因組分子標(biāo)記如SSR、SNP,以更全面的展示該群體的遺傳多樣性,為親魚(yú)選育、苗種培育提供更充分的理論依據(jù)。
4 結(jié) 論
施氏鱘和西伯利亞鱘群體的D-loop比Cytb基因在進(jìn)化中更具優(yōu)勢(shì);西伯利亞鱘D-loop區(qū)突變已達(dá)飽合狀態(tài);施氏鱘群體的中等頻率等位基因占主導(dǎo),該群體未經(jīng)歷過(guò)種群擴(kuò)張,西伯利亞鱘群體中存在許多稀有等位基因;西伯利亞鱘、施氏鱘后備親魚(yú)群體的遺傳多樣性均較匱乏。
