Tagitinin D誘導人肝癌HepG2細胞凋亡的作用機制研究
2020-05-16 04:14:00韋睿然陳遠志楊杏芝何志旭
天然產(chǎn)物研究與開發(fā) 2020年3期
楊 珊,韋睿然,陳遠志,楊 旭,王 娟,楊杏芝,何志旭*,丁 驍*
1貴州醫(yī)科大學基礎醫(yī)學院 免疫學教研室組織工程與干細胞中心,貴陽 550004;2中國科學院昆明植物研究所 植物化學與西部植物資源持續(xù)利用國家重點實驗室,昆明 650201
肝癌是臨床上最常見的惡性腫瘤之一。近年來,肝癌已成為中國發(fā)病率和死亡率較高的癌癥,是全球癌癥死亡的第三大原因[1,2];肝癌的治療包括外科手術、介入治療、化學療法和肝移植,但治療效果并不理想[3]。盡管化學療法極大地提高了患者的生存率,但嚴重的毒副作用阻礙了抗癌藥物的治療效果[4]。尋找對腫瘤細胞具有抑制作用的天然小分子化合物,是抗癌藥物發(fā)現(xiàn)的重要來源。
Tagitinin D(圖1)屬于倍半萜內酯類天然小分子化合物[5],在菊科植物中大量存在。Tagitinin D類化合物具有抗腫瘤[6]、細胞毒性[7]、抗炎[8]、抗菌[9]、抗真菌[10]特性。據(jù)報道tagitinin D的類似物tagitinin A和tagitinin C 能減少種子發(fā)芽和幼苗生長[11];tagitinin C也具有多種生物活性,包括抗癌作用,且tagitinin C衍生物具有作為癌癥治療的巨大潛力[12]。但是對tagitinin D的生物活性研究較少。本論文用人肝癌HepG2細胞為研究對象,探討tagitinin D對肝癌細胞的增殖、凋亡的影響,并檢測ER stress信號通路相關蛋白的變化,探究其抑制腫瘤的可能機制,為開發(fā)利用其藥用價值提供科學依據(jù)。
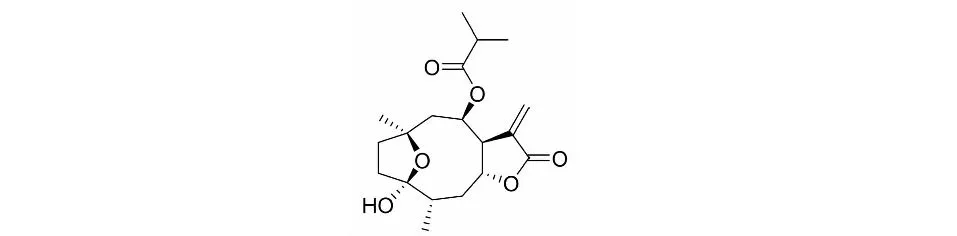
圖1 Tagitinin D的化學結構Fig.1 The chemical structure of tagitinin D
1 材料與儀器
1.1 受試細胞株
人肝癌HepG2細胞株由中國科學院昆明植物研究所活性篩選中心孔清華老師惠贈。
1.2 主要試劑
Tagitinin D,購于云南西力生物技術有限公司,純度99%,碘化丙啶PI、溴化四氮唑藍MTT(Biosharp生物公司),Annexin V-FITC/PI雙染細胞凋亡檢測試劑盒(四正柏生物),二甲基亞砜DMSO(天津市風船化學試劑科技有限公司),高糖DMEM培養(yǎng)基、胎牛血清、胰蛋白酶、雙抗(青霉素10 000 U/mL、鏈霉素10 mg/mL)(GIBCO),抗體:Actin、PDI、Calnexin、Bip、Ero1-Lα、IRE1α、p-eIF2α(CST)。
1.3 主要儀器
BD FACS Celesta3激光流式細胞儀;成像型微孔板檢測儀(BioTek Cytationl);全自動化學發(fā)光圖像分析系統(tǒng)(tanon5200);立式自動壓力蒸汽滅菌器(GR60DR);1300系列Ⅱ級A2型生物安全柜(Thermo);SCILOGEX離心機(CF1524R)。
2 實驗方法
2.1 細胞培養(yǎng)
用含10%的FBS(胎牛血清)、1%雙抗(青霉素10 000 U/mL、鏈霉素10 mg/mL)的高糖DMEM培養(yǎng)基培養(yǎng)HepG2細胞,放置在37 ℃、5% CO2培養(yǎng)箱中,取對數(shù)生長期且細胞狀態(tài)良好的細胞用于后續(xù)實驗。
2.2 檢測人肝癌HepG2細胞活力
采用溴化四氮唑藍(MTT)法檢測細胞活力[13]。取對數(shù)生長期細胞,胰酶消化后吹打呈單細胞懸液將其接種于96孔板,每孔100 μL,待細胞貼壁后,加入不同濃度tagitinin D,每個濃度作4個復孔,置培養(yǎng)箱中72 h,棄上清,每孔中加入100 μL MTT溶液(5 mg/mL),放入培養(yǎng)箱中繼續(xù)培養(yǎng)4 h,棄上清后再加入100 μL DMSO,用多功能酶標儀在波長490 nm處測量各孔的吸光度(OD值),按下列公式計算細胞活力:
細胞活力=處理組平均OD值/
對照組平均OD值 × 100%
2.3 克隆形成實驗
取對數(shù)生長期細胞胰酶消化后呈單細胞懸液,用細胞計數(shù)板計數(shù),按300個/mL接種于6孔板中,每孔2 mL,待細胞貼壁后,加入不同濃度的tagitinin D(1和2 μM),繼續(xù)培養(yǎng)12天,棄培養(yǎng)基,PBS洗三遍,每孔加入考馬斯亮藍R250(2.5 mg/mL)1 mL染色2 h,棄染色液,PBS洗三遍,拍照。
2.4 細胞周期檢測
取對數(shù)生長期細胞胰酶消化后呈單細胞懸液,接種于6孔板,待細胞長到80%左右,加入不同濃度tagitinin D處理12 h,收集細胞制成單細胞懸液,70%乙醇4 ℃過夜,PBS洗一遍,加入500 μL PBS重懸細胞,加入20 μL RNase,放37 ℃孵育30 min,再加入50 μL PI,冰上放置30 min,流式細胞儀檢測細胞周期。
2.5 細胞凋亡檢測
取對數(shù)生長期細胞制呈單細胞懸液,接種于6孔板,待細胞長到80%左右,加入tagitinin D 20 μM處理不同時間(24、48、72 h),收集細胞制成單細胞懸液,PBS洗一遍,按凋亡試劑盒說明書加入試劑,用流式細胞儀檢測細胞凋亡率。
2.6 檢測活性氧的變化
取對數(shù)生長期細胞制呈單細胞懸液,接種于6孔板,待細胞長到80%左右,加入不同濃度tagitinin D處理12 h,棄培養(yǎng)基,加入無血清培養(yǎng)基2 mL,再加入DCFH-DA(20 μM),室溫孵育15 min,棄培養(yǎng)基,加入PBS洗三遍,拍照,再收集細胞用流式細胞儀檢測熒光強度。
2.7 Western blot檢測ER stress相關蛋白表達
取對數(shù)生長期細胞制呈單細胞懸液,接種于6孔板,待細胞長到80%左右,加入tagitinin D 6 μM處理不同時間(12、24、36 h),收集細胞制作蛋白樣本。10%聚丙烯酰胺SDS凝膠電泳后轉至PVDF膜上,5%脫脂牛奶封閉2 h,加入一抗4 ℃孵育過夜,二抗室溫2 h,PBST洗滌3次,加化學發(fā)光液放入全自動化學發(fā)光圖像分析系統(tǒng)中曝光。
2.8 統(tǒng)計學分析
實驗重復3次且每組至少3個復孔,用GraphPad Prism 7軟件進行統(tǒng)計分析處理,結果以均值±標準差表示,用t檢驗進行組間差異比較。P<0.05差異具有統(tǒng)計學意義。
3 結果與分析
3.1 Tagitinin D對人肝癌HepG2細胞活力的影響
用MTT法檢測tagitinin D對HepG2細胞活力的影響,加入不同濃度tagitinin D處理細胞72 h,用酶標儀檢測吸光度值,計算出tagitinin D作用于HepG2細胞的活力及IC50值= 7.79 ± 0.58 μM,如圖2所示,說明tagitinin D能抑制HepG2細胞活力。
3.2 Tagitinin D對HepG2細胞體外成瘤能力的抑制

圖2 Tagitinin D抑制HepG2細胞活力Fig.2 Tagitinin D inhibited cell viability of HepG2 cells
Tagitinin D不同濃度(1和2 μM)作用于HepG2細胞12天后,兩個處理組克隆球的數(shù)量和大小明顯低于對照組(圖3A),說明tagitinin D能顯著抑制腫瘤細胞體外成瘤能力。
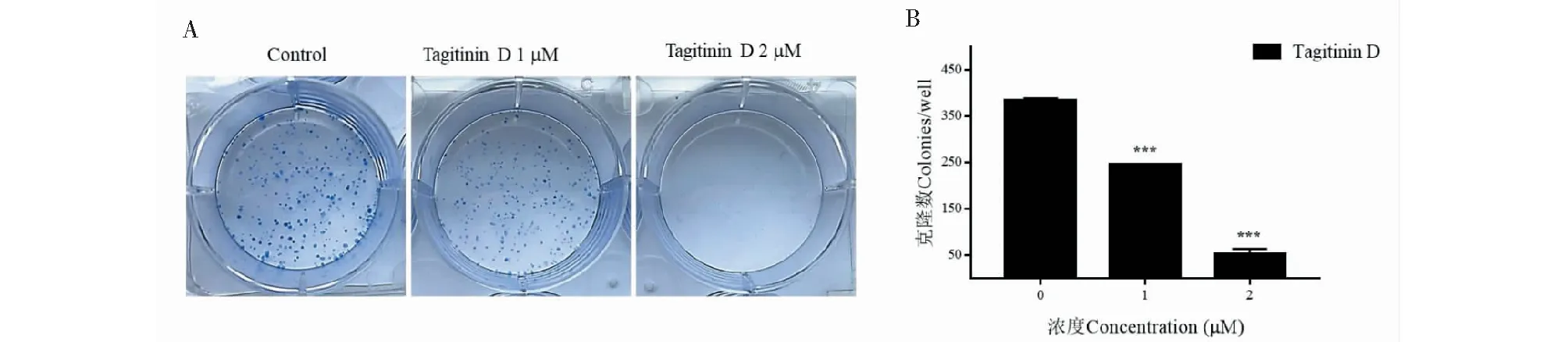
圖3 Tagitinin D對HepG2細胞體外成瘤能力的影響Fig.3 Tagitinin D inhibited the colony formation ability of HepG2 cells
3.3 Tagitinin D使人肝癌HepG2細胞周期阻滯在G2/M期
用流式細胞儀檢測tagitinin D對HepG2細胞周期的影響,如圖4A所示:tagitinin D處理組細胞的S期比例由26.1%下降到17.2%,G2/M期比例由28.9%升高至42.8%,說明tagitinin D處理可將大部分細胞阻滯在G2/M期。

圖4 Tagitinin D對HepG2細胞周期的影響Fig.4 Tagitinin D induced cell cycle arrest at the G2/M phase in HepG2 cells
3.4 Tagitinin D誘導人肝癌HepG2細胞凋亡
用流式細胞儀檢測tagitinin D誘導人肝癌HepG2細胞凋亡,如圖5A所示,當處理濃度為20 μM,處理時間為24、48、72 h細胞凋亡率(早期凋亡+晚期凋亡)分別為22%、59.7%、96.6%;說明tagitinin D促進細胞凋亡是時間依賴的。
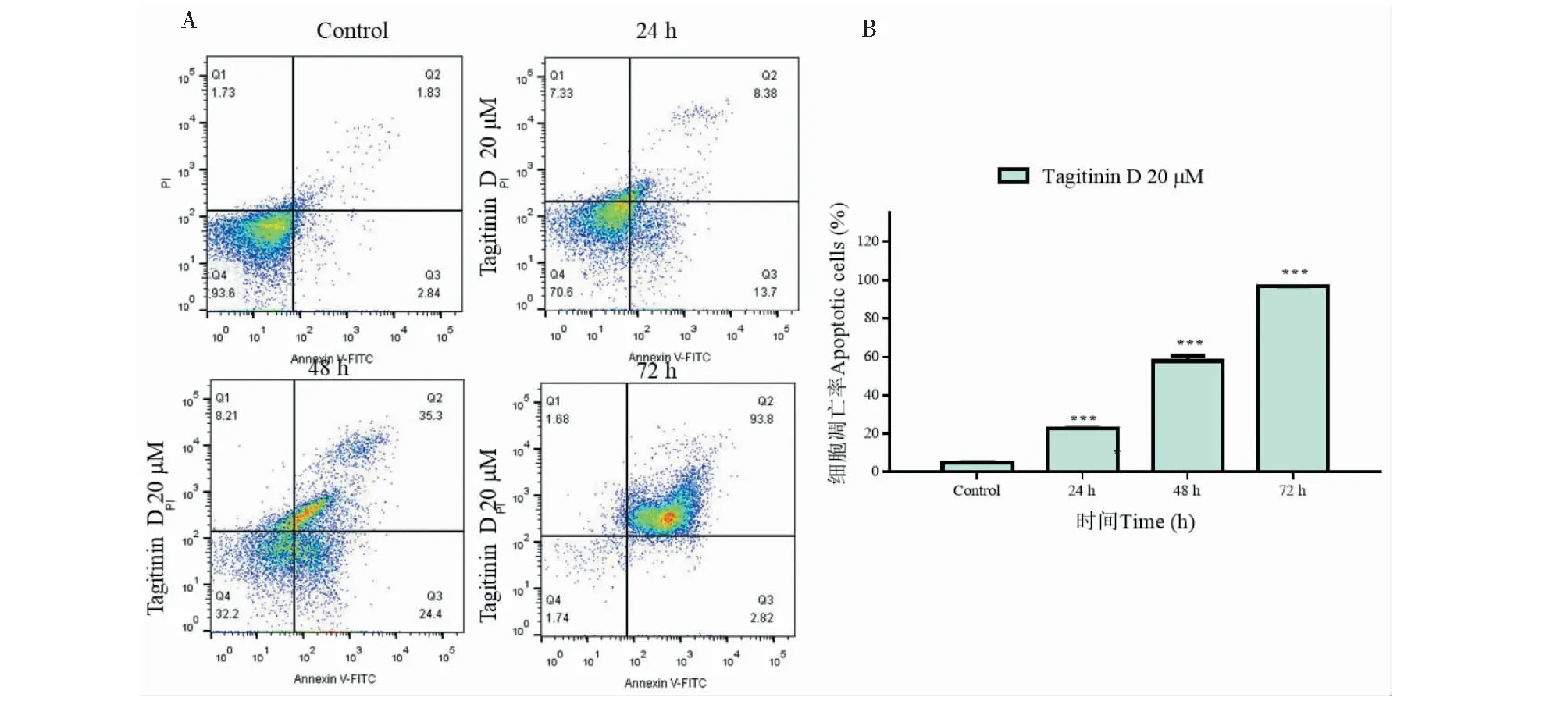
圖5 Tagitinin D促進HepG2細胞凋亡Fig.5 Tagitinin D induced apoptosis of HepG2 cells in a time-dependent manner
3.5 Tagitinin D增加人肝癌HepG2細胞內活性氧
3.5.1 Tagitinin D和NAC聯(lián)用使人肝癌HepG2細胞活力回復
為研究tagitinin D誘導人肝癌HepG2細胞凋亡的作用途徑,加入活性氧(reactive oxygen species,ROS)清除劑NAC和泛caspases抑制劑Z-VAD-FMK與tagitinin D聯(lián)合處理,如圖6所示:加入Z-VAD-FMK,細胞活力沒有回復;加入NAC后,細胞活力明顯回復。以上結果說明tagitinin D降低腫瘤細胞活力是依賴于ROS途徑的。

圖6 Tagitinin D和NAC聯(lián)用使人肝癌HepG2細胞活力回復Fig.6 The cell death induced by tagitinin D can be rescued by the ROS scavenger,NAC
3.5.2 DCFH-DA檢測人肝癌HepG2細胞內活性氧變化
用探針DCFH-DA檢測細胞內源活性氧的累積。如圖7所示,隨著tagitinin D處理濃度增加,熒光強度逐漸增強(圖7A)及平均熒光強度增加(圖7B);加NAC后,熒光強度有所回復;說明tagitinin D能顯著引起細胞內源性ROS的增加。
3.6 Tagitinin D對人肝癌HepG2細胞ER stress 相關蛋白的影響
ER stress作為ROS的下游,并且持續(xù)、強烈的ER stress能顯著促進細胞凋亡。既然tagitinin D促進細胞凋亡依賴于ROS,且不依賴于caspases途徑,因此檢測tagitinin D對ER stress的影響是必要的。如圖8所示,tagitinin D處理組濃度6 μM,隨著處理時間的變化,ER stress相關蛋白PDI、Ero1-Lα、Calnexin、Bip、IRE1α、p-eIF2α表達水平升高,表明tagitinin D是通過增加ROS引起ER stress并激活IRE1α和PERK信號通路進而誘導細胞凋亡。

圖7 Tagitinin D使HepG2細胞內源活性氧增加Fig.7 Tagitinin D increased the endogenous ROS levels in HepG2 cells

圖8 Tagitinin D激活ER stressFig.8 Tagitinin D activated the ER stress signaling pathway in HepG2 cells
4 討論與結論
近年來,由于抗癌藥物的局限性,天然小分子化合物為新藥的研發(fā)起到促進作用,有報道抗癌藥物中一半來源于天然產(chǎn)物及其衍生物[14,15],天然產(chǎn)物因具有廣泛的不同化學結構從而其生物學活性也有區(qū)別,包括抗菌、免疫抑制、抗癌、抗炎等[16]。因此,發(fā)現(xiàn)更多天然產(chǎn)物的藥用價值以期為臨床研究提供依據(jù)是必不可少的。本文主要從周期阻滯、誘導細胞凋亡及ER stress信號通路的影響研究tagitinin D促進人肝癌HepG2細胞凋亡的作用機制。
本實驗中,我們首先用MTT法檢測tagitinin D對人肝癌HepG2細胞活力的影響,實驗結果顯示隨著藥物處理濃度的增加,HepG2細胞活力顯著降低;通過克隆形成實驗表明,與對照組相比,隨著藥物處理濃度的增加,處理組克隆球形成變小且數(shù)量減少;用流式細胞儀檢測細胞周期,發(fā)現(xiàn)經(jīng)過tagitinin D處理后且隨著濃度的增加,G2/M期比例逐漸升高,以上結果說明tagitinin D能顯著抑制HepG2細胞的活力和成瘤能力,并且將大部分細胞阻滯在G2/M期。
內質網(wǎng)途徑是細胞凋亡途徑之一[17],越來越多的證據(jù)表明,內質網(wǎng)應激可能與ROS產(chǎn)生有關,并誘導細胞凋亡[18,19]。ROS是指超氧化物、過氧化氫、過氧亞硝酸鹽和羥自由基,他們是不穩(wěn)定的分子種類,可以破壞細胞內的蛋白質和脂質,并激活細胞內信號傳導級聯(lián)[20]。有研究表明,雷公藤甲素能增加細胞內ROS誘導內質網(wǎng)應激使細胞凋亡[21]。本研究通過凋亡和ROS檢測實驗結果發(fā)現(xiàn),tagitinin D誘導HepG2細胞凋亡且細胞內ROS增加,說明tagitinin D通過依賴于ROS誘導細胞凋亡。
內質網(wǎng)應激由多種因素引起,如鈣離子失調、ROS積累等造成大量未折疊蛋白和錯誤折疊蛋白聚集,引發(fā)ER stress[22]。一旦產(chǎn)生這種應激時間過長,內質網(wǎng)穩(wěn)態(tài)失衡,就會激活內質網(wǎng)凋亡信號,引起細胞凋亡。為進一步探討tagitinin D是否通過細胞內ROS積累激活ER stress信號通路或caspases途徑引起細胞凋亡。加入ROS清除劑NAC和泛caspases抑制劑Z-VAD-FMK后,明顯看到加NAC后細胞活力回復;用探針DCFH-DA檢測到細胞內ROS的增加;Western blot結果可得:PDI、Ero1-Lα、Calnexin、Bip、IRE1α、p-eIF2α蛋白表達增加;說明tagitinin D通過細胞內ROS增加,激活ER stress信號通路誘導細胞凋亡。
綜上所述,tagitinin D抑制人肝癌HepG2細胞活力和體外成瘤能力;通過阻滯細胞在G2/M期影響細胞增殖;同時使細胞內ROS的增加激活ER stress信號通路誘導細胞凋亡。
猜你喜歡
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
天津醫(yī)科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫(yī)藥(2015年16期)2015-03-01 02:03:11
