茶樹白化突變研究進展
2020-04-25 07:15:28劉丁丁梅菊芬王君雅湯榕津陳亮馬春雷
中國茶葉 2020年4期
劉丁丁,梅菊芬,王君雅,湯榕津,陳亮,馬春雷*
1.中國農業科學院茶葉研究所國家茶樹改良中心 農業農村部茶樹生物學與資源利用重點實驗室,310008;2.中國農業科學院研究生院,100081;3.江蘇省茶葉研究所 江蘇省茶樹種質資源圃,214063
茶樹起源于我國西南地區,作為一種多年生木本經濟作物,在世界范圍內廣泛種植[1-2]。隨著茶樹栽培和選育技術的發展,育種者們發掘和培育了許多優良的茶樹品種,其中白化茶樹品種加工而成的茶葉因具有較高的氨基酸含量和優質的外形在市場上廣受歡迎。我國自唐朝起就有對白化茶的介紹,歷史上記載的描述白化茶樹的史料有許多,如唐朝陸羽《茶經》“七之事”中記載“永嘉圖經,永嘉縣東三百里有白茶山”;北宋宋子安《東溪試茶錄》中記載“茶之名有七,一曰白葉茶,民間大重,出于近歲,園焙時有之,地下以山川遠近,發不以社之先后。芽發如紙,民間以為茶瑞”。1864年張星煥《皖游紀聞》中記載“蘭生言,石埭某山近日間產白茶。其味絕殊,但不可多得,或千百株中偶有一株變白,或今年白,明年仍可復原色,土人以為瑞茶,得則珍藏之”;而產于慧苑巖火焰峰下的白雞冠是延續至今的適制烏龍茶的白化茶樹資源[3]。現代對白化茶樹種質資源的研究,要追溯到上世紀七八十年代,研究者最初主要從白葉1號組織結構、生理生化等角度開展白化茶樹資源的相關研究;近十幾年,隨著組學技術的快速發展,有大量利用轉錄組、代謝組、蛋白質組等技術對這一表型進行研究的文章發表。本文從白化茶樹資源的分類、生理生化特征、分子機制等方面對相關研究進展進行了綜述,以期為茶樹白化機制的解析提供參考。
一、白化茶樹突變體分類
白化茶樹是一類在特定環境條件下茶樹新梢呈現白化表型的茶樹突變體。根據誘導白化產生的因素,現有白化資源主要分為生態敏感型、生態不敏感型、生態復合型和未知型幾類。生態敏感型白化茶樹主要是對氣候因子比較敏感,可分為低溫敏感型和光照敏感型(表1),其中低溫敏感型茶樹的白化現象主要是受早春茶芽萌發時的溫度影響,并且隨著溫度的升高,白化表現減弱[4],這種類型的茶樹在自然栽培條件下一般只在春末夏初發生“返白—復綠”現象,如白葉1號、千年雪等。而對于光照敏感型白化茶樹,多數資源全年表現為白化現象,如黃金芽、御金香等。與生態敏感型白化茶樹的“階段性白化”不同的是,生態不敏感型白化茶樹的白化現象屬于“恒定性白化”,即表現為白者恒白,綠者恒綠的現象[5],如花月等。生態復合型白化茶樹是指茶樹組織的一部分依賴溫度或者光照的變異,一部分對生態因子不敏感,如金玉緣、春雪3 號等。而對于未知類型的白化茶樹資源,筆者認為屬于上述3種類型之一,但仍需進一步的試驗驗證。

表1 白化茶樹資源統計
對目前已有報道的白化茶樹資源進行綜合分析可發現,白化茶樹也可依據芽葉顏色進行分類,具體可分為以白色系和黃色系為主的純色類型和以斑馬葉、條紋葉、斑點葉為主的非純色類型(圖1)。其中白色系茶樹主要表現為低溫敏感型,黃色系茶樹主要表現為光照敏感型;在氨基酸含量上,白化茶樹資源的氨基酸含量較常規綠色茶樹品種高,但也并不是所有的白化茶樹資源氨基酸含量都高。表明了這些資源雖同為葉色突變體,但是不同資源之間的葉色形成機制及生理生化特征存在一定的差異,也正是這種差異的存在為我們進行新種質的創制提供了豐富的材料。
二、白化茶樹生理生化特征
植物葉片的色澤是葉綠體體內的光合色素和液泡中的花青素綜合作用的結果,其中光合色素在光合作用中具有吸收、傳遞、轉化光能的作用,其含量是決定光合效率的重要因素[34]。葉綠體不僅能通過光合作用為植物的生長發育提供養料,同時也是葉綠素、類胡蘿卜素和葉黃素等色素物質合成的場所,因此其形態結構與植物葉片色澤的變化具有緊密的聯系[35]。所以研究白化茶樹突變體葉綠體結構變異以及光合性能變化是解答茶樹白化機理的基礎。

圖1 不同葉色的茶樹資源
1.葉綠體超微結構
杜穎穎[36]利用透射電鏡觀察了處于15℃、20℃、25℃條件下的小雪芽葉片中葉綠體的超微結構,發現在15℃條件下,葉肉細胞內無成熟葉綠體存在,可觀察到大量的白色體,但白色體內無類囊體和基粒的堆疊。在20℃和25℃條件下,葉肉細胞內均觀察到發育完整的葉綠體結構,內有發育良好的類囊體和緊密堆疊的基粒片層,不同的是在20℃條件下觀察到的葉綠體膜有損傷,推測可能是此時葉綠體膜的發育還不完全。李素芳等[37]利用透射電鏡觀察白葉1號階段性“白化—返綠”過程中葉綠體的超微結構,發現在返白階段葉綠體不斷退化解體、內部空泡化,在復綠階段葉綠體結構又重新恢復正常,并提出白葉1 號的白化現象可能是由核基因或核質互作引起的[38]。李娜娜[39]則以白色系資源小雪芽為材料,研究發現其不同葉位葉綠體超微結構的變化與白葉1 號不同白化階段的結果極其相似,在白色葉片中也出現了葉綠體解體和內部空泡化的現象,而低葉位的偏綠色葉片葉綠體結構又恢復了正常。韋康等[40]以光照敏感型白化品種中黃2 號和正常綠色品種龍井43 為試驗材料比較葉綠體的超微結構,結果發現中黃2 號葉綠體結構完整,沒有出現解體或大量囊泡,但是基粒片層堆疊少于龍井43。由此可以看出,幾乎所有白化茶樹葉色突變體的形成,在細胞結構層面都伴隨著葉綠體空泡化、類囊體發育不完全、白色體的產生和基粒片層結構缺乏等現象,但它們的相互關系還有待進一步研究。
2.光合作用
光合作用為茶樹碳、氮代謝提供碳源和能量,同時也是決定生產力高低的重要指標。但是茶樹的光合能力不僅受到環境的影響,也受品種自身遺傳因素的制約[41]。到目前為止,對白化茶樹品種在生理生化、葉色相關基因表達調控等方面已有一些研究,但是對白化茶樹的光合生理特性研究還較少。李勤等[42]從凈光合效率、氣孔導度、胞間CO2濃度和蒸騰速率等角度分析白葉1號在“返白—復綠”幾個階段的光合作用變化,發現在白化階段(白化前期、白化中期、白化期)葉片的凈光合速率、氣孔導度、蒸騰速率呈現先升高后下降的趨勢,而胞間CO2濃度則呈現先下降后上升的趨勢;在復綠階段(復綠前期、復綠中期、全綠期)葉片的凈光合速率、氣孔導度、蒸騰速率表現為逐漸上升的趨勢,胞間CO2濃度表現為逐漸下降趨勢。胞間CO2濃度在階段性“返白—復綠”過程中的變化和凈光合速率、氣孔導度、蒸騰速率的變化呈現相反的趨勢,這可能與葉片的發育程度和葉綠體的結構有關。Dong等[43]測定了黃金芽在自然光照和遮陰條件下葉片的凈光合速率、氣孔導度、胞間CO2濃度、蒸騰速率,發現和遮陰條件下的綠色葉片相比較,自然光照條件下的黃色葉片凈光合速率、胞間CO2濃度均降低,而蒸騰速率和氣孔導度均升高。由此可看出,白葉1 號和黃金芽雖同為葉色變異資源,但其光合性能還是有較大區別的。
3.生化成分
茶樹作為一種葉用作物,葉片生化成分的種類和含量是評價茶葉品質和茶葉功能的重要指標[44]。對不同白化茶樹進行生化水平上的分析,有助于了解不同白化茶樹資源的特征特性,從而更好地指導相關資源的開發利用。
(1)主要代謝物
氨基酸、多酚類物質和咖啡堿等是茶樹中重要的代謝物質,也是茶葉中重要的呈味物質,對茶樹生長發育和茶葉品質影響很大。而白化茶樹資源普遍表現為高氨基酸含量、適中的茶多酚,但在不同的處理下,其生化物質也會發生明顯的變化。如Li 等[45]對黃金芽進行遮陰處理,結果表明,遮陰處理的黃金芽新梢咖啡堿含量高于自然光照下的黃金芽新梢中的含量;隨著遮陰程度增加,黃金芽新梢中總氨基酸含量增加;而自然光照下,新梢中茶多酚總量明顯低于單層遮陰的黃金芽,但高于雙層遮陰的黃金芽。這說明遮陰有利于咖啡堿的積累和氨基酸含量的增加,但是過度遮陰會引起茶多酚含量的降低,這可能是遮陰導致茶樹體內碳氮代謝發生變化,進而引起各種次級代謝物質含量發生變化的原因。此外,Lu等[46]以一個“帶白化枝條的正常綠色茶樹”為材料,比較白化枝條和綠色枝條中次級代謝物質的變化。研究表明,與綠色葉片相比,白化葉片中EGCG、EGC、ECG、EC 含量大幅度降低;咖啡堿含量略微降低;而游離氨基酸,如茶氨酸、谷氨酸、谷氨酰胺、精氨酸等顯著升高。杜穎穎[36]以小雪芽、白葉1 號、千年雪、黃金芽和福鼎大白茶為材料,研究白化資源與綠葉品種中茶多酚和咖啡堿含量的差異,結果表明,白化資源中茶多酚和咖啡堿含量顯著低于綠葉品種福鼎大白茶。
(2)色素含量
葉綠素和類胡蘿卜素作為光合色素,不僅在光合作用中起到捕獲、吸收、傳遞光能的作用,同時作為主要的脂溶性色素,其含量的高低直接決定茶樹葉片的顏色,進而影響茶葉干茶和葉底色澤的形成。Cheng等[47]以綠色正常茶樹品種英紅9號及其白化突變體為材料,在春季和冬季分別測定了葉綠素a和葉綠素b的含量,發現兩個季節的白化突變體中葉綠素a和葉綠素b的含量均顯著低于正常茶樹品種中的含量。Li 等[45]以黃金芽為研究材料,對其進行遮陰處理,研究遮陰處理前后黃金芽的各種色素含量的差異。結果表明,遮陰處理綠色葉片的葉綠素a、葉綠素b和葉綠素總量均明顯高于光照下黃色葉片的含量;而遮陰處理綠色葉片的紫黃質、新黃質、葉黃素和類胡蘿卜素總量均低于光照下黃色葉片的含量,分析認為光照強度增強可能刺激了類胡蘿卜素的積累。Wang 等[48]分別測定中黃2 號和龍井43 中的葉綠素、類胡蘿卜素和葉黃素的含量,發現中黃2 號中葉綠素、類胡蘿卜素和葉黃素的含量均顯著低于龍井43。
(3)酶活性
研究發現,白化茶樹新梢在白化時期體內酶活性會發生顯著的變化,主要表現為1,5-二磷酸核酮糖羧化酶活性下降,蛋白水解酶活性增強;在復綠時期兩種酶活性變化與白化時期相反。李素芳[49]研究白葉1號各白化時期和復綠時期酶活性與氨基酸含量的關系時發現:隨白化程度加深,蛋白水解酶活性增強,將體內多余蛋白質水解,導致游離氨基酸含量增加,推測這可能是白葉1 號在白化時期氨基酸含量較高的原因。
過氧化物酶(POD)、超氧化物歧化酶(SOD)、過氧化氫酶(CAT)作為茶樹體內的保護酶,在茶樹白化期間其活性也會發生變化。陸建良等[50]研究發現,在白葉1號返白過程中,POD酶活力升高,SOD 和CAT 酶活力降低,而當芽梢復綠后,POD 酶活力降低,CAT、SOD 酶活力回升;Wu等[51]在研究光照和遮陰處理條件下白雞冠葉片中POD、SOD、CAT 酶活性變化時,也得出相同的結論。推測造成此類現象的原因可能是葉片內葉綠體結構發育異常,打破了活性氧平衡,使茶樹啟動了保護酶機制。
三、組學技術在白化茶樹資源研究中的應用
近些年來,隨著各種組學技術的快速發展,將茶學的相關研究帶入了一個新的高度。截至目前,已有2 個茶樹基因組草圖相繼發表[52-53],標志著茶學研究正式進入了后基因組時代,這為不同白化茶樹分子機制的解析提供了新的研究方向。
1.基因差異表達
基因差異表達作為功能基因組學的重要組成部分,是從RNA水平上研究生物體在不同組織結構、不同發育時期、不同環境條件下基因的表達情況。在白化資源的研究中,相關的分析方法已得到廣泛應用。通過將不同葉色新梢差異表達的基因進行Blast、GO、KEGG等分析,可初步了解引起白化表型的基因有哪些,以及它們主要富集在哪些代謝通路中,從而為探究白化茶樹資源的白化分子機制提供支撐。
因白葉1 號是較早發現的葉色變異資源,所以針對它的相關研究也最為系統,不同階段的差異表達技術在這一資源上均有應用,如王新超等[54]利 用DDRT-PCR (Differential display of reverse transcriptional PCR)技術從白葉1 號不同葉色新梢中獲得58 條差異表達片段,進一步通過RT-PCR分析,驗證了12條陽性片段;Blastx序列比對發現,5個陽性片段分別與編碼血紅素結合蛋白、甲硫氨酸合酶、1-氨基環丙烷羧酸合酶等的基因序列高度相似。李娟[55]利用SSH(Suppression subtractive hybridization)技術從白葉1號綠色葉和白化期葉片中分離80個差異表達基因,進一步分析篩選到1個可能調控白葉1號返白現象的鈣調熱激蛋白基因和2 個與高氨基酸性狀相關的絲氨酸-乙醛酸轉氨酶、絲氨酸羥甲基轉移酶基因。吳 揚[56]、Ruan 等[57]分 別 利 用cDNA-AFLP (cDNA-amplified fragment length polymorphism)技術從白葉1 號不同葉色新梢中獲得了31 個和127 條差異表達片段,對這些差異表達的基因進行同源序列比對發現,均與細胞周期、信號傳導、物質和能量代謝等相關。Ma等[58]利用基因芯片技術從白葉1號不同葉色發育時期的葉片中獲得671個差異表達基因,這些基因主要參與到能量代謝、碳固定、次級代謝、植物生長和防御等過程,并鑒定了一些參與葉綠素合成和葉綠體發育的催化酶和調控蛋白,其中發現編碼鎂-螯合酶基因的表達趨勢和葉色性狀的變化相一致。Li等[59]對白葉1號黃綠期、白化期、返綠期3 個時期的新梢進行RNA-Seq 測序,通過兩兩比較分析差異表達基因,這些差異表達基因主要參與到代謝途徑、次級代謝物生物合成、苯丙烷代謝和碳固定途徑中。
近些年,隨著葉色變異資源的增多,基因差異顯示技術也開始在更多的品種資源中得到應用(表2),如李娜娜等[60]利用RNA-Seq法對福鼎大白茶和小雪芽葉片的差異基因進行了研究,篩選到1 821 個上調表達基因和1 779 個下調表達基因,進一步對差異基因進行GO 和COG 分析,認為引起小雪芽新梢白化的遺傳機制可能與蛋白質翻譯后修飾過程有關。Wu等[51]利用RNA-Seq技術對白雞冠的白化葉片和綠色葉片的差異表達基因進行研究,這些差異表達基因主要參與葉綠體發育、活性氧清除、光合色素合成、次級代謝和晝夜節律調控過程。一些科研人員也分別利用RNA-Seq技術研究了御金香、華白1 號、黃金芽、炎陵銀邊茶等不同葉色時期的差異表達基因,結果發現差異表達基因同樣參與了葉綠體發育、光合色素合成、次級代謝等過程[61-66]。這說明雖然不同白化資源的遺傳背景并不相同,但在基因表達層面上,葉綠體發育異常和葉綠素合成受阻是導致這類資源白化表型產生的直接原因。

表2 白化茶樹資源的基因差異表達研究
2.蛋白質組學
蛋白質組學是以生物體內基因所表達的全部蛋白質為研究對象,從整體、動態的角度研究蛋白質特征,從而幫助科研人員從蛋白質水平上整體而全面地理解生物體的組織變化、物質代謝等過程[67-68]。目前,基于雙向凝膠電泳、質譜的蛋白質組學技術在白化茶樹資源的研究中已得到初步應用(表3)。如Li 等[69]在白葉1 號3 個葉色發育階段利用基質輔助激光解吸電離飛行時間質譜(Matrix-assisted laser desorption/ionization time of flight mass spectrometry,MALDI-TOF-MS)技術鑒定了26個生育期差異表達蛋白,這些差異表達蛋白主要參與碳氮硫代謝、光合作用、RNA 和蛋白質加工、脅迫防御等過程;之后同樣在白葉1 號3個葉色發育階段利用質譜鑒定了22個葉綠體差異表達蛋白,發現與光合作用、物質代謝、核酸代謝、應激響應等相關[70]。Xu 等[71-72]從轉錄后修飾角度,利用LC-MS/MS (Liquid chromatography-tandem mass spectrometry)技術先后從白葉1號不同白化時期的表達蛋白中鑒定了3 161 個乙酰化位點和3 530 個琥珀酰化位點,其中與光合作用、碳固定、氨基酸生物合成和葉綠素代謝相關蛋白的琥珀酰化水平在各個白化時期存在差異,同時與光合作用和次級代謝相關蛋白的乙酰化水平也發生了變化,而且部分琥珀酰化和乙酰化蛋白的作用存在部分重疊。除了對低溫敏感型白葉1 號進行研究之外,還有研究分別利用同位素相對標記和絕對定量技術(Isobaric tags for relative and absolute quantitation, iTRAQ)分別鑒定了光照敏感型的黃金芽和中黃1 號的173 個、56 個、437 個差異表達蛋白,大多數蛋白均參與光合作用、類黃酮代謝、碳水化合物的轉運與代謝、次級代謝物的合成等過程[43,73-74]。
3.代謝組學
代謝組學可以通過定性或定量分析生物體在不同發育時期、不同組織結構、不同環境條件下代謝產物的變化情況,從而整體揭示生物體的代謝調控機制和生命活動規律。近些年來,研究者們利用代謝組學技術與多元統計分析相結合,發現不同葉色茶樹品種之間、同一茶樹品種不同葉色之間的代謝物變化均存在顯著的差異(表4)。如Zeng 等[75]利用UPLC-QTOF-MS(Ultra-high performance liquid chromatography-quadrupole time-of-Flight mass spectrometry)技術在白葉1 號3 個葉色發育階段共鑒定了92 種差異代謝物,這些差異代謝物主要參與類黃酮、苯丙烷、氨基酸等的生物合成途徑,并發現在白化期內氨基酸含量較高、碳水化合物含量較低,兒茶素和黃酮類物質含量在白化前期和返綠期含量較高;Li等[76]利用LC-MS技術同樣在白葉1號的3個葉色發育階段鑒定了65個差異代謝物,這些差異代謝物與半乳糖、色氨酸代謝和苯丙烷、類黃酮生物合成等相關。Zhang等[77]在黃金芽白化葉片和綠色葉片中共鑒定了52個差異代謝物,與類黃酮物質生物合成和苯丙烷代謝相關,并發現CG、槲皮素和黃酮醇苷類物質在白化葉片中大量積累,而楊梅苷和EGCG含量則較低;之后Zhang等[78]又鑒定了41 個差異代謝物,與氨基酸和碳骨架代謝相關,其中在白化葉片中谷氨酸、谷氨酰胺、茶氨酸、檸檬酸和α-酮戊二酸含量增加,而單糖、二糖和咖啡堿含量降低。Shen 等[79]為解析白化葉片氨基酸代謝增強的機制,利用HPLC (High performance liquid chromatography)檢測了黃山白茶白化前期、芽期、白化期和返綠期新梢中的游離氨基酸含量,并以綠色品種黃山種為對照,同時利用qRT-PCR 技術檢測了參與氨基酸代謝的基因在幾個不同時期的表達量,結果表明茶氨酸合成增強和氨基酸轉運能力降低是導致白化時期新梢中氨基酸含量增加的主要原因。此外,Feng 等[80]利用HPLC 技術研究了不同葉色品種之間的代謝譜差異,發現和綠色品種福鼎大白茶相比,白化品種白葉1 號、黃金芽、御金香葉片中含有較低的葉綠素、類胡蘿卜素、咖啡堿、兒茶素,而玉米黃素和茶氨酸含量較高。
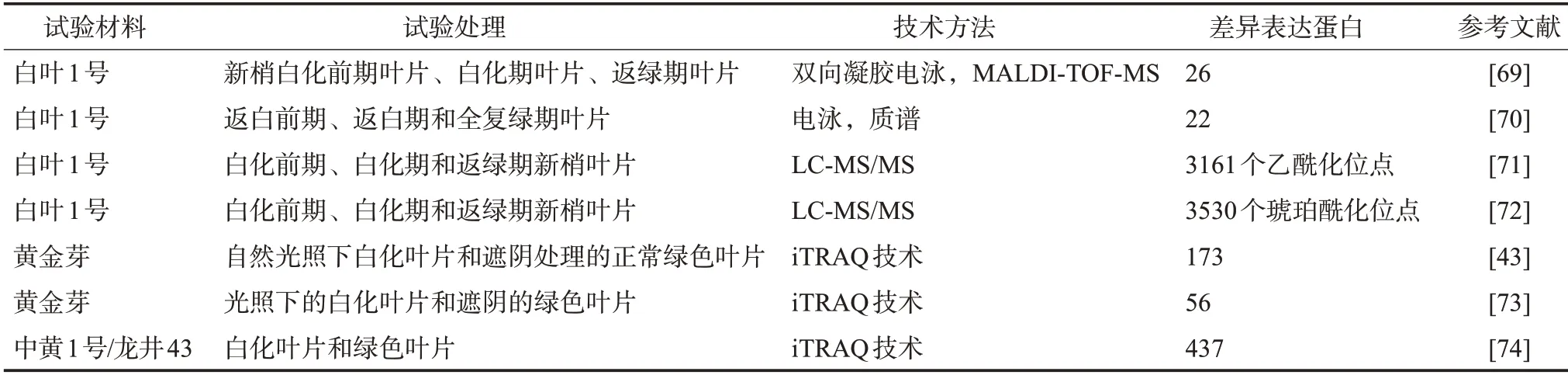
表3 白化茶樹資源的蛋白質組學研究
目前,利用代謝組學技術研究白化茶樹資源的代謝物變化,主要關注氨基酸、黃酮類和糖類等物質,而對香氣物質的報道很少。根據已有研究,Dong 等[81]利用GC-MS 技術分析了4月、8月和12月份英紅9 號白化葉片和綠色葉片中香氣物質的變化,發現3 個時期中白化葉片的順-3-乙烯醇和芳樟醇含量均顯著低于綠色葉片,其他香氣物質含量變化不明顯;這可能就是導致白化葉片中香氣物質含量低于綠色葉片的直接原因。

表4 白化茶樹資源的代謝組學研究
四、分子標記技術在白化資源研究中的應用
隨著測序技術的快速發展,也推動了DNA分子標記技術的廣泛應用,與形態學標記、同工酶標記相比,DNA分子標記直接從基因組水平上研究生物的遺傳變異,不受發育時期、環境因素的限制[82]。目前分子標記技術在白化茶樹資源研究中主要應用于遺傳多樣性分析、品種鑒定、遺傳圖譜構建及QTL定位等方面。
王松琳[83]等利用SSR (Simple sequence repeats)分子標記對16個白化茶樹品種進行遺傳多樣性分析,通過分析等位基因數、多態性信息含量和shannon 值等,發現品種間遺傳結構差異明顯。并篩選出3 對核心SSR 引物,用于品種鑒定。王開榮等[3]利用RAPD(Random amplification polymorphic DNA)技術結合DNA序列測定,以福鼎大白茶為對照,研究黃金芽、千年雪、御金香、白葉1 號、四明雪芽、金光和花月等7 個白化品種的特異DNA 分子標記,以期找到與白化相關的基因。通過對單一品種唯一擴增位點和缺失位點分析,發現一些位點可能與葉綠素代謝和抑制與葉綠素代謝相關基因的表達有關;進一步篩選了21個特異分子標記,并成功測序了4條DNA序列,通過Blast序列比對發現與蛋白激酶、轉運蛋白家族基因等具有同源性;同時這一分子標記也可應用于白化茶樹品種的遺傳鑒定。吳永勝[84]以白葉1號、黃金芽、御金香、四明雪芽等10份茶樹資源為材料,先利用50對RAPD反應體系及程序進行PCR 擴增,引物S20 在御金香中擴增出1條特異性片段,之后在引物S20基礎上設計SCAR引物SY20,用于擴增10份材料,發現該引物同樣在御金香中擴增出特異性片段,表明了RAPD標記成功地轉化為SCAR標記,并且利用該標記能夠將御金香從多種茶樹資源中鑒別出來。
葉綠素、類胡蘿卜素等色素物質直接影響茶樹新梢葉片色澤,屬于數量性狀。為研究其遺傳機制,王松琳[35]以白雞冠和龍井43 作為父母本構建遺傳群體,對親本和子代個體測序獲得大量SNP(Single nucleotide polymorphisms)標記,結合擬測交策略構建遺傳圖譜,隨后利用遺傳圖譜對葉綠素和類胡蘿卜素這2 個性狀進行QTL(Quantitative trait locus)作圖,結果共定位了10個QTL,分布于4 個連鎖群上,表型貢獻率在18.2%~72.0%,均為主效QTL。
五、展望
綜上所述,茶樹白化現象的產生與細胞色素的合成和降解、光合系統、葉綠體的發育等密切相關。在生理水平上表現為葉綠體解體或退化消失,進而影響了葉綠素和類胡蘿卜素等色素的生物合成;在生化成分上體現為含有較高含量的游離氨基酸;但是在分子水平上茶樹的白化機理仍不明確。目前,雖也有部分參與葉綠素、類胡蘿卜素、氨基酸合成相關的茶樹基因得到了分離克隆,如鄧婷婷等[85-86]根據cDNA-AFLP 的結果,利用RACE(Rapid amplification of cDNA ends)技術克隆了泛素活化酶基因和八氫番茄紅素脫氫酶3 基因;馬春雷等[87]根據芯片雜交結果,克隆了3 個參與葉綠素合成途徑的基因:谷氨酸-tRNA 還原酶基因、葉綠素合酶基因、葉綠素醋酸氧化酶基因。但相關研究均屬于同源克隆,因此到底是哪個基因的突變導致了白葉1 號、黃金芽等白化突變體的白化表型問題仍未有答案。
從已報道的研究結果來看,在茶樹白化突變體研究中還存在諸多亟待解決的問題。
首先,雖然近些年各地陸續發現了很多的白化茶樹突變材料,但大多數資源的譜系關系和遺傳背景均不清楚;以這些突變體為研究材料的相關研究也多集中在生理生化、基因表達和代謝層面,而缺少遺傳層面的深入分析。
第二,因茶樹是多年生木本植物,生長周期長,導致構建白化相關遺傳群體的難度很大;而茶樹基因組龐大,遺傳位點高度雜合,使得茶樹白化性狀的遺傳解析困難重重。
第三,因茶樹轉基因技術尚不成熟,很難通過本源表達驗證基因功能,從而導致白化相關基因的鑒定很難開展。針對這些問題,課題組從2013年開始,對保存在國家種質杭州茶樹圃的十幾份白化茶樹資源的遺傳特性進行了連續多年的觀察,發現這些資源中的大多數都具有葉色遺傳規律,如黃金芽、白雞冠、安吉黃茶、中黃1 號等(表5)。

表5 不同白化品種資源的遺傳特性分析
隨后課題組以其中3 個具有葉色遺傳特性的白化資源為父本,通過多年的連續雜交,已建立了3 個白化性狀在子代中能有效分離的F1 群體,這些遺傳群體將是以后解答白化茶樹資源葉色突變機制的重要研究材料。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
音樂探索(2022年2期)2022-05-30 21:01:37
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
資源再生(2017年3期)2017-06-01 12:20:59
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
