載人航天工程中的微生物科學(xué)與技術(shù)應(yīng)用
2020-05-06 00:57:48袁俊霞馬玲玲徐侃彥
載人航天 2020年2期
袁俊霞,印 紅,馬玲玲,徐侃彥
(航天神舟生物科技集團(tuán)有限公司空間微生物研究室,北京100190)
1 引言
微生物是地球上出現(xiàn)最早、分布最廣、多樣性最豐富的生物類群。 在地球漫長(zhǎng)的演化歷史中,微生物因其生長(zhǎng)周期短、繁殖快、變異累積豐富等遺傳特征,形成了極高的生物多樣性,同時(shí)對(duì)多種極端環(huán)境產(chǎn)生了極強(qiáng)的環(huán)境耐受性。 在空間極端環(huán)境下一些微生物被發(fā)現(xiàn)可以生存且表現(xiàn)出良好的環(huán)境適應(yīng)性[1-2]。 考慮到空間環(huán)境的獨(dú)特性與復(fù)雜性,探索微生物在空間環(huán)境中的生存能力與適應(yīng)機(jī)制成為推進(jìn)人類空間探索可持續(xù)發(fā)展、支撐人類開展地外生命探索和宇宙生命起源等基礎(chǔ)科學(xué)研究的核心問(wèn)題。 同時(shí),在載人航天活動(dòng)支撐下,利用微生物在空間環(huán)境下特有的生命機(jī)能、活動(dòng)特性和代謝過(guò)程,發(fā)展服務(wù)于空間和地面環(huán)境的微生物技術(shù)和轉(zhuǎn)化應(yīng)用,將大大豐富地面醫(yī)藥、環(huán)境、能源和農(nóng)業(yè)等領(lǐng)域的發(fā)展。
2 空間環(huán)境特點(diǎn)
空間環(huán)境是由空間高能粒子輻射、微重力、高真空、溫度驟變等復(fù)雜因素組成的獨(dú)特環(huán)境。 隨著載人航天任務(wù)的推進(jìn),人類對(duì)空間環(huán)境的探索已經(jīng)從近地軌道環(huán)境推向深空環(huán)境,兩者的環(huán)境參數(shù)差異如表1 所示。 與近地軌道相比,處于深空環(huán)境中的生命體受環(huán)境影響表現(xiàn)往往更加極端[3]。
空間高能粒子輻射是制約人類開展長(zhǎng)期深空探索的關(guān)鍵因素,主要由各種能量以及隨時(shí)空變化的粒子組成。 對(duì)于近地軌道任務(wù),主要的輻射來(lái)源是銀河宇宙輻射和地磁捕獲粒子(范艾倫帶),輻射粒子主要由質(zhì)子和電子組成[4]。 在深空任務(wù)中,由于脫離了地磁保護(hù),主要的輻射來(lái)源是銀河宇宙輻射和太陽(yáng)粒子事件。 單次火星任務(wù)(單程飛行各6 個(gè)月,表面駐留2 年)航天員受到的累積輻射總量接近1 Sv[5]。 對(duì)微生物而言,高能粒子輻射能量高、貫穿力強(qiáng),可以直接作用于DNA,DNA 雙鏈斷裂是高能粒子輻射造成的最嚴(yán)重?fù)p傷[5-6]。
空間電磁輻射也會(huì)對(duì)進(jìn)入其中的生命體產(chǎn)生影響。 由于沒(méi)有地球大氣臭氧層對(duì)290 nm 以下射線的阻擋,在近地軌道存在全光譜紫外輻射,除了UVA(320~400 nm)、UVB(290 ~320 nm)長(zhǎng)波紫外輻射,還包含被大氣層屏蔽的UVC(190 ~280 nm)和真空UV(<190 nm),其中,UVC 波長(zhǎng)短、能量高,對(duì)地球生命體有強(qiáng)烈的刺激作用[3,6]。
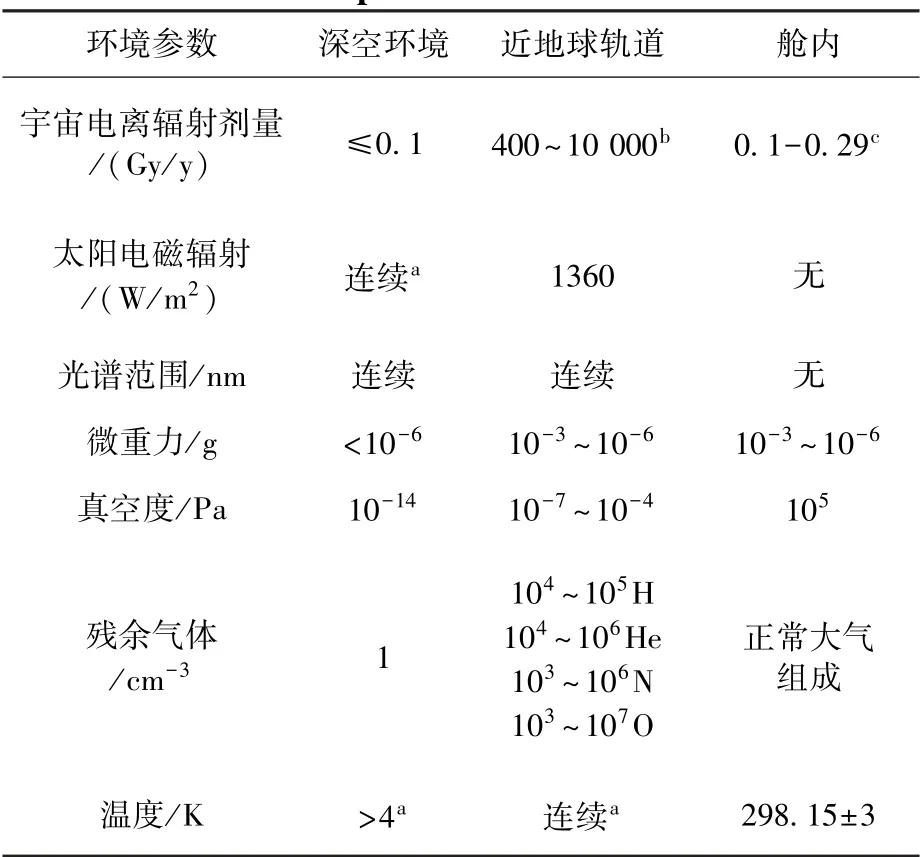
表1 深空環(huán)境、LEO 和空間站艙內(nèi)的環(huán)境參數(shù)比較[3]Table 1 Comparison of environmental parameters in interplanetary space, low Earth orbit, and cabin of manned spacecraft[3]
微重力環(huán)境中自然對(duì)流、沉積等現(xiàn)象的消失,將誘使生命體產(chǎn)生不同于重力環(huán)境的生命特征。近地軌道飛行過(guò)程中微重力介于10-3~10-6g。 此外,不同軌道飛行任務(wù)中壓力和溫度變化也不同。在近地軌道任務(wù)中,真空度介于10-7~10-4Pa,火星上的大氣壓力約為700 Pa,比地球上低100 多倍。 在星際空間,真空度可達(dá)10-14Pa。 空間站外直接暴露在太陽(yáng)下的溫度在-120 ~120 ℃之間波動(dòng)。 在火星任務(wù)中,探測(cè)器數(shù)據(jù)顯示溫度在-153 ℃(極點(diǎn))~20 ℃間波動(dòng)[4]。
3 空間環(huán)境對(duì)微生物的影響
大量的空間實(shí)驗(yàn)表明,在空間環(huán)境下生長(zhǎng)的細(xì)菌易表現(xiàn)出許多不同于地面的生物學(xué)特征,如生長(zhǎng)速度加快、遲緩期縮短、毒力增加、抗生素抗性增強(qiáng)、更易形成生物膜等,并且隨著空間飛行時(shí)間延長(zhǎng),這些生物效應(yīng)明顯加劇[7-8]。
微重力環(huán)境是影響微生物空間行為的主要因素。 與地球重力環(huán)境不同,微重力條件下對(duì)流和沉積現(xiàn)象的消失,會(huì)導(dǎo)致微生物細(xì)胞在生長(zhǎng)、代謝過(guò)程中與環(huán)境中的物質(zhì)交換效率降低,進(jìn)而使微生物對(duì)營(yíng)養(yǎng)物質(zhì)的有效獲取難度增大。 Zea[9]對(duì)大腸桿菌的研究進(jìn)一步支持了上述觀點(diǎn):微重力環(huán)境會(huì)激活大腸桿菌體內(nèi)能量合成相關(guān)基因,誘發(fā)饑餓相關(guān)基因過(guò)表達(dá),加快細(xì)菌的新陳代謝。同時(shí),在空間條件下微生物趨向于聚集成更大的細(xì)胞群落,與生物膜相關(guān)的細(xì)胞外基質(zhì)形成加速,進(jìn)而通過(guò)形成生物膜增加對(duì)環(huán)境的耐受性,增加其在空間條件下的存活幾率。 空間環(huán)境下微生物生物膜的形成通常伴隨著毒性、感染性和抗生素抗性等特征的改變[10-12]。 此外,在空間條件下,微重力與空間輻射環(huán)境的互作可能會(huì)對(duì)微生物產(chǎn)生疊加效應(yīng),微重力環(huán)境影響DNA 損傷修復(fù)系統(tǒng)的正常功能也是導(dǎo)致微生物在空間環(huán)境下生物學(xué)行為變化的因素之一[6,13]。
研究表明,空間環(huán)境對(duì)微生物的影響涉及基因、生物組件、細(xì)胞功能和代謝產(chǎn)物等多個(gè)層面。Kim 等[12]基于微陣列和蛋白質(zhì)組學(xué)分析顯示,經(jīng)過(guò)空間培養(yǎng)的腸沙門氏菌體內(nèi)大量轉(zhuǎn)錄物和蛋白質(zhì)的表達(dá)會(huì)發(fā)生變化,其中,培養(yǎng)基中的無(wú)機(jī)磷酸鹽變化是引起其生物膜形成,毒性增強(qiáng)的重要因素[14]。 多項(xiàng)研究顯示,細(xì)菌RNA 分子伴侶蛋白Hfq 被認(rèn)為是調(diào)控上述反應(yīng)的關(guān)鍵因子,Hfq 調(diào)控系統(tǒng)也是發(fā)現(xiàn)的第1 個(gè)影響微生物空間環(huán)境適應(yīng)性的潛在作用機(jī)制[15-16]。 Hfq 作為細(xì)菌轉(zhuǎn)錄后調(diào)控網(wǎng)絡(luò)中的核心成員,通過(guò)結(jié)合sRNA 促進(jìn)其與目標(biāo)mRNA 配對(duì),從而觸發(fā)蛋白質(zhì)水平的調(diào)控。在細(xì)菌中,Hfp 缺失突變株表現(xiàn)出復(fù)雜表型,包括致病性、毒性減弱,環(huán)境適應(yīng)能力差等[17]。 此外,Wilson 等[18]研究表明,rpoS 編碼的穩(wěn)定σ 因子也是細(xì)菌應(yīng)激響應(yīng)的核心調(diào)控因子,被發(fā)現(xiàn)參與調(diào)控鼠傷寒沙門氏菌中的各種應(yīng)激反應(yīng)。 逐步挖掘與空間適應(yīng)性相關(guān)的潛在調(diào)控因子,揭示其與宿主細(xì)胞的相互作用機(jī)制,并將上述機(jī)制與微生物生物膜形成、毒性改變和適應(yīng)性變化等生物學(xué)特征相關(guān)聯(lián)是未來(lái)空間微生物研究的重點(diǎn)內(nèi)容。
4 微生物在載人航天工程中的應(yīng)用
4.1 空間微生物安全
在載人航天活動(dòng)中,俄羅斯早期的禮炮系列、和平號(hào)以及目前正在運(yùn)行的國(guó)際空間站中都發(fā)現(xiàn)了多種微生物生存和繁衍。 空間密閉艙內(nèi)微生物的過(guò)度積累以及輻射、微重力等空間因素將導(dǎo)致微生物生長(zhǎng)、致病性和毒性改變,并可能進(jìn)一步引發(fā)航天員出現(xiàn)感染、過(guò)敏等癥狀,及空氣和水源污染,各種結(jié)構(gòu)材料降解等。 因此,對(duì)空間站內(nèi)微生物生態(tài)長(zhǎng)期追蹤分析,確保微生物安全是支撐長(zhǎng)期載人航天活動(dòng)的重要內(nèi)容。
以國(guó)際空間站為例,科學(xué)家針對(duì)微生物安全的研究策略主要是:確定微生物在艙內(nèi)環(huán)境定殖的特征;分離和識(shí)別微生物的組成;確定抑制微生物活動(dòng)的有效方法[19-20]。 在空間站密閉艙內(nèi),多種生物組成生物膜是微生物在環(huán)境表面定殖的主要形式,這種生命形式可以增加微生物對(duì)環(huán)境的耐受性,其存在機(jī)制包括了微生物間合作、競(jìng)爭(zhēng)、信號(hào)分子傳遞以及水平基因轉(zhuǎn)移等[21]。 在微重力環(huán)境下,生物膜的形成具有與地面環(huán)境中不同的生物特征。 Kim 等[12]通過(guò)“Micro-2”實(shí)驗(yàn)研究微重力對(duì)銅綠假單胞菌、金黃色葡萄球菌生物膜細(xì)胞聚集的影響,發(fā)現(xiàn)在空間寡養(yǎng)條件下生長(zhǎng)的生物膜具有比在地球上更高的細(xì)胞密度,生物膜的厚度明顯增加。 同時(shí),銅綠假單胞菌在微重力環(huán)境下還有柱狀和冠狀生物膜的形成,鞭毛結(jié)構(gòu)在這種結(jié)構(gòu)的形成中發(fā)揮了關(guān)鍵作用。 當(dāng)前,由于空間實(shí)驗(yàn)條件的限制,針對(duì)微重力如何改變生物膜(微生物聚集)形成,以及微重力影響微生物腐蝕性等機(jī)理研究仍不清楚,開展上述研究對(duì)于制定新的微生物安全控制策略,最大限度地減少微生物對(duì)航天器的有害影響具有重要意義。
在國(guó)際空間站運(yùn)行過(guò)程中,多項(xiàng)空間微生物采樣和監(jiān)測(cè)項(xiàng)目用于分離和識(shí)別艙內(nèi)微生物的組成,并對(duì)后期微生物的動(dòng)態(tài)變化規(guī)律進(jìn)行預(yù)測(cè)。早期研究主要通過(guò)對(duì)不同環(huán)境介質(zhì)采樣、培養(yǎng)、觀察、統(tǒng)計(jì)進(jìn)行。 近年來(lái),基于PCR、芯片以及高通量測(cè)序的方法,發(fā)現(xiàn)了多種非培養(yǎng)法獲得的條件致病菌,進(jìn)一步完善了空間站微生物檢測(cè)結(jié)果[22-23]。 微生物在軌觀察項(xiàng)目“ISS-MO”將微生物群落變化、基因組變化與艙內(nèi)環(huán)境特征相關(guān)聯(lián),對(duì)空間站艙內(nèi)微生物展開了長(zhǎng)期多代的動(dòng)態(tài)分析。 對(duì)分離的20 種細(xì)菌比較發(fā)現(xiàn),長(zhǎng)期生長(zhǎng)的細(xì)菌對(duì)多種抗生素的抗性明顯增加[24];分離出的兩株煙曲霉的化學(xué)抗性雖然與地面相比無(wú)明顯變化,但其對(duì)斑馬魚幼蟲感染的毒性更加致命[25-26]。 該結(jié)果進(jìn)一步預(yù)示了微生物群落特征變化對(duì)航天員免疫系統(tǒng)的潛在風(fēng)險(xiǎn)。
基于此,美歐科學(xué)家正在對(duì)分離自空間站的微生物菌株遺傳信息等進(jìn)行收集分析,建立了相應(yīng)的空間微生物數(shù)據(jù)庫(kù)。 將微生物系統(tǒng)信息分析與特異微生物基因組分析方法相結(jié)合,對(duì)潛在的病原生物、腐蝕性微生物等進(jìn)行識(shí)別鑒定,進(jìn)而對(duì)某些微生物的生物安全風(fēng)險(xiǎn)進(jìn)行評(píng)估。 隨著我國(guó)空間站在軌建造任務(wù)臨近,首個(gè)航天微生物數(shù)據(jù)庫(kù)也正在構(gòu)建,數(shù)據(jù)庫(kù)囊括了我國(guó)空間站、火星探測(cè)器等航天器在建造過(guò)程中的環(huán)境微生物菌株[27-28],這些菌株將構(gòu)成未來(lái)我國(guó)空間站運(yùn)營(yíng)過(guò)程中微生物安全監(jiān)測(cè)和風(fēng)險(xiǎn)評(píng)估的“基線”。
4.2 微生物在生物再生生命保障系統(tǒng)中的應(yīng)用
利用天然、工程微生物菌群進(jìn)行艙內(nèi)航天員生活代謝產(chǎn)生的廢物轉(zhuǎn)化同時(shí)完成生物質(zhì)(食物)生產(chǎn),實(shí)現(xiàn)人類所需的營(yíng)養(yǎng)、氧氣和水等重要資源的再生是空間環(huán)境下生物再生生命保障系統(tǒng)中微生物功能部件開發(fā)的主要思路。 值得注意的是,微生物功能部件在空間環(huán)境下的穩(wěn)定性也是評(píng)估系統(tǒng)整體性能的重要因素。 因此,微生物,特別是針對(duì)特定目標(biāo)進(jìn)行遺傳改造的微生物工程菌在近地軌道環(huán)境以及深空環(huán)境的中儲(chǔ)存、運(yùn)輸以及復(fù)蘇后的遺傳穩(wěn)定性、生物活性、毒性等特征均需經(jīng)過(guò)空間驗(yàn)證與評(píng)估。
Lasseur 等[29]建造的微生態(tài)生命支持系統(tǒng)替代系統(tǒng)MELiSSA(Micro-Ecological Life Support System Alternative)是由微生物與植物共同組成的閉合系統(tǒng),微生物生物部件作為生產(chǎn)者與分解者均發(fā)揮了重要作用。 系統(tǒng)主要包含由嗜熱厭氧細(xì)菌組成的廢物分解系統(tǒng),由光合異養(yǎng)細(xì)菌、微藻(螺旋藻)、高等植物組成的食物生產(chǎn)系統(tǒng),以及由亞硝化細(xì)菌和硝化細(xì)菌組成的硝化室(將循環(huán)中產(chǎn)生的氨轉(zhuǎn)化為硝酸鹽,供光合細(xì)菌使用)。光合細(xì)菌由于具有營(yíng)養(yǎng)價(jià)值高、易消化、生長(zhǎng)速度快、收獲及后處理簡(jiǎn)單等優(yōu)點(diǎn),是系統(tǒng)中食物生產(chǎn)部件的良好選材。 在MELiSSA 系統(tǒng)大量地面研究的基礎(chǔ)上,節(jié)旋藻(Arthrospira)作為氧氣和生物質(zhì)生產(chǎn)的候選者,驗(yàn)證了其在空間站長(zhǎng)周期培養(yǎng)過(guò)程中生物質(zhì)生產(chǎn)動(dòng)力學(xué)和光合作用效率。 在此基礎(chǔ)上,評(píng)估了微重力、輻射等條件對(duì)節(jié)旋藻菌株的形態(tài)、結(jié)構(gòu)和生理學(xué)以及遺傳變化的影響[30]。
此外,為解決未來(lái)深空探測(cè)人類營(yíng)養(yǎng)素的原位按需補(bǔ)充,Ball 等[31]的“Bionutrients”實(shí)驗(yàn)搭建了1 個(gè)用于實(shí)現(xiàn)目標(biāo)營(yíng)養(yǎng)素的原位微生物生產(chǎn)平臺(tái)。 該實(shí)驗(yàn)使用能夠制造玉米黃素的工程酵母從可食用培養(yǎng)基中生成類胡蘿卜素,以補(bǔ)充長(zhǎng)期儲(chǔ)存的食物中潛在的維生素?fù)p失。 在大量地面研究的基礎(chǔ)上,針對(duì)微生物生物部件的空間驗(yàn)證實(shí)驗(yàn)正在逐步展開,這也是人類未來(lái)向地外軌道生物基生命支持系統(tǒng)開發(fā)邁出的重要一步,對(duì)于未來(lái)長(zhǎng)期飛行任務(wù)的執(zhí)行中生物質(zhì)的生產(chǎn)以及發(fā)射成本的控制等都具有重要意義。 同時(shí),這些技術(shù)在未來(lái)地面食品生產(chǎn)、環(huán)境保護(hù)等領(lǐng)域的應(yīng)用也極具潛力。
4.3 微生物在深空探測(cè)中的原位應(yīng)用
原位資源利用(In-situ Resource Utilization,ISRU)是指利用地外目標(biāo)星體的資源生產(chǎn)載人航天任務(wù)所需要的物資資源,如氧氣和水,進(jìn)而提高飛行任務(wù)中的安全性和經(jīng)濟(jì)可承受性。 盡管當(dāng)前原位資源利用技術(shù)仍處于初級(jí)階段,基于生物的原位資源利用仍表現(xiàn)出巨大的應(yīng)用前景。 Montague 等[32]認(rèn)為微生物可以利用月球上大量存在的含碳物質(zhì)、水分生產(chǎn)食物。 Hogan 等[33]在國(guó)際空間站上開展的“Micro-12”實(shí)驗(yàn)通過(guò)研究微重力條件下胞外呼吸模式菌Shewanellaoneidensis MR-1的生物膜形成、細(xì)胞外電子傳遞特征,以期將其用于生物電化學(xué)系統(tǒng)(BES)中微生物電合成過(guò)程。Loudon 等[34]的“Biorock”實(shí)驗(yàn)著眼于火星巖石在未來(lái)原位資源中的應(yīng)用前景,通過(guò)研究鞘氨醇單胞菌、枯草芽孢桿菌及耐金屬貪銅菌對(duì)巖石的營(yíng)養(yǎng)物質(zhì)利用和對(duì)巖石蝕變的影響,為未來(lái)基于微生物的空間開采技術(shù)奠定基礎(chǔ),提高未來(lái)的原位資源利用能力。
5 微生物在天體生物學(xué)研究中的應(yīng)用
研究地球及地外星球的生命起源、進(jìn)化、生命在宇宙中的分布以及生命和環(huán)境相互作用,進(jìn)而在太陽(yáng)系內(nèi)、外尋找生命及宜居環(huán)境是天體生物學(xué)的主要研究?jī)?nèi)容。 地球上極端環(huán)境中(包括鹽湖、旱地、深海、極地、永久凍土等)微生物的存在使人類推測(cè)微生物能夠在地外極端空間環(huán)境下生存。 由于生命的起源過(guò)程不可能在實(shí)驗(yàn)室里完整復(fù)現(xiàn),因此,將微生物用作代表性的生物模型系統(tǒng),在宇宙進(jìn)化的框架下關(guān)注地球生命體在真實(shí)空間環(huán)境下的存活、耐受和適應(yīng)機(jī)制是開展天體生物學(xué)研究的主要方法。 目前,歐美國(guó)家已經(jīng)開發(fā)了多種用于短期和長(zhǎng)期微生物空間暴露實(shí)驗(yàn)的生物學(xué)裝置,能夠針對(duì)不同實(shí)驗(yàn)需求提供相應(yīng)實(shí)驗(yàn)環(huán)境,實(shí)現(xiàn)了對(duì)細(xì)菌、孢子、地衣、真菌等生命體的空間暴露實(shí)驗(yàn)[3,6,35-36]。
在太陽(yáng)系的各大行星中,火星是最“類地”的行星,和地球環(huán)境不同的是,由于缺乏內(nèi)在磁場(chǎng)、表面稀薄的大氣對(duì)輻射的屏蔽能力極弱,火星幾乎是持久地暴露于空間輻射場(chǎng)中,這種環(huán)境與早期地球環(huán)境極為相似。 因此,揭示地球微生物在火星表面環(huán)境的生存與繁衍方式,對(duì)于探討地球生命起源和火星生命探測(cè)至關(guān)重要。 多次的空間暴露實(shí)驗(yàn)[37-38]表明,火星上的紫外輻射(λ ≥200 nm)是影響微生物存活的主要限制因子。UV-B、UV-C 射線能夠誘發(fā)生物體DNA 鏈中相鄰的嘧啶堿基產(chǎn)生嘧啶二聚體,阻礙DNA 的復(fù)制和堿基的正常配對(duì),使DNA 空間構(gòu)象發(fā)生改變,進(jìn)而影響轉(zhuǎn)錄以及蛋白質(zhì)的生物功能。 短波的真空紫外輻射會(huì)被生物DNA 分子吸收,直接或間接導(dǎo)致生物體DNA 鏈斷裂,引發(fā)生物的高突變率或者致死。 Wassman 等[38]將具有抗紫外輻射能力的枯草芽孢桿菌株MW01 在地球低軌道環(huán)境中搭載559 天發(fā)現(xiàn)只有少數(shù)的芽孢能夠存活,表明太陽(yáng)紫外輻射對(duì)枯草芽孢桿菌的芽孢極具破壞力。
相比之下,溫度、壓力和宇宙電離輻射等環(huán)境因素對(duì)微生物孢子的存活率影響較小[4,6]。 特別是,向微生物加以適當(dāng)?shù)谋Wo(hù)結(jié)構(gòu)對(duì)于提高微生物對(duì)火星環(huán)境的耐受性極為重要。 在生物盤(Biopan)上開展的生存實(shí)驗(yàn)中,暴露的枯草芽孢桿菌在經(jīng)過(guò)2 周飛行后,返回地面時(shí)全部失活,而被包被在泥土、隕石層或者鹽晶中的芽孢的存活率則為100%[7,39]。 Pacelli 等[40]將石生微生物空間暴露了548 天后生物類群仍能存活,在一定程度上支持了微生物能夠在星際塵埃或者隕石中存活、運(yùn)移的可能性,同時(shí)也表明石生微生物在宇宙生命起源研究中的應(yīng)用潛力。
隨著人類航天活動(dòng)的日趨頻繁,更多的天體生物學(xué)研究通過(guò)空間暴露的方式開展,但許多研究?jī)H關(guān)注于空間單一因素對(duì)生命體的影響。Nicholson 等[41]對(duì)枯草芽孢桿菌的轉(zhuǎn)錄組學(xué)分析顯示:地面單一模擬環(huán)境對(duì)枯草芽孢桿菌孢子的破壞性小于真實(shí)空間環(huán)境,因此,天體生物學(xué)研究需要更多地考慮空間復(fù)合環(huán)境對(duì)微生物造成的協(xié)同效應(yīng)。 此外,受當(dāng)前空間實(shí)驗(yàn)技術(shù)能力的限制,多數(shù)實(shí)驗(yàn)主要圍繞休眠態(tài)微生物的生存能力評(píng)估展開,未來(lái)的研究中只有開發(fā)多種生物技術(shù)手段,才能實(shí)現(xiàn)從休眠態(tài)到代謝旺盛的微生物,從基因到個(gè)體,甚至群落適應(yīng)空間真實(shí)環(huán)境的生物學(xué)機(jī)制,在此基礎(chǔ)上,才能利用微生物開發(fā)、預(yù)測(cè)和假設(shè)空間環(huán)境中存在生命的可能性。
6 空間微生物技術(shù)轉(zhuǎn)化與應(yīng)用
微生物在多個(gè)空間環(huán)境因素的協(xié)同作用下,其基礎(chǔ)生物學(xué)特征以及致病性、抗性等均會(huì)產(chǎn)生變化,甚至產(chǎn)生一些地面環(huán)境中難以獲得的生物特性。 將微生物實(shí)驗(yàn)技術(shù)和空間生物工程研究相結(jié)合,進(jìn)而實(shí)現(xiàn)空間微生物技術(shù)研究與轉(zhuǎn)化是微生物應(yīng)用的重要形式。
在空間飛行任務(wù)中,航天員表現(xiàn)出心血管功能失調(diào)、骨質(zhì)流失、肌肉萎縮等癥狀,解決航天員在空間飛行環(huán)境下的生存、健康和工效的問(wèn)題是載人航天的首要問(wèn)題。 研究表明,藥物在空間環(huán)境中施用時(shí)的表現(xiàn)與在地球上表現(xiàn)不同。 酵母是一種適用于多種健康疾病的模式生物,Zea[42]利用釀酒酵母在微重力環(huán)境下評(píng)估了二甲雙胍影響細(xì)胞代謝途徑,了解微重力條件下藥物在細(xì)胞內(nèi)的作用途徑,進(jìn)而提高藥物在空間環(huán)境下的有效性。 Johanson 等[43]、Coleman 等[44]在空間環(huán)境下分析了Msn4、Sfp1 介導(dǎo)的酵母細(xì)胞簇的遺傳分化,進(jìn)而為尋找抑制腫瘤細(xì)胞簇的新藥或?qū)ふ倚碌幕虬悬c(diǎn)奠定基礎(chǔ)。
利用空間特殊條件篩選產(chǎn)率高、活性強(qiáng)的生產(chǎn)用菌也是微生物技術(shù)應(yīng)用的重要方面。 利用微生物開發(fā)抗病毒研究和癌癥治療的γ 干擾素、治療肺氣腫的彈性蛋白酶、治療糖尿病的胰島素等已經(jīng)取得很大的進(jìn)展[45]。 更重要的是,將空間環(huán)境作為創(chuàng)新平臺(tái),利用微生物工程菌實(shí)現(xiàn)藥品、食品、化學(xué)品的制備,能夠促進(jìn)空間生物技術(shù)向生物發(fā)酵、生物化工以及生物基材料生產(chǎn)等工業(yè)領(lǐng)域滲透和轉(zhuǎn)化。 如NASA Briggs 研究團(tuán)隊(duì)[46]利用微重力環(huán)境改變大腸桿菌工程菌異丁烯合成的代謝網(wǎng)絡(luò),進(jìn)而指導(dǎo)空間環(huán)境下異丁烯生產(chǎn);Birlem等[47]通過(guò)開展空間特殊環(huán)境下產(chǎn)甲烷菌的培養(yǎng),旨在尋找未來(lái)地面燃料的替代品。 Nickerson等[48]在空間培養(yǎng)重組減毒的沙門氏菌疫苗(RASV)菌株,通過(guò)提高療效和保護(hù)性免疫反應(yīng)促進(jìn)下一代疫苗的設(shè)計(jì)和開發(fā)。
7 結(jié)束語(yǔ)
探討空間環(huán)境對(duì)微生物生命體的影響,其最終目標(biāo)是為維持人類的空間探索可持續(xù)發(fā)展制定相應(yīng)的對(duì)策。 開展微生物在空間特殊環(huán)境下的生命現(xiàn)象及其活動(dòng)規(guī)律的基礎(chǔ)研究,特別是長(zhǎng)期處于空間條件下微生物生存、變異、以及與宿主生物的相互作用機(jī)制,對(duì)于長(zhǎng)期航天任務(wù)的安全性和可持續(xù)性開展具有重要的支撐作用。 同時(shí),立足于微生物在空間環(huán)境下獨(dú)特的代謝特征,利用空間微生物技術(shù)指導(dǎo)地面生產(chǎn),挖掘人類對(duì)空間資源利用潛力,也是未來(lái)我國(guó)空間站建成后航天技術(shù)應(yīng)用的重要方面。 因此,如何最大化利用空間微生物科學(xué)與技術(shù)成果,將是我國(guó)載人航天工程發(fā)展以及空間微生物研究的重要課題。
參考文獻(xiàn)(References)
[ 1] Horneck G,Moeller R,Cadet J,et al. Resistance of bacterial endospores to outer space for planetary protection purposes-experiment PROTECT of the EXPOSE-E mission [J]. Astrobiology, 2012, 12(5):445-456.
[ 2] de Vera J P, Alawi M, Backhaus T, et al. Limits of life and the habitability of Mars: The ESA space experiment BIOMEX on the ISS [J]. Astrobiology, 2019, 19(2):145-157.
[ 3] Horneck G, Rettberg P. Complete Course in Astrobiology[M]. Wiley-VCH, 2007: 273-320.
[ 4] Karouia F, Peyvan K, Pohorille A. Toward biotechnology in space: High-throughput instruments for in situ biological research beyond earth [J]. Biotechnology Advances, 2017,35:905-932.
[ 5] Zeitlin C, Hassler D M, Cucinotta F A, et al. Measurements of energetic particle radiation in transit to Mars on the Mars Science Laboratory [J]. Science, 2013, 340(6136): 1080-1084.
[ 6] 袁俊霞, 張美姿, 印紅,等. 空間環(huán)境對(duì)微生物的影響及應(yīng)用[J]. 載人航天, 2016, 22(4): 500-506.
Yuan J X, Zhang M Z, Yin H, et al. Effect of space environment on microorganisms and its applications [J]. Manned Spaceflight, 2016, 22(4): 500-506. (in Chinese)
[ 7] Benoit M R, Li W, Stodieck L S, et al. Microbial antibiotic production aboard the International Space Station [J]. Applied Microbiology and Biotechnology, 2006, 70(4): 403-411.
[ 8] Hammond T G, Stodieck L S, Birdsall H H, et al. Effects of microgravity on the virulence of Salmonella toward Caenorhabditis elegans[J]. New Space, 2013, 1(3): 123-131.
[ 9] Zea L. Phenotypic and Gene Expression Responses of E. coli to Antibiotics during Spaceflight[D]. Boulder:University of Colorado at Boulder, 2015.
[10] Kim W, Tengra F K, Young Z, et al. Spaceflight promotes biofilm formation by Pseudomonas aeruginosa[J]. Plos One,2013, 8(4): e62437.
[11] Urbaniak C, Sielaff A C, Frey K G, et al. Detection of antimicrobial resistance genes associated with the International Space Station environmental surfaces [J]. Scientific Reports,2018, 8(1): 814.
[12] Kim W, Tengra F K, Shong J, et al. Effect of spaceflight on Pseudomonas aeruginosa final cell density is modulated by nutrient and oxygen availability [J]. BMC Microbiology, 2013,13(1): 241.
[13] Takahashi A, Ohnishi K, Takahashi S. The effects of microgravity on induced mutation in Escherichia coli and Saccharomuces cerevisiae [J]. Advances in Space Research, 2001,28(4): 555-561.
[14] Wilson J W, Ott C M, Quick L, et al. Media ion composition controls regulatory and virulence response of Salmonella in spaceflight [J]. Plos One, 2008, 3(12):e3923.
[15] Sittka A,Pfeiffer V,Tedin K,et al. The RNA chaperone Hfq is essential for the virulence of Salmonella typhimurium [J].Molecular Microbiology, 2007, 63(1): 193-217.
[16] Grant K C, Khodadad C L M, Foster J S. Role of Hfq in an animal-microbe symbiosis under simulated microgravity conditions [J]. International Journal of Astrobiology, 2014, 13(1):53-61.
[17] Wilson J W,Ottc C M,HonerzuBentrup K,et al. Space flight alters bacterial gene expression and virulence and reveals a role for global regulator Hfq [J]. Proceedings of the National Academy of Sciences, 2007,104 (41):16299-16304.
[18] Wilson J W,Ott C M,Ramamurthy R,et al. Low-Shear modeled microgravity alters the Salmonella enteric serovar Typhimurium stress response in an RpoS-independent manner [J].Applied and Environmental Microbiology, 2002, 68(11):5408-5416.
[19] Yamaguchi N,Roberts MS,Castro S,et al. Microbial monitoring of crewed habitats in space-current status and future perspectives [J]. Microbes and Environments, 2014, 29(3):250-260.
[20] Ichijo T,Yamaguchi N,Tanigaki F,et al. Four-year bacterial monitoring in the International Space Station-Japanese Experiment Module “Kibo” with culture-independent approach [J].npj Microgravity, 2016, 2: 16007.
[21] McLean R J,Cassanto J M,Barnes M B, et al. Bacterial biofilm formation under microgravity conditions [J]. FEMS Microbiology Letters, 2001, 195: 115-119.
[22] Sielaff A C, Singh N K, Allen J E, et al. Draft genome sequences of biosafety level 2 opportunistic pathogens isolated from the environmental surfaces of the International Space Station [J]. Genome Announcements,2016,4(6):e01263-16.
[23] Singh N K, Blachowicz A, Checinska A, et al. Draft genome sequences of two Aspergillus fumigatus strains, isolated from the International Space Station [J]. Genome Announcements,2016, 4(4): e00553-16.
[24] Singh N K, Wood J M, Karouia F, et al. Succession and persistence of microbial communities and antimicrobial resistance genes associated with International Space Station environmental surfaces [J]. Microbiome, 2018, 6(1): 204.
[25] Knox B P, Blachowicz A,Palmer J M,et al. Characterization of Aspergillus fumigatus isolates from air and surfaces of the International Space Station [J]. mSphere, 2016, 1(5):e00227-16.
[26] Be N A, Avila-Herrera A, Allen J E, et al. Whole metagenome profiles of particulates collected from the International Space Station [J]. Microbiome, 2017, 5(1): 81.
[27] 袁俊霞, 印紅, 趙彪,等. 航天器AIT 中心微生物多樣性分析[J]. 空間科學(xué)學(xué)報(bào), 2017, 37(2): 185-191.Yuan J X, Yin H, Zhao B, et al. Microbial diversity analysis in spaceflight AIT center [J]. Chinese Journal of Space Science, 2017, 37(2): 185-191. (in Chinese)
[28] 張文德, 印紅, 徐侃彥. 一種適用于空間條件的材料防霉性能試驗(yàn)方法[J]. 載人航天, 2017, 23(3): 419-426.
Zhang W D,Yin H,Xu K Y. A method for testing anti-mould properties of materials under space environment [J].Manned Spaceflight, 2017, 23(3): 419-426. (in Chinese)
[29] Lasseur C, Brunet J,de Weever H,et al. MELiSSA:the European project of closed life support system [J]. Gravitational and Space Biology, 2010, 23(2): 3-12.
[30] Leys N, Dussap G, Farge B. Arthrospira sp. Gene expression and mathematical modelling on cultures grown in the International Space Station [EB/OL].(2017)[2019-10-08].https:/ /www.nasa.gov/mission_pages/station/research/experiments/explorer/Investigation.html? #id =1821. html.
[31] Ball N, Kagawa H, Hindupur A, et al. Development of storage methods for Saccharomyces strains to be utilized for in situ nutrient production in long-duration space missions [C]/ /47thInternational Conference on Environmental Systems (ICES 2017). Charleston, SC, 2017, ARC-E-DAA-TN44786.
[32] Montague M, McArthur G H, Cockell C S, et al. The role of synthetic biology for in situ resource utilization (ISRU) [J].Astrobiology, 2012, 12(12):1135-1142.
[33] Hogan J, Dougherty M, Arkin A. Investigating the physiology and fitness of an exoelectrogenic organism under microgravity conditions (Micro-12) [EB/OL].(2018)[2019-10-10].https:/ /www.nasa.gov/mission_pages/station/research/experiments/explorer/Investigation.html? #id =7470. html.
[34] Loudon C, Nicholson N, Finster K, et al. BioRock: new experiments and hardware to investigate microbe-mineral interactions in space [J]. International Journal of Astrobiology,2017:1-11.
[35] Backhaus T, Mee?en J, Demets R, et al. Characterization of viability of the lichen Buellia frigida after 1.5 years in space on the International Space Station [J]. Astrobiology, 2019,19(2): 233-241.
[36] Billi D,Verseux C,F(xiàn)agliarone C,et al. A desert cyanobacterium under simulated Mars-like conditions in low earth orbit:implications for the habitability of Mars [J]. Astrobiology,2019, 19(2): 158-169.
[37] Noblet A,Stalport F,Guan Y Y,et al. The PROCESS experiment:amino and carboxylic acids under mars-like surface UV radiation conditions in low-earth orbit [J]. Astrobiology,2012, 12 (5): 436-444.
[38] Wassmann M,Moeller R,Rabbow E,et al. Survival of spores of the UV-resistant Bacillus subtilis strain MW01 after exposure to low-earth orbit and simulated Martian conditions:Data from the space experiment ADAPT on EXPOSE-E [J]. Astrobiology, 2012, 12(5): 498-507.
[39] Rettberg P, Rabbow E, Panitz C, et al. Biological space experiments for the simulation of Martian conditions: UV radiation and Martian soil analogues [J]. Advances in Space Research, 2004, 33(8):1294-1301.
[40] Pacelli C, Selbmann L, Zucconi L, et al. Responses of the black fungus Cryomyces antarcticus to simulated Mars and space conditions on rock analogs[J]. Astrobiology,2019,19(2): 209-220.
[41] Nicholson W L, Moeller R, Horneck G. Transcriptomic responses of germinating Bacillus subtilis spores exposed to 1.5 years of space and simulated martian conditions on the EXPOSE-E experiment PROTECT [J]. Astrobiology, 2012, 12(5): 469-486.
[42] Zea L. Drug discovery and development in space BioServe Space Technologies [C]/ / 66thInternational Astronautical Congress. Jerusalem, 2015: IAC-15-A1.8.1,x27627.
[43] Johanson K, Allen P L, Gonzalez-Villaobos R A, et al. Haploid deletion strains of Saccharomyces cerevisiae that determine survival during space flight [J]. Acta Astronautica,2007, 60(4-7): 460-471.
[44] Coleman C B, Allen P L, Rupert M, et al. Novel Sfp1 transcriptional regulation of Saccharomyces cerevisiae gene expression changes during spaceflight[J]. Astrobiology, 2008, 8(6): 1071-1078.
[45] 程紹馳, 劉映國(guó). “國(guó)際空間站”空間科學(xué)與應(yīng)用發(fā)展及影響分析[J]. 國(guó)際太空, 2010, 12: 39-45.
Cheng S C, Liu Y G. Development and impact analysis of space science and application in the International Space Station [J]. Space International, 2010, 12: 39-45. (in Chinese)
[46] Briggs B. Tapping microgravity to enhance biofuel production[EB/OL].(2018) [2019-10-08].https:/ /www.nasa.gov/mission_pages/station/research/experiments/explorer/Investigation.html? #id=7783. html.
[47] Birlem M. Space Tango payload card MMARS-1 [EB/OL].(2019)[ 2019-09-10].https:/ /ww.nasa.gov/mission_pages/station/research/experiments/explorer/Investigation. html? #id =7551. html.
[48] Nickerson C A. Recombinant Attenuated Salmonella Vaccine(RASV) [EB/OL]. (2018)[2019-09-20 ].https:/ /www.nasa. gov/mission _ pages/station/research/experiments/explorer/Investigation.html? #id =301. html.
猜你喜歡
天天愛(ài)科學(xué)(2022年9期)2022-09-15 01:12:54
天天愛(ài)科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
中老年保健(2021年12期)2021-08-24 03:30:40
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
中國(guó)傳媒大學(xué)學(xué)報(bào)(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
軍事文摘(2020年20期)2020-11-28 11:42:50
中國(guó)生殖健康(2020年6期)2020-02-01 06:28:50
航空世界(2020年10期)2020-01-19 14:36:20
新世紀(jì)智能(英語(yǔ)備考)(2019年12期)2020-01-13 06:07:18
