全球抗新型冠狀病毒藥物研發現狀和瓶頸
2020-04-26 02:42:48陳丹龍羅志英謝煥章董海燕
中國藥理學通報 2020年4期
陳丹龍,楊 芳,羅志英,謝煥章,董海燕,賈 力,
(1.閩江大學海洋研究院,福建 福州 350108;2. 福州大學化學院、福建省腫瘤轉移藥物預防重點實驗室,福建 福州 350116;3.福建醫科大學轉化醫學研究院、福建省癌癥和神經退行性疾病轉化研究重點實驗室,福建 福州 350108 )
1 引言
2003年2月我國廣東暴發的嚴重急性呼吸系統綜合征(severe acute respiratory syndrome,SARS)和2012年9月在沙特暴發的中東呼吸綜合征(middle east respiratory syndrome,MERS)分別由SARS-CoV和MERS-CoV冠狀病毒引發。根據WHO的公開資料,我國SARS確診8098人,喪生774人,但在一年內就完全遏制了。而MERS從2012年9月一直延續到2019年,WHO報告了2 468例確診和851例死亡。埃博拉病毒在2013年底從幾內亞開始肆虐非洲,并擴展至歐洲多國,截至2015年6月3日,埃博拉病毒已造成11 164人死亡。2019年12月,新型冠狀病毒疾病(COVID-19)疫情在武漢爆發,疫情發展極為迅速,短短1個多月的時間確診感染人數已超過6萬人,死亡1千多人(截止2020年2月15日),并波及全球20多個國家和地區。為了遏制疫情的快速擴散,政府采取了積極有效及時的措施,多管齊下,目前每日新增病例數已經開始下降,疫情的拐點逐漸顯露,也預示著我們有望在短期內基本控制COVID-19,再次彰顯我國政治治理的優越性。
面對2003年后再次襲來的冠狀病毒,以及未來還可能卷土重來冠狀病毒,我們在研發抗冠狀病毒藥物方面吸取了哪些經驗和教訓,為什么至今沒有專門的特效藥產生?未來這類藥物的研發思路和瓶頸是什么?這些都是我們應思考的問題。
病毒是絕對的寄生物種,離開宿主細胞就沒有生命了。同時,病毒對宿主細胞的依賴性引導病毒在其微環境產生適應性的演化和變異。我們只能在具體的宿主樣本發現病毒、研究病毒。如從呼吸道上皮細胞采樣尋找或鑒定流感病毒;在肝炎病人的肝臟和血樣中研究乙肝病毒。在沒有明確宿主情況下做病毒序列分析,是無法確定病毒的真實性能與致病力。另一方面,病毒以人體肺與呼吸道、胃腸道和泌尿生殖道等部位的細菌為宿主而存在。其可能影響細菌微生態的平衡從而間接影響人類健康。
入侵人類的病毒可分為兩類,一類是以人類為長期宿主的病毒群,如人類常見的流行性病毒性疾病:麻疹、天花、皰疹病毒、腮腺炎、甲肝、乙肝、以及人流感病毒,包括HIV、HPV、SARS、Ebola、MERS、Zika等病毒,這類病毒可與人共處,實現人際傳播和疫情。另一類是以臨近人類的物種(雞狗豬馬羊等)為長期宿主但能跨物種感染人類的病毒群,如禽流感病毒、狂犬病毒、漢坦病毒等,這類病毒稱為外源病毒群,在其長期宿主發生疫情時(雞瘟或豬瘟暴發時),病毒才能大量逸出而機遇性跨物種感染人類。
一種新型冠狀病毒的急性傳染期過后,患者可產生相應的持久(或終身)免疫。因此,從藥學經濟學角度考慮,幾乎沒有任何藥企會投巨資研發治療病毒感染的新藥。因為即使幾年后該新藥研發成功上市,其靶向病毒也許不復存在或變異了。因此,雖然目前多種冠狀病毒的分離、結構解析在病毒暴發期就已經完成了,但至今沒有特效藥物。抗冠狀病毒藥物的研發不但要精準靶向病毒本身,還要針對感染后致命的急性呼吸窘迫綜合征 (acute respiratory distress syndrome,ARDS)。所以針對目前和未來冠狀病毒感染的藥物研發必須在了解病毒侵入、復制和釋放的機制及其導致呼吸系統疾病的基礎上,采取“老藥新用”和聯合給藥的策略,才能在短時間內取得突破。本文總結了全球冠狀病毒老藥新用的研發,也提出我們針對抗擊COVID-19的研發新思路。
1.1 SARS、MERS和SARS-Cov-2的流行病學研究冠狀病毒是具外套膜(envelope)的正鏈單股RNA病毒 。它們屬于病毒科冠狀病毒亞科,與圓環病毒亞科一起組成了冠狀病毒科。冠狀病毒分為四種:α-型冠狀病毒、β-型冠狀病毒、γ-型冠狀病毒和δ-型冠狀病毒。冠狀病毒有著相似的基因組結構。開放閱讀框1a和1b接近基因組的2/3,負責編碼非結構蛋白。下游開放閱讀框編碼多種結構蛋白,包括棘突、核衣殼、包膜和膜蛋白[1-3]。SARS-CoV、MERS-CoV、SARS-Cov-2都同屬于β-型冠狀病毒。COVID-19也有包膜,顆粒呈圓形或橢圓形,常為多形性,直徑 60~140 nm。其基因特征與 SARS-CoV 和 MERS-CoV 有明顯區別[4]。
中國的馬蹄蝠是SARS-CoV病毒的疑似宿主[5]。目前尚不清楚人畜共患病的發生和適應機制。從這些蝙蝠中分離出的SARS樣病毒與從人類和果子貍中分離出來的SARS病毒的序列相似性高達95%。在疫情暴發的初始階段,SARS-CoV曾在市場中的棕櫚果子貍中分離出來,但在野生果子貍種群中并未發現[6]。蝙蝠體內有許多冠狀病毒,是感染果子貍或駱駝等中間宿主的宿主,這些中間宿主再把疾病傳播給人類[5]。人與人之間的傳播主要是在醫護人員,及與患者近距離接觸且無保護措施的人群中進行[7-8]。由于MERS-CoV和蝙蝠冠狀病毒是C譜系的一部分,并且在沙特阿拉伯王國的蝙蝠樣本中發現了MERS-CoV RNA,研究人員推測蝙蝠可能是MERS-CoV的天然宿主[9-10]。最近的一項研究結果表明蝙蝠可能是MERS-CoV的宿主,而駱駝和山羊是潛在的中間宿主[11]。在這項研究中,已從感染了MERS-CoV的單峰駝的鼻分泌物中分離得到MERS-CoV,但這些單峰駝的病情發展時間短,病情輕微。
從現有的資料來看, 新型冠狀病毒與蝙蝠病毒 TG13 的全基因組序列一致性高達96%[12]。華南農業大學沈永義團隊對 新型冠狀病毒的可能的動物來源,開展了溯源工作,通過分析測序,對多種野生動物進行了宏基因組學分析,證明穿山甲可能是新型冠狀病毒的潛在中間宿主。
1.2 臨床特征MERS和SARS的臨床特征相似,從無癥狀到輕度癥狀到伴有急性呼吸窘迫綜合征(ARDS),以及多器官功能衰竭的重癥肺炎[13]。雖然MERS和SARS的臨床特征相似,但MERS的死亡率為40%,SARS的死亡率為10%[14]。大約75%的MERS死亡病例伴有并發癥,存在并發癥患者的死亡率為60%(包括心肺異常、肥胖癥和糖尿病)。相比之下,10%~30%的SARS患者伴有并發癥,存在并發癥患者的死亡率為46%[13-14]。
截至2月15日19點30分,全國新型冠狀病毒感染肺炎確診病例共66 580起,其中死亡病例1 524起,病死率為2.2%,低于SARS和 MERS。但各個年齡段的人都可能被感染,被感染的主要是成年人,特別是老年人和體弱多病者。從武漢市早前公開的44個死亡病例中,有30例(68.2%)患有高血壓、冠心病、糖尿病等慢性疾病,也就意味著,本身患有慢性疾病的人對新冠肺炎的抵抗力相對較弱,容易致死。COVID-19最常見的癥狀是發熱、咳嗽、肌痛或疲勞、呼吸困難;影像學特征為肺部CT呈現雙側毛玻璃狀影像。
2 潛在的抗病毒藥物
2.1 臨床應用
2.1.1SARS藥物治療 利巴韋林是一種核苷類似物,可以阻止RNA和DNA病毒的復制,由于其具有廣譜效應,因此用于治療SARS。在臺灣的一項研究中,51名SARS患者在確診后每天服用氟喹諾酮類抗生素(左氧氟沙星500 mg或莫西沙星400 mg)。在51名患者中,44名SARS患者同時靜脈注射2 000 mg利巴韋林,每天口服1 200 mg,另外7名SARS患者沒有使用利巴韋林,必要時給予皮質激素、靜注甲基強的松龍或口服強的松龍治療惡化的肺浸潤和發熱。利巴韋林治療導致缺氧和貧血,反而增加了SARS患者的死亡風險[15]。在一項回顧性分析中,來自香港、新加坡和多倫多的229名患者接受了利巴韋林聯合皮質類固醇、免疫球蛋白或抗生素的治療[16],利巴韋林沒有顯示出療效。香港和新加坡的患者在診斷時口服1 200 mg利巴韋林,然后口服2 400 mg·d-1,或持續靜脈注射利巴韋林[8 mg·kg-1,每8 h一次]。在多倫多,患者首次靜脈給藥利巴韋林2 000 mg,后續每6 h 1 000 mg持續4 d,每8 h 300 mg持續3 d。不幸地是,利巴韋林治療組和對照組的死亡率相似。由于利巴韋林會導致貧血、低氧血癥和血紅蛋白水平下降等不良反應,并沒有改善患者的預后,甚至增加了SARS患者的死亡風險[17]。由于不良反應且療效不足,加拿大衛生部停止利巴韋林用于治療SARS[15]。
另一項研究是關于利巴韋林與抗逆轉錄病毒藥物洛匹那韋(lopinavir)聯合使用的療效。洛匹那韋已在體外被證明有抗SARS-CoV活性[18]。在一項非隨機、開放臨床試驗中,152名疑似SARS患者接受了的利巴韋林和糖皮質激素的治療。而另外41名確診的SARS患者采用洛匹那韋(400 mg)和利托那韋(100 mg)的聯合治療,治療組在d 10的拭鼻實驗的平均病毒負荷量降到檢測限以下。與歷史對照組相比,SARS相關癥狀消退,病情進展較輕,且無不良反應報告。
在另一項22名SARS患者的開放標記、非隨機臨床研究中,9名患者接受了皮下注射干擾素-1(IFN),初始劑量為9 lg·d-1,持續2 d,并根據病情進展逐漸增加劑量到15 lg·d-1,共10 d。9例患者全部存活且副反應較少[19]。
2.1.2MERS藥物治療 利巴韋林(含或不含干擾素/皮質類固醇)是MERS暴發期間的主要治療手段。在一項回顧性分析中,20名患者接受口服利巴韋林和皮下注射PEG-IFN-2α治療,劑量為180 lg/周,持續2周[20]。利巴韋林的初始劑量為2 000 mg,隨后根據肌酐清除率調整為200~1 200 mg,而24例接受支持性護理和皮質激素治療的患者為對照組。在MERS確診后14 d,治療組的存活率為70%,對照組為29%。診斷后28 d,治療組的存活率為30%,而對照組的存活率為17%[20]。在另一項病例研究中,一名69歲的希臘患者在吉達感染了MERS后,口服洛必那韋/利托那韋(400/100 mg,每日兩次)、PEG-IFN (180 lg 皮下注射,每周一次,持續12周)和利巴韋林(從診斷后d 13開始,2 000 mg初始劑量,后續1 200 mg每8 h,持續8 d)。治療開始兩天后,檢測不到病毒血癥。然而,在診斷后14周,在病人的糞便、呼吸道分泌物和血清中檢測到病毒RNA。盡管存活時間延長,但病人還是診斷后2個月死于膿血性休克[21]。
2.2 具有治療冠狀病毒感染潛力的藥物由于新爆發的傳染性疾病(如SARS和MERS),已有的治療方案往往不起作用,因此迫切需要新療法。傳統藥物研發由于開發周期非常長,往往無法滿足要求。而“老藥新用”可能為新型冠狀病毒的治療提供一個新的突破口。首先,如果這些藥物的安全性已經得到充分的驗證,只要在體外和動物研究中被證實具有治療作用,那它們可用于抗擊新型冠狀病毒的第一線。其次,這些藥物比較容易得到,可以相對快速地提供給具有緊急使用授權的患者。
齊多夫定最初是在1964年作為一種抗癌藥物被開發出來的,1985年,人類發現齊多夫定對人類免疫缺陷病毒(HIV)活性具有抑制作用。兩年后,它成為第一個被批準用于治療獲得性免疫缺陷綜合征(AIDS)的藥物[22]。
目前,關于已上市的藥物用于抗病毒感染(包括冠狀病毒)治療的研究已經有不少被報道。接下來,本文將總結幾類對MERS-CoV和SARS-CoV具有抗病毒活性的藥物,同時也包括一些對其他病毒家族有作用、廣譜的,并在臨床上用于其他病毒感染的藥物。需要強調的是,本文所述的治療方案僅限于學術研究討論,不推薦直接用于臨床。
2.2.1抗瘧藥 抗瘧藥氯喹、阿莫地喹、甲氟喹對SARS和MERS-CoV冠狀病毒都有效[23-25]。氯喹是美國FDA批準的抗瘧藥,也可用于治療自身免疫性疾病,如類風濕關節炎的抗炎[26]。氯喹在體外對多種病毒有抑制作用。在體內對包括登革熱病毒(DENV)、托加病毒(CHIKV)、尼帕病毒、流感病毒、艾滋病毒以及埃博拉病毒(EBOV)有效。氯喹的抗病毒作用包括非特異性地阻止病毒早期進入宿主細胞,或抑制病毒后期在宿主細胞復制。氯喹在酸性細胞器中(如核內體、高爾基囊泡和溶酶體)積聚。在這些細胞器中藥物質子化可導致囊泡pH增加。病毒依靠這些酸性細胞器進入宿主細胞、復制和后代成熟。MERS-CoV進入細胞依賴于蛋白酶(proteases)。二肽基肽酶4(DPP4)是一種功能性的病毒受體[27]。細胞蛋白酶激活病毒spike糖蛋白(如Ⅱ型跨膜絲氨酸蛋白酶(TMPRSS2)及cathepsin家族)[28]。氯喹可能對這些蛋白酶中的任何一種都有影響。氯喹還影響細胞高爾基體的糖基化,從而影響病毒蛋白運動和成熟。氯喹的抗病毒作用可使SARS-CoV的血管緊張素轉換酶2(ACE2)受體的糖基化缺陷[29]。
氯喹的廣譜抗病毒活性使它可以“老藥新用”的方式治療新型冠狀病毒和其它新發病毒感染。盡管氯喹的體內活性在MERS或SARS動物模型中還沒有報道。這種藥物抗其它病毒的活性報道的臨床前和臨床研究結果不一致。氯喹的血漿穩定濃度在小鼠和人約10 μmol·L-1,這一濃度落在其抗MERS-CoV(3.6 μmol·L-1)和抗SARS-CoV(2.3 μmol·L-1)的有效EC50值范圍內[30]。小鼠試驗證明氯喹可提高流感和埃博病毒感染的存活率。臨床研究表明,氯喹能有效地降低無癥狀HIV患者的病毒負荷[31]。但是氯喹治療CHIKV和DENV病毒感染的結果不相一致[32-33]。
氯喹的廣譜抗病毒的研究報道有助于抗MERS、SARS和COVID-19的治療。氯喹的可藥性(drugability)特點包括胃腸道吸收快、成本低(我國具有成熟的氯喹生產線)、良好的體內分布。氯喹可能是最佳的與其它抗病毒藥物聯合用藥的候選者。但是還需要增強對其抗病毒藥效學、血漿濃度和毒理學的研究[34]。羥基氯喹由于其與氯喹相似的藥理作用但毒性低,可以作為替代品。抗瘧藥阿莫地喹也有體外抗MERS-CoV和SARS-CoV的活性,其抑制絲狀病毒復制的機制和作用與氯喹相似[30]。阿莫地喹耐受性好,已經用于許多國家抗瘧治療。阿莫地喹聯合青蒿琥酯(artesunate)在2013-2016年EVD病毒流行期間顯著降低死亡率[35]。WHO已經報道了阿莫地喹用于非靈長類動物EVD模型的研究。此外,甲氟喹(mefloquine)可能也是一種具有抗MERS、SARS活性的奎寧衍生物[25]。
2.2.2干擾素 (interferons) 干擾素(IFNs)已FDA批準用于丙型肝炎等適應癥。盡管IFN-α能夠減緩小鼠和靈長類動物體內的SARS-CoV的復制過程[36-37],但IFN-α對SARS患者的治療效果不一(見2.1.1節)。從體外研究來看,另一種I型干擾素IFN-β1a可能比IFN-α單獨或聯合IFN-γ使用更有效[38-40]。在體外實驗中,IFN-β和IFN-γ聯合使用對抗SARS-CoV有協同作用[40]。
體內外的臨床前研究表明,如果在疾病早期給予MERS患者IFN-β2b單用或與利巴韋林聯合使用,可能產生治療效果[41, 42]。然而,在臨床試驗中,IFN-α2b(與其他治療聯合使用)并沒有產生療效 (見2.1.2節)。與IFN-β2a(EC50=160.8 IU·mL-1)、IFN-α2b(EC50=21.4 IU·mL-1)和IFN-c(EC50=56.5 IU·mL-1)相比,IFN-β1a (EC50=1.37 IU·mL-1)在體外對MERS-CoV的活性抑制作用更強。目前一項評估IFN-β1b(與洛品那韋/利托那韋聯合)對MERS-CoV的作用隨機臨床試驗正在進行。研究表明IFN-β亞型(1a和1b)與其他抗病毒藥物聯合具有療效,因為潛在的協同組合可以減少有效藥物劑量和IFN的不良反應[43]。
2.2.3激酶抑制劑 蛋白激酶參與了許多重要細胞通路的調節。Dyall等[25]通過細胞篩選實驗發現了兩種Abelson (Abl)激酶抑制劑(伊馬替尼和達沙替尼)可以抑制MERS-CoV和SARS-CoV感染。兩種化合物均能顯著抑制MERS-CoV和SARS-CoV活性,其EC50值為微摩爾,細胞毒性較低。其中,Abl2被認為是MERS-CoV和SARS-CoV病毒入侵的關鍵,這可能是伊馬替尼能抑制這兩種病毒入侵的原因[44]。最近一項關于MERS-CoV體外感染的系統激酶組學分析研究表明,ERK/MAPK和PI3K/Aκt/mTOR信號通路在病毒感染過程發揮了特殊作用[45]。隨后,針對這些信號通路,使用ERK/MAPK激酶抑制劑司美替尼(selumetinib)和曲美替尼(trametinib),發現在MERS-CoV感染前或感染后的抑制率可以達到95%以上[45]。此外,曲米替尼對MERS-CoV的抑制作用明顯強于塞魯米替尼,提示ERK/ MAPK信號通路的特異性中間體可能是病毒生命周期中早期(病毒進入)和晚期(病毒復制)事件的關鍵靶點。相比之下,索拉菲尼(Raf-1、B-Raf抑制劑)(ERK/MAPK信號通路的組成部分)的抑制劑,在體外細胞實驗中,如果在感染前添加,MERS-CoV的抑制率可達90%,但如果在感染后添加,其抑制率則降到30%以下,表明Raf激酶主要參與了病毒生命周期的早期事件。此外,感染后使用Raf激酶抑制劑達拉非尼(dabrafenib),其抑制活性也大大減弱。mTOR抑制劑西羅莫司(sirolimus)或埃維莫司(everolimus),在MERS-CoV感染前或感染后的抑制率都在60%左右,表明mTOR在MERS-CoV感染中起著作用。Wang等[46]最近的一項臨床研究,通過隨機對照試驗,評估了在標準抗病毒治療之中添加使用西羅莫司和糖皮質激素對重癥H1N1肺炎和急性呼吸衰竭患者的療效。發現西羅莫司可以改善患者的預后,包括減少缺氧和多器官功能障礙,提高病毒清除率。
2.2.4肉桂硫胺(cinanserin或cinnamamine) 1960年代,美國藥企Squibb合成了一種5-羥色胺抑制劑,即肉桂硫胺(代號SQ 10643;這種代號是美國國立衛生研究院NIH系統設置的,表明NIH參與了這一藥物的研發)。肉桂硫胺分子式C20H24N2OS,分子量340.482。動物試驗表明它有抗羥色胺作用、中樞神經系統作用、免疫抑制作用和消炎作用[47]。這種藥物早期作為免疫抑制劑用于狗的腎移植,可增加腎移植狗的生存率而沒有顯著骨髓抑制作用。對于腎移植狗,它的主要副作用是每天重復劑量超過40 mg·kg-1·d-1時產生的肝毒性。當靜脈輸注藥物(0.5 mg·kg-1·min-1)時,致死劑量為100 mg·kg-1[48]。對精神分裂癥、躁狂癥和類癌綜合征患者進行了小規模臨床試驗觀察到肉桂硫胺的良好效果,患者口服劑量是600~800 mg·d-1(10 mg·kg-1)。在短期治療用藥最大劑量為1 200 mg·d-1(20 mg·kg-1)[49]。雖然在人體沒有觀察到明顯的副作用,但進一步的臨床試驗被FDA暫停。因為在大鼠長期高劑量(120 mg·kg-1·d-1,持續59~81周)給藥后發現該藥可誘發肝癌[50]。上述數據表明,此藥短期、低劑量使用可能是安全的。
由歐盟的4家研究機構和中國的4家研究機構(包括中科院上海藥物所)參與的《中歐SARS診斷和抗病毒項目》課題組在我國2003年SARS后,提出肉桂硫胺的老藥新用策略[47]。他們將SARS-CoV的3C樣蛋白酶(3CLpro)的同型結構和晶體結構與肉桂硫胺進行分子對接,表明該藥對酶的活性區結合較好。表面等離子共振(SPR)技術證明肉桂硫胺與SARS-CoV和HCoV-229E均可結合。該藥的IC50值是5 μmol·L-1。在無毒性的濃度范圍內,該藥針對冠狀病毒RNA的復制和感染性病毒顆粒的IC50值分別是19和34 μmol·L-1。肉桂硫胺可能是一種有效的控制SARS的藥物。
2.2.5核酸合成抑制劑 多種RNA/DNA合成抑制劑對包括SARS-CoV和MERS-CoV等病毒具有廣譜活性。肌苷單磷酸脫氫酶(IMPDH)抑制劑,如利巴韋林、麥考酚酸和咪唑比等,可抑制核酸的從頭合成過程中的重要步驟。利巴韋林已與干擾素聯合應用于臨床,用于治療丙型肝炎等病毒感染。利巴韋林已用于SARS和MERS (見2.1),利巴韋林在體外對MERS-CoV的抑制作用較弱,而關于利巴韋林對SARS-CoV的的抑制作用的數據則有些矛盾[36, 38, 51]。
霉酚酸(MPA)是一種預防器官排斥反應的免疫抑制劑,具有廣譜抗病毒活性,并具有抗真菌、抗菌、抗癌和抗銀屑病的特性[52-54]。MPA在體外對SARS-CoV的抑制活性較弱,但對MERS-CoV有較好的抑制活性[51,55]。其前藥霉酚酸酯可改善口服生物利用度[56]。咪唑立賓(mizoribine)也是一種用于器官移植的免疫抑制劑,在體外已經顯示出對HCV和牛病毒性腹瀉病毒(BVDV)的活性,被作為利巴韋林/IFN聯合治療HCV感染的替代方案[57-58]。
化療藥物吉西他濱(gemcitabine)在體外具有抗MERS-CoV和SARS-CoV活性[25]。它通過抑制核糖核苷酸還原酶,從而抑制嘧啶的生物從頭合成,從而抑制病毒RNA轉錄和復制,并在小鼠白血病病毒模型中顯示了體內抗逆轉錄病毒活性[59-60]。
2.2.6蛋白酶抑制劑 MERS-CoV和SARS-CoV需要宿主的蛋白酶來激活其包膜糖蛋白,才能通過內體或非內體途徑進入細胞。因此,宿主細胞蛋白酶也是潛在的靶點,其抑制劑作為潛在的抗病毒藥物正在研究[61]。絲氨酸蛋白酶TMPRRSS2通過非內體途徑介導MERS-CoV 和SARS-CoV的進入[28, 62-64]。甲磺酸卡莫司他用于治療慢性胰腺炎,可抑制TMPSSR2介導的糖蛋白對MERS-CoV、SARS-CoV和流感病毒的激活[61, 65-67]。半胱氨酸蛋白酶抑制劑K11777目前正處于治療寄生蟲感染的臨床開發階段。K11777對冠狀病毒(MERS-CoV、SARS-CoV、HCoV-229E)、絲狀病毒(EBOV、Marburg病毒)和副粘病毒(Nipah病毒)具有廣譜活性[68]。Zhou等證明了卡莫司他和K11777對SARS-CoV有抑制活性,而EBOV僅被K11777抑制,這表明這些病毒所需的宿主蛋白酶不同[68]。E-64-D是一種內泌體半胱氨酸蛋白酶的抑制劑,目前正處于治療肌營養不良癥的Ⅲ期臨床試驗中。E-64-D也能抑制絲狀病毒細胞進入[69-70]。由于病毒依賴于特定的宿主絲氨酸或半胱氨酸宿主蛋白酶,因此需要進一步了解宿主蛋白酶與病毒發病機制之間的關系,從而選擇合適蛋白酶抑制劑,確定最有效的治療方案。
洛吡那韋(lopinavir)是HIV蛋白酶的抑制劑,臨床上用于治療HIV感染[71]。但其在體外也具有抑制MERS-CoV和SARS-CoV,添加時間實驗表明該藥物在病毒進入的早期階段起作用[23]。因為洛必那韋的生物利用度很低,它常與細胞色素P450 3A4抑制劑利托那韋聯合使用,以增加血液濃度[71]。洛吡那韋/利托那韋治療可降低MERS 非人靈長類模型的死亡率。洛吡那韋可以靶向SARS-CoV的主要蛋白酶(Mpro)[72],但它也可以作用于冠狀病毒在細胞內的其他復制過程,因此需要更多的研究來充分理解洛必那韋的作用機制。在2003年SARS暴發期間,在公開臨床試驗中使用洛吡那韋/利托那韋聯合利巴韋林治療的患者病程較輕,致死率較歷史對照組有所降低[17-18]。
2.2.7蛋白質合成抑制劑 蛋白合成抑制藥能夠靶向抑制蛋白質合成,從而抑制冠狀病毒,通過與藥物聯合治療,將具有抗冠狀病毒的潛力。依米汀(emetine)是一種天然的植物生物堿,茴香霉素是一種抗生素,都具有抑制蛋白質的伸長的作用,被用于抗原生動物[73-74]。此外,依米汀的合成衍生物去氫依米汀(dehydroemetine),其副作用較少,目前也正在藥物開發階段[75]。茴香霉素(anisomycin)最初作為肽基轉移酶抑制劑在使用,被發現可以激活MAP激酶信號通路[76]。除了具有抑制MERS-CoV和SARS-CoV的活性外,埃米汀和茴香霉素也具有抑制動物小核糖核酸病毒腦心肌炎病毒的活性[77]。茴香霉素在體外有抗脊髓灰質炎病毒活性[78]。高三尖杉酯堿(omacetaxine mepesuccinate)是一種植物性生物堿,是一種抗癌藥物,于2012年獲得FDA批準用于治療慢性髓系白血病。高三尖杉酯堿可抑制MERS-CoV、牛冠狀病毒、人腸道冠狀病毒和乙型肝炎病毒[79-80]。
2.2.8血管緊張素轉換酶(ACE)系統藥物對肺冠狀病毒感染的最新研究 腎素-血管緊張素系統(renin-angiotensin system,簡稱RAS)由血管緊張素原、血管緊張素轉化酶(ACE)及其同源血管緊張素轉換酶2(ACE2)和血管緊張素Ⅱ(Ang Ⅱ)1型和2型受體(AT1,AT2)組成。ACE將10肽Ang I轉變為8肽Ang Ⅱ;而ACE2將Ang Ⅱ切除一個殘基生成Ang-(1-7),繼而阻斷Ang Ⅱ并抑制ACE[81, 82]。從而,ACE2軸負調節ACE活性(Fig 1)。
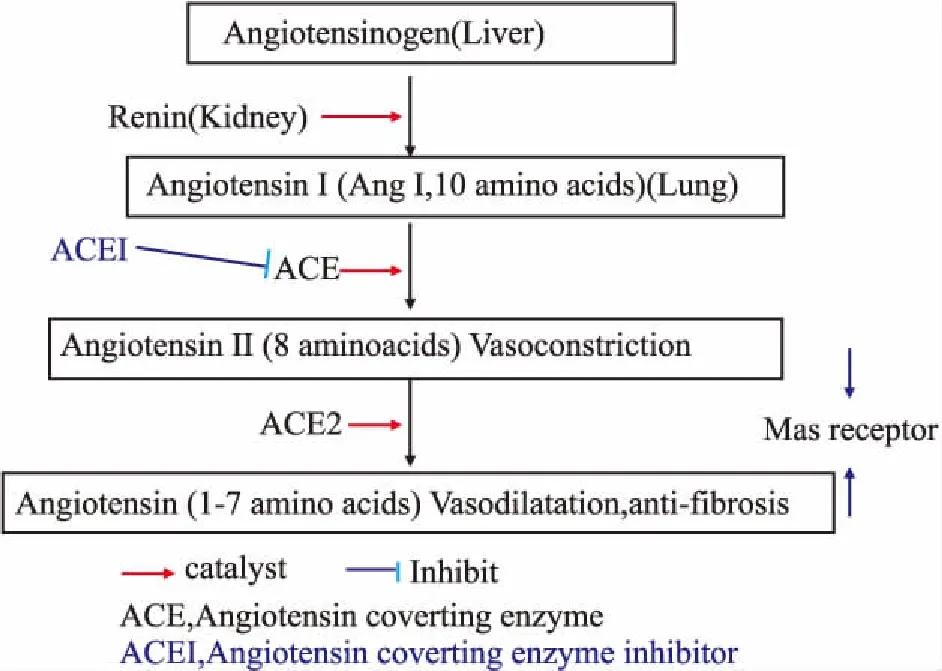
Fig 1 Relationship between ACE, angiotensins and ACEI
以前人們主要關注RAS在血壓穩態和心血管疾病中的作用[83],現在也開始重視RAS在肺病理生理中的作用。肺毛細血管內皮細胞表達20%的ACE[84]。因此,肺血管中大量的ACE可將Ang I轉換為Ang Ⅱ,引起血管收縮、血流量降低,導致通氣/灌流不匹配和組織缺氧。 ACE2主要在Clara 細胞和Ⅱ型肺泡上皮細胞產生。在急性呼吸窘迫綜合征(ARDS)和/或呼吸機誘發的肺損傷(VILI)時,生成ACE2的能力受損,導致ACE活性占主導地位[85-86]。活性氧(ROS)、NADPH氧化酶和過氧化氫(H2O2)是RAS和ACE的上游調節劑[7]。RAS誘導細胞內活性氧的產生,進而調節細胞內第二信使及其下游信號。在正常情況下ROS和RAS之間的相互作用維持肺功能的完整性。在ARDS和VILI條件下,ROS和RAS的相互作用失常,導致肺部損傷[85, 87]。
ACE系統與呼吸系統的相互作用很早就闡明了。主要是ACE-AngⅡ軸的可引起肺毛細血管收縮,降低氣血交換。Ferrario等[88]報道了ACEI和AngⅡ受體抑制劑對心臟ACE2酶的作用,指出ACE抑制的綜合結果是減少AngⅡ的形成和延緩Ang(l-7)的代謝。ACE2本身也可增加AngⅡ的代謝。Imai等[89]2005年報告ACE2和血管緊張素Ⅱ2型受體(AT2)對小鼠嚴重急性肺損傷有保護作用,而ACE、血管緊張素Ⅱ和血管緊張素Ⅱ1a型受體(AT1a)則誘發肺水腫、損害肺功能;缺乏ACE的小鼠表現出明顯的疾病改善,并且重組ACE2單抗對小鼠嚴重急肺損傷有保護作用。ACE2受體與SARS-CoV的聯系因此引起注意。
從5個新型冠狀病毒肺炎暴發初期病人支氣管肺泡灌洗液中分離出來的病毒全基因組序列與SARS-Cov全基因組序列79.5%相同。新型冠狀病毒與SARS-CoV共用ACE2受體進入上皮細胞[4, 12]。冠狀病毒表面S蛋白影響病毒對宿主的感染與傳播。S1亞基結合細胞表面受體,S2亞基幫助病毒進入細胞。SARS-CoV spike糖蛋白的冷凍電鏡結構分析表明,上調S1的C端結構域1(CTD1s)有利于病毒結合[90]。人冠狀病毒NL63(HCoV-NL63)感染細胞的3D模型表明[91],感染的第一步是通過硫酸乙酰肝素蛋白多糖(或稱HS proteoglycan;mucin)與靶細胞(如LLC-Mk2細胞系)的ACE2受體蛋白相互作用,觸發網格蛋白(clathrin),隨后囊泡被動力蛋白(dynamin)切斷,導致病毒內吞,形成囊泡(vesicle)。后者通過肌動蛋白皮質,完成細胞骨架重組。最后,內體微環境(endosomal microenvironment)的酸化使病毒基因組成功融合并釋放到細胞質中。由于發現阻止SARS表面糖蛋白與ACE2的相互作用可能抑制SARS進入宿主細胞,Adedeji等人提出了3種制止SARS-CoV疾病的策略并篩選了相應的3種抑制劑: 1)抑制早期SARS-S-ACE2相互作用;2)抑制clathrin L活性; 3)阻止融合[28]。
目前尚難斷定ACEI抑制劑在COVID-19和SARS中的利弊,因為沒有直接的臨床試驗數據可供參考。我們也不知道ACEI是否可以與ACE2結合從而阻止病毒進入細胞,但至少ACEI可舒張肺毛細血管。外源性Ang-(1-7)皮下或注射給藥對實驗性腎大部切除大鼠可誘發心臟副作用包括高血壓、心肌肥厚和纖維化。ACEI藥物雷米普利(ramipril)可對抗這種副作用[92]。ACE活性增加或ACE2活性下降可能會促進肺損傷[93]。重組人ACE2抗體(rhACE2;GSK2586881)的臨床Ⅱ期結果表明rhACE2是安全的。每天兩次輸注可導致血漿Ang Ⅱ快速下降,Ang-(1-7)和Ang-(1-5)水平增加,以及血漿IL-6濃度下降[94]。卡托普利可促進ACE2-Ang(1-7)-Mas軸的表達,抑制ACE1、AngⅡ和AT1受體[95]。
2.3 尚在開發中的藥物
2.3.1抑制MERS-CoV和SARS-CoV的潛在靶點 除了老藥新用外,還需要研發新的抗病毒藥物或育苗來對付新冠狀病毒,新設計藥物通常靶向病毒生命周期、宿主上相關通路的重要靶標蛋白。瑞德西韋(remdesivir、GS-5734)和BCX4430等兩種核苷抑制劑有可能成為廣譜抗病毒藥物,它們作用于病毒RNA所依賴的RNA聚合酶[96-97]。這兩種藥物在細胞培養中對MERS-CoV和SARS-CoV均有抑制作用。特別是瑞德西韋,在美國首例VODID-19確診病例中有優異的臨床表現,目前已經通過綠色通道,在我國快速進入Ⅲ期臨床試驗。
SARS-CoV和MERS-CoV等冠狀病毒的表面糖蛋白(S)由兩個結構域組成,S1包含細胞外結合所需的受體結合域(RBD),S2含有膜融合釋放所需的融合肽。SARS-CoV的內吞作用是通過RBD與宿主細胞上的血管緊張素轉換酶2 (ACE2)受體結合而實現的。膜上的組織蛋白酶L裂解S蛋白,使S2融合蛋白暴露,S2再與膜融合并釋放病毒RNA。因此,組織蛋白酶L、ACE2-SARS-S1復合物、S2融合肽都可能是抑制SARS-CoV進入的合適靶點[98]。最近的研究發現,病毒進入抑制劑、病毒蛋白酶和解旋酶均可有效抑制MERS-CoV和SARS-CoV[98]。宿主細胞上的蛋白酶,如木瓜蛋白酶和3C樣蛋白酶,也可以作為潛在的抗病毒靶點,因為它們會裂解非結構蛋白,而這正是病毒成熟所需要的步驟。現有的大多數蛋白酶抑制劑都屬于“自殺”式蛋白酶抑制劑,即不可逆地結合到靶標上。而如果可以設計成可逆的蛋白酶抑制劑,將可能具有更大的潛力,因為毒性更小,病人耐受性更好些[99]。此外,近年來對解旋酶抑制劑的研究表明,N端金屬結合域、鉸鏈域和NTP/解旋酶域這三個域靶點具有開發新藥的潛力[100]。
2.3.2RNA干擾 定向RNA干擾(RNAi)技術可以通過基因沉默,從而對新型病毒進行特異性治療[101]。最近的研究表明,小干擾RNA (siRNA)或短發夾RNA可以抑制病毒基因的表達,從而阻斷培養細胞中SARS-CoV的復制[102-103]。小干擾RNA 藥物siSC2-5,在經鼻給藥后,可靶向SARS-CoV開放閱讀框1和S蛋白,從而降低非人靈長類的SARS發病[173]。目前已有幾種用于治療病毒感染的RNAi療法進入了臨床試驗,包括TKM-Ebola,它是一種治療EBOV的siRNA/脂質納米顆粒[101, 104-105]。由于病毒基因組測序目前可以在很短的時間內完成,這項技術在新發現病毒的治療上將有很大的潛力,特別是在新型病毒暴發流行的情況下。RNA干擾目前的主要障礙在于如何確定合適的病毒靶標,以及解決藥物在體內對靶細胞的傳遞效率。
2.3.3肽類病毒進入抑制劑 多肽具有陽離子兩親性質,使其容易進入細胞膜,并可以在病毒進入時,對細胞與病毒膜的融合進行干擾。針對包膜病毒的肽類病毒進入抑制劑的研發目前已經取得了一些進展[106],如艾滋病毒抑制劑恩福韋肽(enfuvirtide)就是從HIV gp41的兩親環/c-螺旋七肽重復域中提取的一種含有36個殘基肽。通過尋找融合蛋白序列的兩親性序列,并進行合理設計,已經成為開發病毒(包括MERS-CoV和SARS-CoV)的肽抑制劑的有效手段[107-109]。此外,可以通過化學修飾,提高多肽藥物在體內的穩定性和生物利用度,提高其在臨床應用潛力[110-112]。
3 觀點與展望
高特異性和廣譜抗病毒藥物都各有各的優勢,抗病毒藥物可以在“廣譜”和“高度特異性”之間靈活選擇[113]。高特異性的藥物,針對特定病毒或病毒家族,雖然活性譜較窄,但具有較高的藥效和較低的毒性,不過這種藥物也容易產生耐藥[113]。與之相反,廣譜抗病毒藥物通常靶向宿主因素或信號通路,作用的范圍廣,比較不容易產生耐藥,但藥物的毒性往往較大、活性較低和療效較慢。
我們關于抗病毒藥物開發的大部分經驗來自對慢性病毒感染的研究。雖然針對DNA病毒的抗病毒開發策略已經成功地開發出一系列專門針對病毒蛋白的藥物,但這種策略在RNA病毒上并不太成功,因為RNA病毒比DNA病毒的變異率更高,從而更容易產生耐藥性。
3.1 聯合用藥為了達到更好的抗病毒效果,往往采用的兩種或兩種以上藥物同時或先后應用,從而增加藥物的療效、為了減輕藥物的毒副作用、并可以避免病毒出現耐藥性。藥物的聯合使用可以是廣譜抗病毒藥與特定抗病毒藥、作用機理不同的藥物、或針對病毒生命周期不同步驟的藥物。將一種或多種有效的活性組合,將有可能大大提高對新型冠狀病毒暴發的治療效果。但是聯合用藥有時也可能產生相反的結果。所以合理的聯合用藥,應以提高療效和(或)降低不良反應為基本原則。聯合用藥時,藥物的相互作用,應包括影響藥動學的相互作用,應包括影響藥效學的相互作用。已經有人提出建立一個包含藥物動力學數據和傳染病生物學數據的數據庫,這將能夠更好地指導藥物的聯合使用[114]。
許多現有藥物具有廣譜抗病毒活性的潛力,并且已經在臨床上用于治療其他病毒感染。此外,進入臨床試驗階段的藥物也可考慮用于聯合治療。但應當對每種組合進行謹慎的藥理學評估,以避免藥物不良反應及疾病方面的禁忌癥。新型廣譜復制抑制劑,如瑞德西韋 (Gilead Sciences),免疫調節劑(如硝唑胺;類固醇;他汀類藥物和正在開發的針對冠狀病毒的直接作用抗病毒藥物是聯合用藥方案。
3.2 基于結構的藥物設計病毒蛋白晶體結構的解析為合理設計藥物提供了新的途徑。基于蛋白質結構信息進行合理藥物設計研究,通過計算機虛擬篩選的方法可以快速篩選出能與病毒靶標結合,具有活性的化合物。HIV-1蛋白酶抑制劑的設計是最成功例子之一。在HIV-1蛋白酶結構發表后僅6年,沙奎那韋就創紀錄地從實驗室走上臨床,并于1995年被批準用于治療艾滋病[115-117]。從1995年到2000年,有6種通過合理設計針對HIV-1的抗病毒藥物被批準上市。此外,基于已知病毒蛋白晶體結構的計算機輔助方法還成功地指導了多個HCV NS3/NS4A蛋白酶抑制劑的設計和合成,如肽酶、特拉匹韋和博西潑韋[118-120]。
目前SARS-CoV和MERS-CoV病毒蛋白酶Mpro和PLpro的晶體結構被解析出來[121]。這些結構已經被用于設計和開發對蛋白酶活性位點具有高結合親和力的抑制劑。這些結構的確定可以為新型冠狀病毒肺炎的藥物開發提供新的病毒靶點。
4 結論
當前新冠肺炎來勢洶洶,盡管政府采取了諸多強有力的防控措施,有效遏制病情的快速發展,但是病毒還是在一個多月感染了6萬多人,死亡上千人。目前針對COVID-19的研究剛剛起步,臨床上極度缺乏針對性治療藥物,迫切需要尋找和開發新的治療方法和藥物。而開發新的針對性藥物及病毒疫苗需要大量的時間和費用,無法滿足疫情控制和臨床上的迫切需求。而“老藥新用”則可能提供一個新的突破口,在已上市藥物中篩選出能夠抑制病毒活性的藥物,將是一個更好的選擇。由于已上市藥物的安全性已經得到充分的驗證,只要在體外和動物研究中被證實具有治療作用,就可以快速用于抗擊疫情的第一線。此外,這些藥物比較容易獲得,可以相對快速地提供給具有緊急使用的患者。本文總結多種對MERS-CoV和SARS-CoV具有抗病毒活性的已上市藥物,包括一些對其他病毒有作用、相對廣譜藥物,希望能夠對當前抗擊疫情有所幫助。
我們成功戰勝了2003的SARS,今年的新冠肺炎我們也將必勝,但是病毒也在快速變異,將來還可能有新的冠狀病毒卷土重來,使已有的診療方案失效。因此,在尋找病毒特效藥的同時,也需要注意已有廣譜藥物抗病毒的收集(如中藥或已有藥物),和新廣譜藥物研發。在面對新病毒和疫情時,可以從一系列備選藥物中快速找到抗病毒的診療方案,并采用聯合用藥的方法,以提高藥效,降低不良反應,并減少耐藥性,從而能夠面對各種新舊病毒的挑戰。