脫落酸與水楊酸處理調(diào)節(jié)李果實(shí)抗冷性及氧化酶活性
2020-05-27 09:41:48張一冉王雅楠楊楊韓育梅
江蘇農(nóng)業(yè)學(xué)報(bào) 2020年2期
張一冉 王雅楠 楊楊 韓育梅


摘要:為研究水楊酸(SA)提高李果實(shí)抗冷能力過程中脫落酸(ABA)的作用,本研究以黑琥珀李果實(shí)為原料,分別采用1.0 mmol/L SA、47.0 μmol/L ABA、5.0 mmol/L的Na2WoO4、1.0 mmol/L SA結(jié)合5.0 mmol/L的Na2WoO4以及蒸餾水為對(duì)照浸泡30 min。對(duì)李果實(shí)冷害指數(shù)、總酚含量、過氧化物酶(POD)、多酚氧化酶(PPO)、超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)的活性進(jìn)行測(cè)定。結(jié)果表明,SA和ABA處理組降低了冷害指數(shù),提高了其總酚含量并延緩了POD和PPO活性的上升,還顯著地提高了SOD、CAT和APX等活性氧清除酶的活性,Na2WoO4、SA結(jié)合Na2WoO4處理組抗冷能力均低于對(duì)照組。說明SA和ABA可以延緩李果實(shí)的冷害,提高其抗冷性和活性氧代謝能力。Na2WoO4顯著抑制李果實(shí)的抗冷性,且該作用不受SA的影響,表明SA提高李果實(shí)抗冷性依賴于ABA。
關(guān)鍵詞:李果實(shí);冷害;活性氧代謝;抗冷性
中圖分類號(hào):S662.3文獻(xiàn)標(biāo)識(shí)碼:A文章編號(hào):1000-4440(2020)02-0471-06
Abstract:In order to study the role of abscisic acid (ABA) in the process of salicylic acid (SA) improving the cold resistance of plum fruit, Black Amber plum fruit was used as raw material and soaked in 1.0 mmol/L SA, 47.0 μmol/L ABA, 5.0 mmol/L Na2WoO4, 1.0 mmol/L SA combined with 5.0 mmol/L Na2WoO4, and distilled water (control group) for 30 min. Chilling injury index, total phenolic content, peroxidase (POD) activity, polyphenol oxidase (PPO) activity, superoxide dismutase (SOD) activity, catalase (CAT) activity, ascorbate peroxidase (APX) activity were measured. The results showed that the SA and ABA treatments reduced the chilling injury index, increased the total phenolic content, delayed the increase of POD and PPO activities and significantly increased the activities of active oxygen scavenging enzymes such as SOD, CAT and APX. The cold resistance of Na2WoO4 and SA combined with Na2WoO4 treatment groups was lower than that of the control group. It is indicated that SA and ABA can delay the chilling injury of plum fruit and improve its cold resistance and active oxygen metabolism. Na2WoO4 significantly inhibited the cold resistance of plum fruit, and the effect was not affected by SA, which indicated that SA depended on ABA to increase the cold resistance of plum fruit.
Key words:plum fruit;chilling injury;active oxygen metabolism;cold resistance
李果實(shí)在低溫貯藏時(shí)極易發(fā)生冷害,造成果實(shí)腐爛變質(zhì),影響了產(chǎn)品貯藏品質(zhì)和商品價(jià)值[1]。使用小分子信號(hào)物質(zhì)調(diào)控果實(shí)冷害是近年來的研究熱點(diǎn)。已有研究結(jié)果表明,可以通過施用一些外源物質(zhì)(包括植物激素和植物生長(zhǎng)調(diào)節(jié)劑在內(nèi)的外源小分子物質(zhì))有效地調(diào)控活性氧代謝及抗氧化相關(guān)酶活性[2-4],控制果實(shí)冷害的發(fā)生。
在逆境脅迫下,活性氧大量積累超出機(jī)體自由清除能力后,導(dǎo)致活性氧代謝失衡,最終植物受到損傷[5]。近年來的國(guó)內(nèi)外研究結(jié)果表明,活性氧代謝失衡主要體現(xiàn)在清除活性氧自由基能力減弱、細(xì)胞膜脂過氧化程度加劇、抗氧化酶活性受到抑制等方面[6-7]。水楊酸(SA)可以提高保護(hù)酶的活性來清除活性氧,維持細(xì)胞膜系統(tǒng)的平衡,提高植物抵抗逆境脅迫的能力,從而達(dá)到保護(hù)植物細(xì)胞的作用。有研究結(jié)果表明,適宜濃度SA可以提高杏果實(shí)抗冷性及活性氧代謝[8]。類似的現(xiàn)象也在番茄[9]、桃[10]等果蔬中得到了驗(yàn)證。脫落酸(ABA)作為一種功能性激素,可以使植物對(duì)逆境做出快速應(yīng)答[11]。在低溫條件下,ABA可以通過調(diào)節(jié)植物體內(nèi)水分及維持細(xì)胞膜穩(wěn)定性多個(gè)方面來增加植物的低溫脅迫、鹽脅迫、水分脅迫的抵抗能力[12-13]。 ABA可以提高果實(shí)抗冷性,延長(zhǎng)果實(shí)貯藏期已在草莓[14]、香蕉[15]、桃[16]等多種果蔬中得到證實(shí)。黃杏等在研究中指出鎢酸鈉可以抑制內(nèi)源ABA合成并阻斷其信號(hào)傳遞,還指出鎢酸鈉可以抑制甘蔗幼苗的抗冷性和抗氧化能力[17]。但是SA和ABA對(duì)果實(shí)的抗冷信號(hào)是否存在交叉,二者之間相互關(guān)系怎樣,以及ABA是否參與了SA提高植物抗冷性的信號(hào)過程還有待研究。
因此,本研究采用SA、ABA、ABA合成抑制劑鎢酸鈉(Na2WoO4)以及SA結(jié)合Na2WoO4,蒸餾水為對(duì)照分別浸泡處理李果實(shí),通過研究其對(duì)冷害及活性氧相關(guān)酶活性的影響以明確SA、ABA處理減輕李果實(shí)冷害的作用,并通過Na2WoO4阻斷ABA的合成,探究SA提高李果實(shí)抗冷性是否依賴于ABA。
1材料與方法
1.1試驗(yàn)材料
李果實(shí)為中等熟度(果實(shí)顏色紫紅色,著色面積占2/3,香氣明顯,手感較軟)的黑琥珀李果實(shí),從內(nèi)蒙古呼和浩特市土左旗李子種植基地采摘獲得,大小、成熟度均一,無病蟲害。
1.2儀器與設(shè)備
TGL-16M高速冷凍離心機(jī)由上海盧湘離心機(jī)儀器有限公司生產(chǎn),T6新世紀(jì)型紫外分光光度計(jì)由北京普析通用儀器有限公司生產(chǎn),DDSJ-318電導(dǎo)率儀由上海儀電科學(xué)儀器股份有限公司生產(chǎn),電熱恒溫水浴鍋由北京長(zhǎng)安科學(xué)儀器廠生產(chǎn)。
1.3試驗(yàn)方法
1.3.1樣品處理將李果實(shí)隨機(jī)分成5組,分別采用1 mmol/L SA、47 μmol/L ABA 、5 mmol/L鎢酸鈉、1 mmol/L SA結(jié)合5 mmol/L鎢酸鈉、蒸餾水(對(duì)照組)浸泡30 min后,取出晾干。置于4 ℃下存放,每隔6 d取樣30個(gè)果,將果肉切成0.5 cm×0.5 cm×0.5 cm的小塊用液氮速凍后放置于-80 ℃下,用于記錄冷害發(fā)生情況及活性氧相關(guān)代謝指標(biāo)的測(cè)定。每個(gè)處理組150個(gè)果實(shí),重復(fù)3次。
1.3.2測(cè)定指標(biāo)和方法
1.3.2.1冷害指數(shù)的計(jì)算參照郭雨萱等[18]的方法。根據(jù)受冷害程度共分為5級(jí)。1級(jí),沒有冷害;2級(jí),輕微冷害:表面有直徑小于0.5 cm的凹陷或褐色斑點(diǎn);3級(jí),中度冷害:表面有直徑超過1 cm的褐色斑;4級(jí),嚴(yán)重冷害:褐變面積已超過果實(shí)的1/3;5級(jí),極其嚴(yán)重:果實(shí)已發(fā)生完全褐變。
冷害指數(shù)=(冷害級(jí)別數(shù)×發(fā)生冷害的果實(shí)個(gè)數(shù))
1.3.2.2總酚含量的測(cè)定根據(jù)曹建康等[19]的方法,略有改動(dòng)。提取緩沖液為1% HCl-甲醇溶液。以提取緩沖液為空白,測(cè)定提取液在280 nm下的吸光值,以每克李果實(shí)組織在波長(zhǎng)280 nm處的吸光值為總酚含量。
1.3.2.3過氧化物酶(POD)和多酚氧化酶(PPO)活性的測(cè)定根據(jù)曹建康等[19]的方法,略有改動(dòng)。POD活性測(cè)定:提取緩沖液為0.1 mol/L的檸檬酸-檸檬酸鈉,pH 6.6,其中含4% (質(zhì)量體積比) PVPP和1% Triton X-100。0.5 ml提取液,加3.0 ml 25 mmol/L 愈創(chuàng)木酚溶液和0.2 ml 0.5 mol/L H2O2溶液?jiǎn)?dòng)反應(yīng)。在反應(yīng)15 s時(shí)開始記錄反應(yīng)體系在470 nm處吸光值,5 min內(nèi)每隔30 s記錄1次。PPO活性測(cè)定:提取緩沖液為0.1 mol/L乙酸-乙酸鈉,pH 5.5,其中含1 mmol/L聚乙二醇6000,4% (質(zhì)量體積比) PVPP,1% Triton X-100。100 μl提取液,加4.0 ml 50 mmol/L(pH 5.5)的乙酸-乙酸鈉緩沖液和1.0 ml 50 mmol/L的鄰苯二酚溶液,在反應(yīng)15 s時(shí)開始記錄反應(yīng)體系在420 nm處吸光值,6 min內(nèi)每隔1 min記錄1次。
1.3.2.4活性氧代謝相關(guān)酶(SOD、CAT、APX)活性的測(cè)定根據(jù)曹建康等[19]的方法,略有改動(dòng)。SOD活性測(cè)定:采用氮藍(lán)四唑比色法。提取緩沖液為0.1 mol/L磷酸緩沖液,pH 7.8,其中含5 mmol/L DTT二硫蘇糖醇和5% PVP。0.1 ml提取液,加入1.7 ml 50 mmol/L(pH 7.8)磷酸緩沖液,0.3 ml 13 mmol/L甲硫氨酸(MET),0.3 ml 75 μmol/L氮藍(lán)四唑(NBT),0.3 ml 10 μmmol/L EDTA-Na2和2 μmol/L核黃素,光照25 min后,記錄在560 nm下的吸光值。CAT活性測(cè)定:提取緩沖液為0.2 mol/L(pH 7.9)磷酸鈉緩沖液,其中含133 L Triton-X100。吸取20 μl提取液,加1.5 ml(pH 7.9)磷酸鈉緩沖液,再加1 ml蒸餾水,30 ℃靜置30 min。測(cè)定時(shí)再加入0.3 ml 0.1 mol/L H2O2,在反應(yīng)15 s時(shí)開始記錄反應(yīng)體系在240 nm處吸光值,5 min內(nèi)每隔30 s記錄1次。APX活性測(cè)定:提取緩沖液為0.1 mol/L Tris-HCl(pH 8.0),其中含0.1 mmol/L EDTA,1.0 mmol/L抗壞血酸和2% PVPP。0.1 ml提取液,加入2.6 ml反應(yīng)緩沖液和0.3 ml 2 mol/L的H2O2溶液,5 min內(nèi)每隔30 s記錄1次在290 nm下的吸光值。
1.4數(shù)據(jù)處理
應(yīng)用Excel 2007統(tǒng)計(jì)分析所有數(shù)據(jù),計(jì)算標(biāo)準(zhǔn)偏差并制圖。應(yīng)用SPSS16.0軟件對(duì)所有數(shù)據(jù)進(jìn)行方差分析和差異顯著性分析。P<0.05表示差異顯著,P<0.01表示差異極顯著。
2結(jié)果與分析
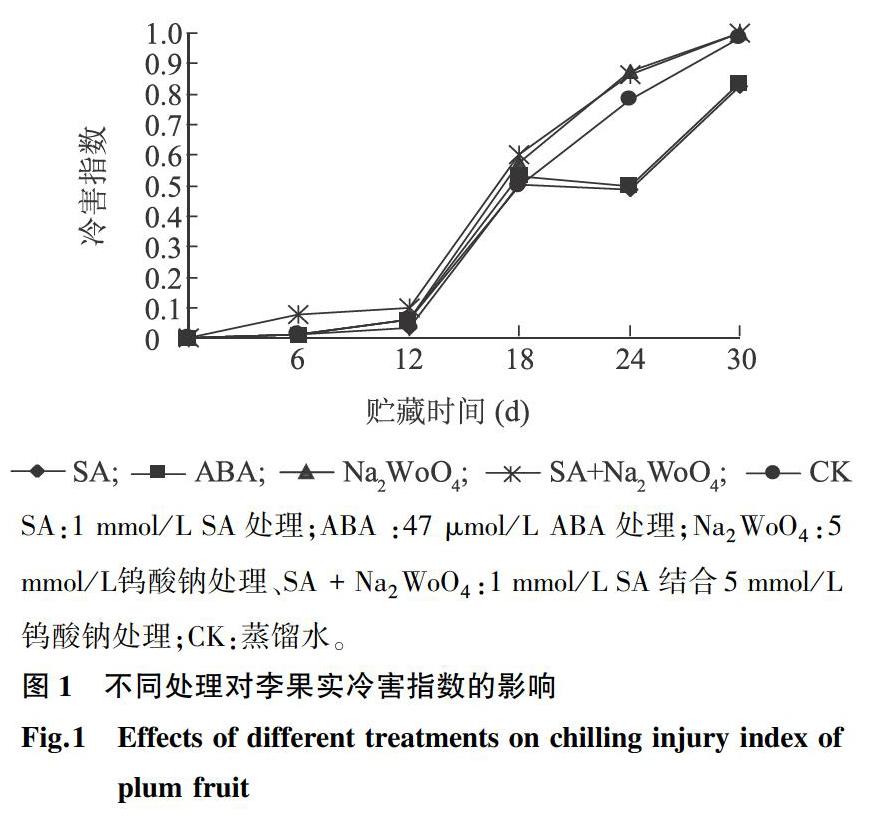
2.1不同處理對(duì)李果實(shí)冷害指數(shù)的影響
由圖1可以看出,隨著貯藏時(shí)間延長(zhǎng),各處理組果實(shí)冷害程度不斷增加。其中ABA處理組冷害指數(shù)上升速度低于對(duì)照組(P<0.05),Na2WoO4和SA結(jié)合Na2WoO4處理組上升速度高于對(duì)照組(P<0.05)。18 d以后,SA和ABA處理組冷害指數(shù)顯著低于對(duì)照組(P<0.05),而Na2WoO4和SA結(jié)合Na2WoO4處理組顯著高于對(duì)照組(P<0.05),且均在30 d達(dá)到最大值。表明,SA和ABA能夠顯著抑制果實(shí)采后冷害的發(fā)生,ABA合成抑制劑Na2WoO4降低了果實(shí)的抗冷能力,且該作用不受SA的影響。
2.2不同處理對(duì)李果實(shí)總酚含量的影響
酚類物質(zhì)主要與果實(shí)的色澤發(fā)育、成熟衰老、組織褐變、抗逆性和抗病性有關(guān)。如圖2所示,隨著貯藏時(shí)間延長(zhǎng),對(duì)照組總酚含量呈現(xiàn)先升高后下降的趨勢(shì)。SA和ABA處理組的上升速度顯著高于對(duì)照組(P<0.05),Na2WoO4和SA結(jié)合Na2WoO4處理組的上升速度顯著低于對(duì)照組(P<0.05)。SA和ABA處理組的總酚含量分別在第24 d和30 d達(dá)到最大值,且顯著高于對(duì)照組(P<0.05)。Na2WoO4和SA結(jié)合Na2WoO4處理組的總酚含量始終顯著低于對(duì)照組(P<0.05)。表明,SA和ABA提高酚類物質(zhì)含量可能與提高果實(shí)抗病能力有關(guān),ABA合成抑制劑Na2WoO4顯著抑制了ABA這一能力,且該作用不受SA的影響。
2.3不同處理對(duì)李果實(shí)POD和PPO活性的影響
PPO和POD可促使果實(shí)內(nèi)酚類物質(zhì)氧化發(fā)生褐變,褐變是果實(shí)發(fā)生冷害的首要現(xiàn)象,導(dǎo)致果實(shí)品質(zhì)下降[20]。如圖3所示,隨著貯藏時(shí)間延長(zhǎng),對(duì)照組的POD活性呈現(xiàn)上升趨勢(shì),并在30 d達(dá)到最大值。在貯藏期間的第12 d、第24 d、第30 d,SA和ABA處理組的上升趨勢(shì)顯著低于對(duì)照組(P<0.05),且ABA處理組在整個(gè)貯藏期間始終維持較低水平。而Na2WoO4和SA結(jié)合Na2WoO4處理組上升趨勢(shì)顯著高于對(duì)照組(P<0.05)。第12 d、第30 d時(shí),SA處理組的POD活性顯著低于對(duì)照組(P<0.05),而Na2WoO4和SA結(jié)合Na2WoO4處理組的POD活性在第30 d時(shí)迅速上升達(dá)到最大值,顯著高于對(duì)照組(P<0.05)。如圖4所示,貯藏期間PPO上升趨勢(shì)與POD上升趨勢(shì)一致。表明,SA和ABA可以延緩果實(shí)POD和PPO活性的上升,抑制果實(shí)氧化褐變,延緩果實(shí)冷害。ABA合成抑制劑Na2WoO4加劇果實(shí)氧化褐變,該作用不受SA的影響。
2.4不同處理對(duì)李果實(shí)SOD、CAT和APX活性的影響
SOD、CAT和APX作為活性氧清除酶,可以有效地減少活性氧積累,減輕細(xì)胞膜傷害[21]。如圖5所示,對(duì)照組SOD活性呈現(xiàn)先升高后下降的趨勢(shì),在第6 d達(dá)到最大值。SA和ABA處理組的上升速度顯著高于對(duì)照組(P<0.05),且在第18 d達(dá)到最大值后緩慢下降。第6 d后,Na2WoO4和SA結(jié)合Na2WoO4處理組整體呈現(xiàn)下降趨勢(shì)。SA結(jié)合Na2WoO4處理組雖在第18 d SOD活性升高,但在第24 d后活性迅速下降,顯著低于對(duì)照組(P<0.05)。SA和ABA處理組的SOD活性在第18 d后顯著高于對(duì)照組(P<0.05),Na2WoO4處理組在第12 d后SOD活性與對(duì)照組無顯著性差別,SA結(jié)合Na2WoO4處理組在第6~12 d與對(duì)照無顯著性差別,在第24~30 d顯著低于對(duì)照組(P<0.05)。如圖6所示,CAT活性變化趨勢(shì)與SOD活性變化趨勢(shì)一致,但Na2WoO4處理組的上升速度略高于對(duì)照組。如圖7所示,APX活性變化趨勢(shì)與SOD活性變化趨勢(shì)一致。在第24 d后,SA和ABA處理組的APX活性顯著高于對(duì)照組(P<0.05),Na2WoO4和SA結(jié)合Na2WoO4處理組的APX活性顯著低于對(duì)照組(P<0.05)。表明,SA和ABA能夠顯著提高果實(shí)對(duì)活性氧的清除能力,而ABA合成抑制劑Na2WoO4顯著抑制了活性氧清除酶的活性,且該作用不受SA的影響。
3討論
果蔬在正常條件下,其活性氧產(chǎn)生能力與消除能力相抵消,系統(tǒng)穩(wěn)定。但在不適低溫條件下,就會(huì)造成活性氧代謝紊亂,膜脂過氧化造成細(xì)胞膜的損傷,最終導(dǎo)致冷害的發(fā)生。從生理的角度看,低溫影響抗氧化酶的活性,所以一般以抗氧化酶作為研究果蔬冷害的對(duì)象[22],抗氧化酶主要有SOD、POD、CAT等[23-24]。SA可以減輕膜脂過氧化,提高抗氧化酶活性,加強(qiáng)活性氧代謝途徑來減輕低溫傷害。ABA可以提高果蔬抵御逆境脅迫的能力。ABA處理甘蔗的研究發(fā)現(xiàn)施用外源脫落酸可以誘發(fā)內(nèi)源脫落酸的增加,有效提高其抗冷能力[25]。徐文玲等[26]發(fā)現(xiàn)外源ABA可以有效提高大白菜相關(guān)抗氧化酶的活性。孔祥佳[27]通過熱處理對(duì)橄欖葉抗冷性的研究指出,降低果實(shí)內(nèi)POD和PPO活性可以有效地延緩果實(shí)內(nèi)酚類物質(zhì)的下降,并且通過保持其細(xì)胞結(jié)構(gòu)的完整,抑制果實(shí)POD、PPO與果實(shí)酚類物質(zhì)接觸,從而延緩果實(shí)褐變,減輕冷害的發(fā)生。酚類物質(zhì)不但可以提高生物對(duì)逆境脅迫的抵御能力,而且具有很強(qiáng)的抗氧化能力[28]。李文娟等[29]在施用外源氯化鉀對(duì)玉米的研究中指出,通過對(duì)酚類物質(zhì)代謝的調(diào)節(jié),還可以提高其抗病性。本研究希望通過SA、ABA等外源小分子物質(zhì)處理李果實(shí),通過提高其總酚含量及氧化酶活性,進(jìn)而提高植物的抗冷性。
本研究結(jié)果表明,經(jīng)過SA、ABA處理后的李果實(shí)可以降低冷害指數(shù),說明SA和ABA可以延緩冷害的發(fā)生,降低冷害的損傷速度。SA、ABA處理提高了李果實(shí)的總酚含量并推遲了POD、PPO活性的升高,不但提高了果實(shí)的抗病性,還延緩其褐變和冷害的發(fā)生。而活性氧代謝相關(guān)酶SOD、CAT、APX活性顯著提高,抑制了膜脂過氧化的發(fā)生,維持活性氧代謝平衡。這與前人在杏果實(shí)[30]、藍(lán)莓果實(shí)[31]、獼猴桃果實(shí)[32]的研究結(jié)果一致。
參考文獻(xiàn):
[1]張銀志,孫秀蘭,劉興華,等. 低溫脅迫和變溫處理對(duì)李子生理特性的影響[J].食品科學(xué),2003(2):134-138.
[2]范小玉,張顯. 油菜素內(nèi)酯對(duì)低溫弱光脅迫下西瓜幼苗耐冷性的影響[J].北方園藝,2012(7):5-8.
[3]廖金柯,趙克,胡小燕,等. 外源鈣對(duì)棉花幼苗抗冷性的影響[J].新疆農(nóng)業(yè)科學(xué),2012,49(4):687-693.
[4]楊美森,王雅芳,干秀霞,等.外源-氧化氮對(duì)冷害脅迫下棉花幼苗生長(zhǎng)、抗氧化系統(tǒng)和光合特性的影響[J].中國(guó)農(nóng)業(yè)科學(xué),2012,45(15):3058-3067.
[5]黃亞成,秦云霞.植物中活性氧的研究進(jìn)展[J].中國(guó)農(nóng)學(xué)通報(bào),2012,28(36):219-226.
[6]趙麗英,鄧西平,山侖.活性氧清除系統(tǒng)對(duì)干旱脅迫的響應(yīng)機(jī)制[J]. 西北植物學(xué)報(bào),2005, 25(2): 413-418
[7]GUAN N, BLOMSMA S A, FAHY G M L. Analysis of gene expression changes to elucidate the mechanism of chilling injury in precision-cut liver slices[J].Toxicology in Vitro, 2013, 27(2): 890-899.
[8]侯媛媛,朱璇,王英,等. 水楊酸處理對(duì)杏果實(shí)冷害及活性氧代謝的影響[J].食品科學(xué),2014,35(4):195-199.
[9]AGHDAM M S,ASGHARI M R,MORADBEYGI H,et al. Effect of postharvest salicylic acid treatment on reducing chilling injury in tomato fruit[J].Romanian Biotechnological Letters,2012, 17(4):7466-7473.
[10]張紅宇,尹京苑,萬嗣寶. 水楊酸和茉莉酸甲酯處理對(duì)水蜜桃低溫貯藏冷害的影響[J].食品工業(yè),2012,33(10):88-91.
[11]姚俠妹,紀(jì)敬,岳劍云,等. ABA對(duì)鹽脅迫下側(cè)柏活性氧代謝及其相關(guān)基因表達(dá)的研究[J].西北植物學(xué)報(bào),2017,37(1):105-114.
[12]GR AHAM D,PATTER SON B D. Responses of plants to low,nonfreezing temperatures: proteins,metabolism,and acclimation[J]Annual Reviews of Plant Physiology,2003,33: 347-372.
[13]WASILEWSKA,VLAD,SIRICHANDRA,et al. An update on abscisic acid signaling in plants and more[J].Molecular Plant,2008,1(2):198-217.
[14]CHEN J,MAO L,MI H,et al. Involvement of abscisic acid inpostharvest water-deficit stress associated with the accumulation of anthocyanins in strawberry fruit[J]. Postharvest Biology & Technology,2016,111( 31):99-105.
[15]WANG Y, LU W J,ZHANG Z Q,et al. ABA and putrescine treatments alleviate chilling injury in banana fruits during storage at 8 ℃[J]. Acta Photophysiologica Sinica,2003,29(6):549-554.
[16]ZHANG Q T, ZHANG L L, GENG B,et al.Interactive effects of abscisic acid and nitric oxide on chilling resistance and active oxygen metabolism in peach fruit during cold storage[J].Journal of the Science of Food and Agriculture,2019, 99(7):3367-3380.
[17]黃杏,梁勇生,楊麗濤,等. 低溫脅迫下脫落酸及合成抑制劑對(duì)甘蔗幼苗抗氧化系統(tǒng)的影響[J].華南農(nóng)業(yè)大學(xué)學(xué)報(bào),2013,34(3):356-361.
[18]郭雨萱,郝利平,盧銀潔,等. 不同貯藏溫度對(duì)茄子冷害發(fā)生及質(zhì)構(gòu)特性的影響[J].核農(nóng)學(xué)報(bào),2016,30(9):1763-1769.
[19]曹建康,姜微波,趙玉梅. 果蔬采后生理生化實(shí)驗(yàn)指導(dǎo)[M].北京:中國(guó)輕工業(yè)出版社,2007.
[20]趙迎麗,李建華,閆根柱,等. 不同貯藏溫度下石榴褐變生理特性的研究[J].食品研究與開發(fā),2013,34(18):90-93.
[21]YOO Y K. Effects of acetylsalicylic acid and calcium chloride on photosynthetic apparatus and reactive oxygen-scavenging enzymes in chrysanthemum under low temperature stress with low light[J].Agricultural Sciences in China,2010,9(12):1777-1786.
[22]趙黎明,李明,鄭殿峰,等. 冷害后植物生理變化及外源物質(zhì)調(diào)控研究進(jìn)展[J].中國(guó)農(nóng)學(xué)通報(bào),2015,31(12):217-223.
[23]李雅潔,張其安,陸曉民. 不同外源物質(zhì)對(duì)低溫弱光次生鹽漬化復(fù)合逆境下黃瓜幼苗生長(zhǎng)、抗氧化系統(tǒng)及光合作用的影響[J].江蘇農(nóng)業(yè)學(xué)報(bào), 2018, 34(2):404-410.
[24]趙曾菁,宋奇琦,趙虎,等. 模擬霜凍條件對(duì)黃瓜幼苗生長(zhǎng)及抗氧化酶活性的影響[J].南方農(nóng)業(yè)學(xué)報(bào),2018,49(4):757-762.
[25]黃杏,楊麗濤,張保青,等. 甘蔗脫落酸脅迫成熟誘導(dǎo)蛋白基因(SoASR)的克隆和表達(dá)分析[J].生物技術(shù)通報(bào),2013(2):93-99.
[26]徐文玲,王翠花,牟晉華,等. 不同濃度脫落酸對(duì)大白菜抗冷特性的影響[J].山東農(nóng)業(yè)科學(xué),2012,44(1):47-50.
[27]孔祥佳. 熱空氣處理誘導(dǎo)采后橄欖果實(shí)抗冷性機(jī)理的研究[D].福州:福建農(nóng)林大學(xué),2011.
[28]周白雪,謝君,包垠秋,等. 低溫貯藏對(duì)蓮藕酚類物質(zhì)組成的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2019,47(1):189-191.
[29]李文娟,何萍,金繼運(yùn). 氯化鉀對(duì)玉米莖腐病抗性反應(yīng)中酚類物質(zhì)代謝的影響[J].植物營(yíng)養(yǎng)與肥料學(xué)報(bào),2008,14(3):508-514.
[30]袁潔. 水楊酸和熱處理對(duì)杏抗冷性及熱激蛋白產(chǎn)生影響的研究[D].烏魯木齊:新疆農(nóng)業(yè)大學(xué),2014.
[31]曲文穎,劉真真,謝琳淼,等. 外源脫落酸和乙烯利對(duì)藍(lán)莓重要品質(zhì)的調(diào)控[J].江蘇農(nóng)業(yè)科學(xué),2017,45(17):126-129.
[32]孫波,焉志遠(yuǎn),佟斌. 水楊酸處理對(duì)軟棗獼猴桃冷害性狀的影響[J].經(jīng)濟(jì)林研究,2017,35(4):197-201,206.
(責(zé)任編輯:陳海霞)
