兩株蘇云金芽胞桿菌晶體與芽胞形成過程的細胞學對比分析
2020-05-28 07:40:14全梅芳彭金莉何昊城夏立秋丁學知
激光生物學報 2020年2期
全梅芳,彭金莉,何昊城, 夏立秋, 丁學知*
(1.湖南師范大學醫學院, 長沙 410013; 2.微生物分子生物學湖南省重點實驗室, 淡水魚類發育生物學國家重點實驗室, 湖南師范大學生命科學學院, 長沙 410081)
蘇云金芽胞桿菌(Bacillusthuringiensis, Bt)是一種自然界廣泛分布的革蘭陽性細菌,其主要特征是在菌體形成芽胞的同時,伴隨有大量殺蟲晶體蛋白(insecticidal crystal proteins,ICPs)產生[1]。這種ICPs的高效表達源于它特殊的轉錄調控模式,且與芽胞形成過程緊密偶聯。芽胞形成主要通過順序激活6個RNA聚合酶 σ 因子,分別為:σA、σH、σF、σE、σG和σK。這些因子在菌體生長的不同階段分別發揮調控功能,指導芽胞形成相關基因有序而準確的表達[2]。
Bt的芽胞形成過程與枯草芽胞桿菌(Bacillussubtilis)極為相似,大致分為以下7個階段。第一階段(Stage Ⅰ),與營養體細胞在形態上沒有明顯差別;第二階段(Stage Ⅱ),軸絲和芽胞隔膜開始形成;第三階段(Stage Ⅲ),前芽胞內卷,游離在母細胞的胞質里,可觀察到伴胞晶體;第四階段(Stage Ⅳ),芽胞外壁逐漸形成;第五階段(Stage Ⅴ),芽胞內衣沉積形成;第六、七階段(Stage Ⅵ&Ⅶ),皮層形成,芽胞逐漸成熟,隨后,芽胞和伴胞晶體從裂解的母細胞中釋放。其中cry基因的表達調控主要是由σE和σK因子控制,ICPs出現最初始于Stage Ⅲ早期,在Stage Ⅳ至Ⅴ階段完成其形成過程[3,4]。
Bt 4.0718 菌株為本室選育的一株庫斯塔克亞種高毒力菌株,能產生橢圓形芽胞和典型的菱形與方形晶體,對棉鈴蟲、小菜蛾和甜菜夜蛾等農業害蟲表現出很強的毒殺活性[5,6]。Bt ΔleuB是以Bt 4.0718為出發菌株,對其3-異丙基蘋果酸脫氫酶(3-isopropylmalate dehydrogenase)基因leuB實現無痕敲除得到的一株工程菌。Bt ΔleuB在液體LB培養基中生長時無芽胞出現,只產生晶體,這種特殊表型打破了現有認識,即認為Bt的晶體必須伴隨芽胞而出現。
FM4-64染料是親脂性苯乙烯基化合物,能與質膜及細胞器膜結構特異結合后發出紅色熒光,可被應用于芽胞桿菌中芽胞形成及母細胞凋亡過程的研究[7-9]。本研究以Bt 4.0718 和Bt ΔleuB菌株為研究對象,采用FM4-64染料對不同生長階段的菌體進行染色,在激光共聚焦掃描顯微鏡(confocal laser scanning microscope,CLSM)下對比觀察兩株菌芽胞發育與晶體形成的過程,以期從細胞水平上為理解這一類特殊表型發生的機制提供線索。
1 材料與方法
1.1 菌株與培養基
Bt 4.0718菌株(BacillusthuringiensisCCTCC No.M200016)為本室選育,屬于庫斯塔克亞種。Bt ΔleuB(CCTCC No.M2019112)是以Bt 4.0718為出發菌株,leuB基因無痕敲除的工程菌。Bt菌株的培養采用LB培養基(NaCl 10 g,tryptone 10 g,yeast extract 5 g,pH7.0,加超純水定容至1 L,121 ℃滅菌20 min)。
1.2 主要試劑與儀器
FM4-64(T3166)購自Invitrogen公司。基因組DNA去除以及逆轉錄反應試劑盒HiScript?II Q RT SuperMix for qPCR(+gDNA wiper)購自Vazyme公司。實時熒光定量PCR反應(qRT-PCR)試劑盒PowerUPTM SYBRTM Green Master Mix購自ABI公司。正置顯微鏡購自德國卡爾-蔡司公司,型號為AXIO SCOPE A1;激光共聚焦掃描顯微鏡購自卡爾-蔡司,型號為LSM 710。
1.3 生長情況分析
將各菌株接種于10 mL LB 液體培養基過夜活化后,按照1%比例分別轉接于50 mL LB培養基繼續培養,每組設置3個平行。每2 h取樣,以新鮮LB作為空白對照,利用分光光度計測定OD600值,確定各菌株的生長曲線。
1.4 染色液配置
1 mg FM4-64 染料溶解于0.5 mL DMSO中配備成母液,再用Hank’s平衡鹽溶液(8 g/L NaCl,0.4 g/L KCl,1 g/L Glucose,0.06 g/L KH2PO4,0.126 g/L Na2HPO4·12H2O,0.35 g/L NaHCO3)稀釋成工作液(25 μg/mL)。
1.5 染色與顯微鏡觀察
染色:收集100 μL菌液,水洗3遍后重懸于200 μL無菌水中;取上述待染菌懸液10 μL,加入FM4-64工作液100 μL混勻,冰上染色1 min。
顯微鏡制樣:取染色后的菌懸液(或未染菌懸液)6 μL滴于潔凈載玻片上,蓋上蓋玻片、壓片,激光共聚焦掃描顯微鏡(或倒置顯微鏡)觀察。
1.6 總RNA提取與目的基因轉錄水平分析
活化過夜的Bt菌液按1%的接種量接種于30 mL液體LB培養基,30 ℃培養30 h,12 000 r/min離心3 min,收集0.1 g菌體;總RNA提取使用Trizol試劑;基因組DNA去除、逆轉錄反應及qRT-PCR反應按試劑盒說明書進行;引物序列及名稱見表1;各基因的mRNA相對表達水平以16s rRNA為內參進行歸一化處理;基因相對表達量計算按照2-ΔΔCT標準方法進行。

表1 本研究所用引物Tab.1 Primers used in this study
2 結果與分析
2.1 Bt 4.0718與Bt ΔleuB的生長情況分析
生長曲線測定結果顯示,在延滯期、對數生長期和穩定期前、中期,Bt 4.0718與Bt ΔleuB兩株菌的生長曲線沒有顯著差異,但從 26 h 開始(穩定期后期),Bt 4.0718由于菌體的迅速裂解,導致培養物的OD600值急劇下降,然而Bt ΔleuB的OD600值始終維持在比較平穩的狀態(圖1)。
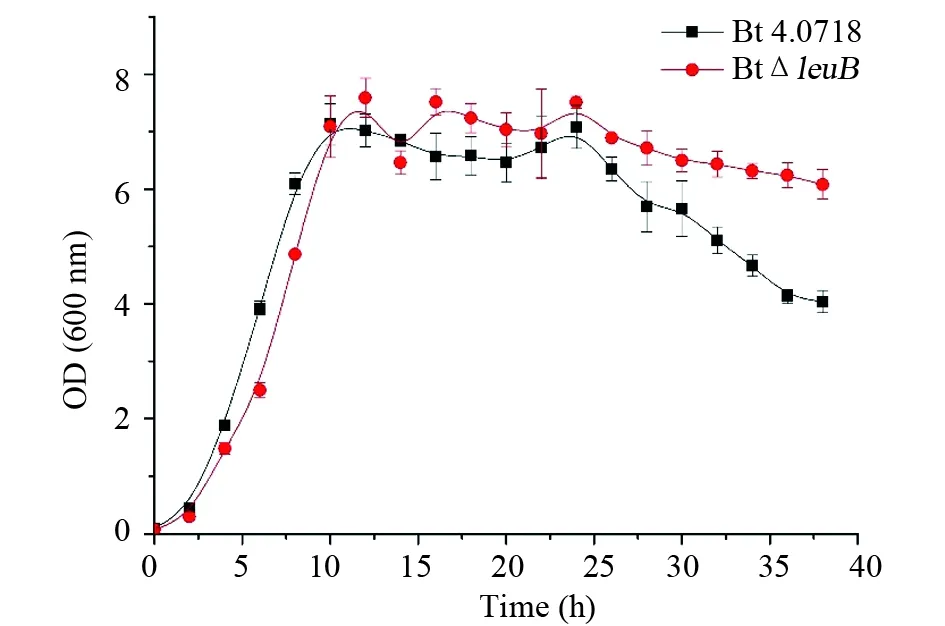
圖1 Bt 4.0718和Bt ΔleuB在LB培養基中的生長曲線比較Fig.1 Growth curves of Bt 4.0718 and Bt ΔleuB in LB medium
2.2 相差顯微鏡觀察
相差顯微鏡觀察結果顯示:在培養至24 h時(穩定期后期),Bt 4.0718 細胞內可見典型的卵圓形芽胞和菱形晶體;同時期的 Bt ΔleuB無芽胞,大部分菌體內可見規則的菱形晶體。Bt 4.0718菌體細胞在48 h 時已完全裂解,芽胞和晶體隨之釋放;而Bt ΔleuB培養至96 h 時仍未裂解,菌體內包裹的菱形晶體體積進一步增大,有的甚至占據母細胞體積的1/3(圖2)。
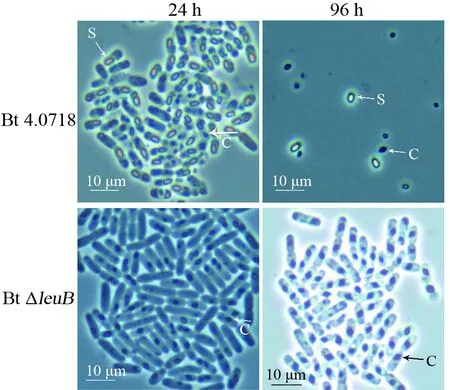
圖2 Bt 4.0718和Bt ΔleuB在LB培養基中相差顯微鏡觀察Fig.2 Observation of Bt 4.0718 and Bt ΔleuB cells in LB medium by phase-contrast microscopyS:芽胞;C:伴胞晶體S:Spores;C:Parasporal crystals
2.3 激光共聚焦掃描顯微鏡觀察
為了探索Bt ΔleuB特殊表型背后的生物學機制,分別將菌株Bt 4.0718和Bt ΔleuB培養至芽胞期,取不同生長時間點的樣品,用FM4-64染料對菌體染色,CLSM觀察對比分析兩株菌的芽胞發育過程。結果表明:當Bt 4.0718菌株生長到10 h時,少部分細胞開始形成不對稱隔膜;14 h時,大部分細胞進入前芽胞內吞階段;17 h時,芽胞內吞已完成,此時芽胞未能標記上FM4-64,只能觀察到活細胞的外部膜狀輪廓,相差模式下可見明顯的芽胞和晶體;24 h時,進入凋亡階段,母細胞膜破裂,FM4-64進入母細胞胞質溶膠,與聚集的膜碎片結合,導致細胞的外部輪廓不規則(圖3)。
樣品Bt ΔleuB的觀察結果顯示,14~17 h時,菌體逐漸形成不對稱隔膜,相差模式下17 h時細胞內已可見明顯的菱形晶體;24 h時,部分細胞進入胞吞階段,隔膜形成期細胞仍然可見,晶體體積進一步增大;31 h時的芽胞發育狀態與24 h無明顯差別(結果未顯示);40 h時,晶體體積達到最大,但仍無芽胞,母細胞尚無裂解跡象。然而CLSM觀察發現:紅色熒光在細胞內聚集,且顯示的細胞外部輪廓不規則,表明大部分母細胞的質膜可能已破裂(圖4)。
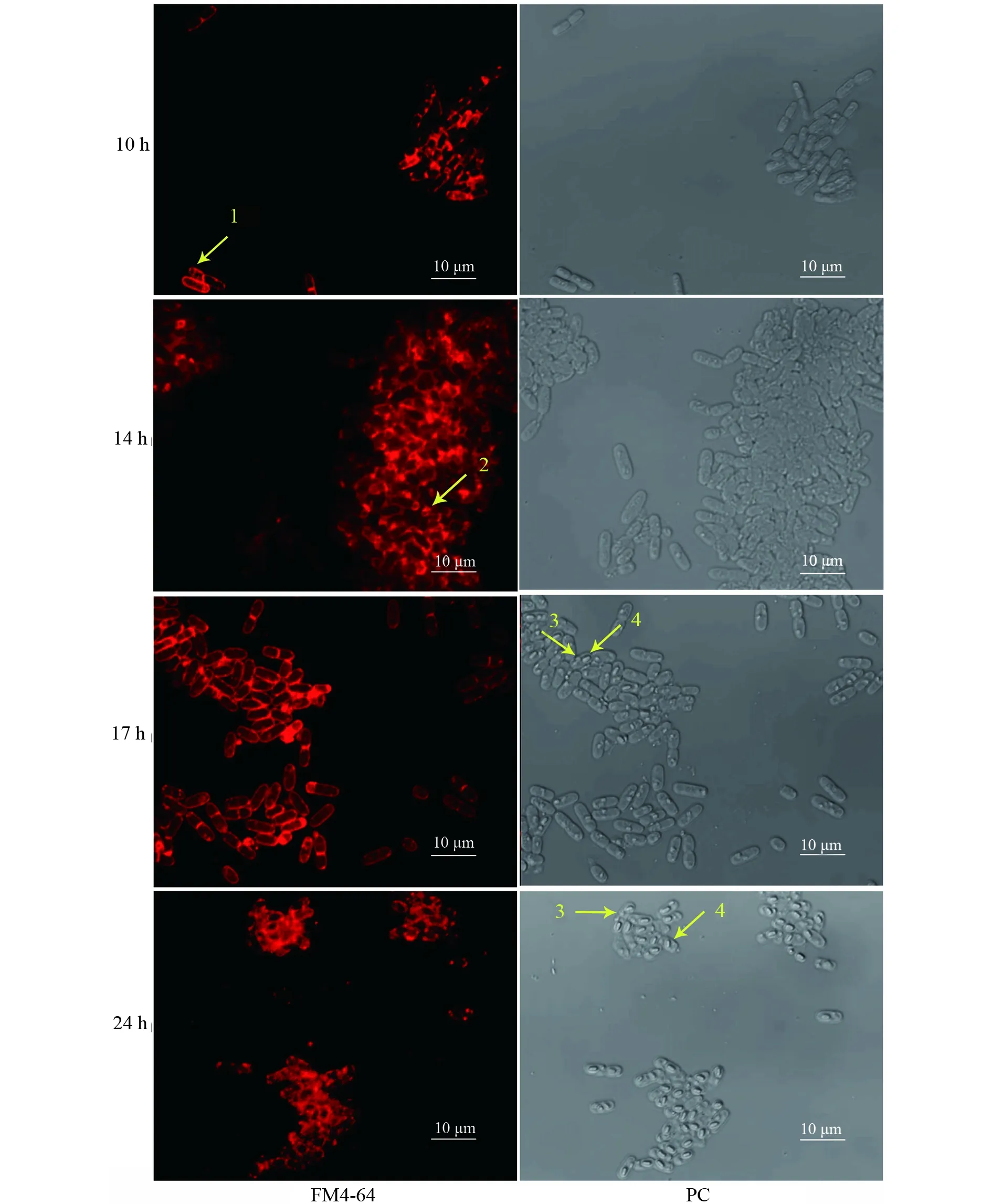
圖3 激光共聚焦掃描顯微鏡分析Bt 4.0718菌株的芽胞發育過程以及母細胞裂解情況Fig.3 Analysis of the spore development process and mother cell lysis in Bt 4.0718 by CLSM箭頭1:不對稱隔膜;箭頭2:前芽胞內吞;箭頭3:伴胞晶體;箭頭4:芽胞Arrow 1:The asymmetric septum;Arrow 2:The process of spore engulfment;Arrow 3:Parasporal crystal;Arrow 4:Spore
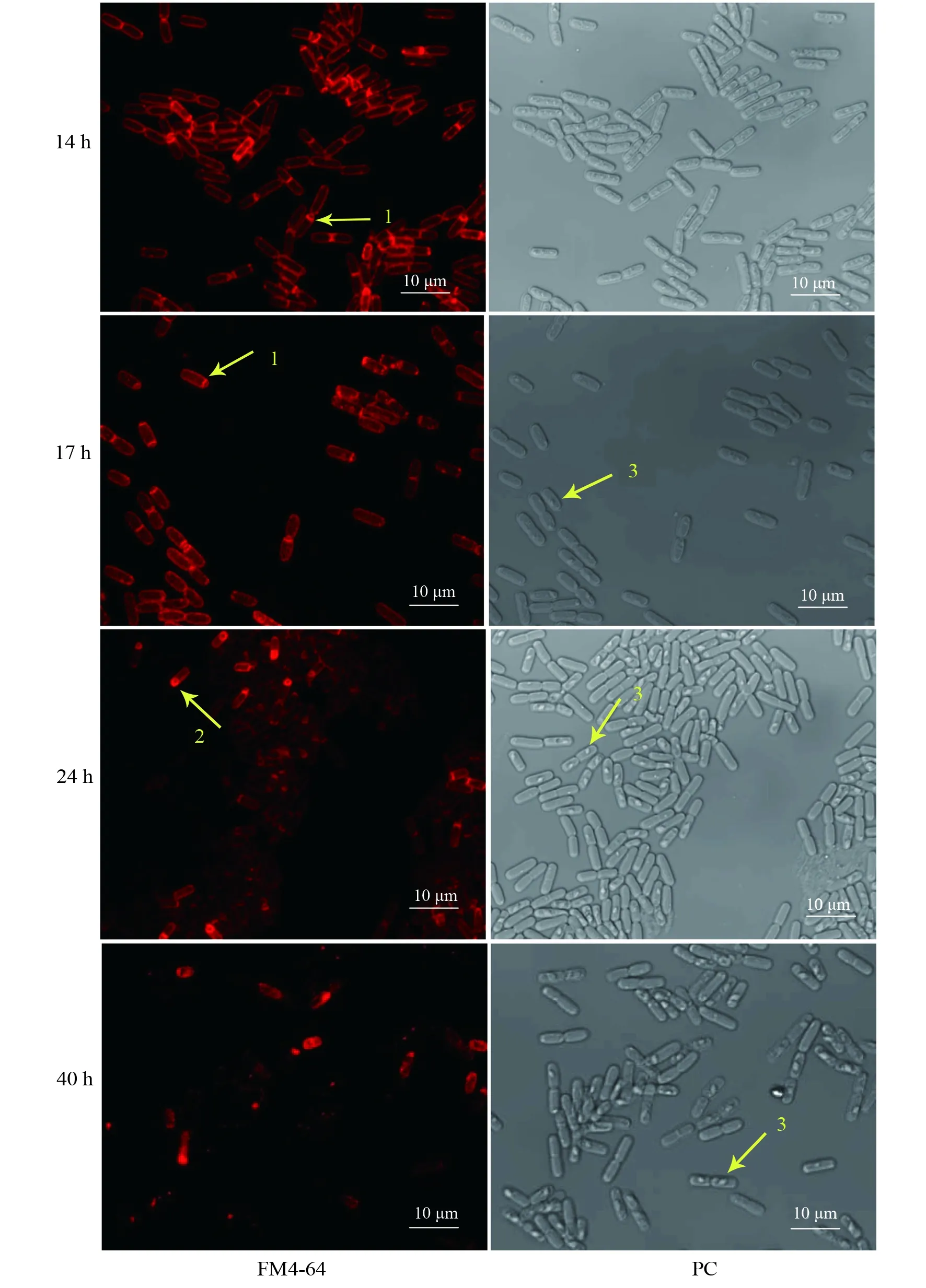
圖4 激光共聚焦掃描顯微鏡分析Bt ΔleuB菌株的芽胞發育過程以及母細胞裂解情況Fig.4 Analysis of the spore development process and mother cell lysis in Bt ΔleuB by CLSM箭頭1:不對稱隔膜;箭頭2:前芽胞內吞;箭頭3:伴胞晶體Arrow 1:The asymmetric septum;Arrow 2:The process of spore engulfment;Arrow 3:Parasporal crystal
2.4 σE、spoIIR和spoIIGA的轉錄水平分析
晶體蛋白形成依賴于芽胞形成,因為cry基因的表達調控主要受σE和σK因子控制,它們分別在芽胞形成早期和晚期的母細胞中表達。鑒于Bt ΔleuB菌株的特殊表型,推測其細胞內維持ICPs正常表達的主要為σE。qRT-PCR結果顯示,相對于Bt 4.0718,Bt ΔleuB中σE和上游信號分子spoIIR以及負責pro-σE切割活化的蛋白酶spoIIGA的轉錄水平均上升。
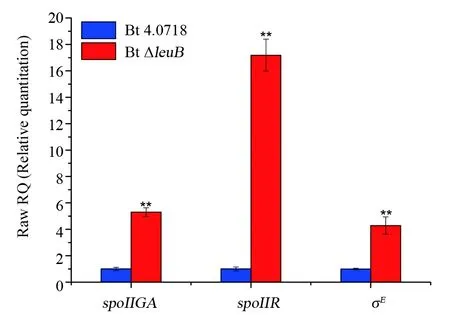
圖5 兩株Bt菌株中spoIIGA、spoIIR和σE基因的相對轉錄水平分析。** :P≤0.01 vs Bt 4.0718Fig.5 The relative transcriptional levels of the spoIIGA,spoIIR and σE genes in the two Bt strains. ** :P≤0.01 vs Bt 4.0718
3 討論
在生長穩定期后期,Bt細胞會逐漸區室化形成一個卵圓形的芽胞,同時在母細胞內會形成伴胞晶體,這也是Bt區別于蠟狀芽胞桿菌(Bacilluscereus)和炭疽芽胞桿菌(Bucilusanthracis)的顯著特征,而只產生芽胞或晶體的突變菌株是研究Bt芽胞與晶體關系的重要遺傳材料。無晶體突變株的構建方法已非常成熟,編碼晶體蛋白的基因大多數位于Bt質粒上,因此多采用質粒消除的方法得到無晶體突變株[10]。只產生晶體的無芽胞突變菌株的構建原理則相對較復雜,這一類工程菌基本上是芽胞形成關鍵調控因子如spo0A、σE、spoIIID或σK因子的基因缺陷型突變株。其中spo0A或σE因子突變將嚴重影響芽胞形成依賴型cry基因的轉錄。Lereclus等[11]將cry3A或受cry3A啟動子控制的cry1基因轉入spo0A突變株后,能過量表達晶體蛋白且母細胞不裂解,晶體被包裹在細胞內,對解決晶體蛋白的紫外穩定性差等問題具有潛在應用價值[11]。spoIIID或σk基因缺失突變株基本喪失芽胞形成能力,但依然形成晶體[12,13]。與之相似的是,本研究中的Bt ΔleuB菌株也不能正常產生芽胞,但能形成伴胞晶體。已知Bt ΔleuB中的晶體蛋白編碼基因為典型的芽胞依賴型cry基因,因此推斷Bt ΔleuB菌株在生長過程中應該能夠進入芽胞期,但沒能順利完成芽胞發育過程,而是被阻斷于中間某一階段。
芽胞形成以不對稱細胞分裂開始,原來的細胞被雙層膜結構的隔膜分成了大小不等的兩個區室,即前芽胞(較小的區室)和母細胞。在不對稱分裂之后,芽胞形成的下一個形態學階段是母細胞吞噬前芽胞,使前芽胞成為其細胞質中的游離原生質體。接下來位于芽胞核心周圍的一系列保護結構被逐漸組裝,如皮質層、芽胞衣結構等[2,14,15],最后母細胞經由程序性細胞死亡機制裂解后,成熟的芽胞得以釋放,其中前芽胞內吞的完成是控制芽胞后期發育成熟的關鍵事件。CLSM觀察結果顯示:Bt 4.0718和Bt ΔleuB分別于10 h、14 h左右時開始形成不對稱隔膜(Stage Ⅱ);并分別于14 h、24 h左右時開始進入前芽胞內吞階段(Stage Ⅲ),這表明突變株能順利進入芽胞形成期,然而相比野生菌Bt 4.0718能完成芽胞發育的整個過程,Bt ΔleuB的芽胞發育可能被阻斷于內吞階段,因此導致該菌株無芽胞產生,但伴胞晶的體形成從細胞不對稱分裂期就已開始,且晶體體積較大。
Bt在芽胞發育過程中通過先后激活6個不同的σ因子,使調控芽胞形成基因準確無誤地表達。其中σF和σE分別在芽胞形成早期的前芽胞和母細胞中表達,σG和σK分別在芽胞形成晚期的前芽胞和母細胞中表達。細胞完成不對稱分裂后不久,激活的σF致使前芽胞分泌信號蛋白spoIIR進入母細胞,一旦接收到來自前芽胞的信號,定位于膜上的pro-σE即被天冬氨酸蛋白酶spoIIGA切割為活化的σE,活化的σE啟動母細胞中cry基因的轉錄[16,17]。qRT-PCR結果顯示,Bt ΔleuB中σE、spoIIR和spoIIGA的轉錄水平均顯著上調。因此,雖然Bt ΔleuB菌株芽胞發育缺陷(被阻滯于胞吞階段),但晶體蛋白積累并未受到影響,這再次印證了Bravo等[12]的研究結論,單獨的σE因子就足以支撐σE、σK依賴型雙重疊啟動子(Bt Ⅰ+ Bt Ⅱ)cry基因的高水平表達。另一方面,Bt ΔleuB菌株母細胞不裂解的表型與其芽胞發育缺陷密切相關,中國農科院張杰研究員等[18]研究發現,細胞壁水解酶基因cwlC的轉錄受σK控制,并且cwlC缺失可完全阻斷Bt母細胞的裂解。因此我們推測,Bt ΔleuB菌株中晚期sigma因子σK活性降低或缺失是其母細胞裂解受阻的關鍵因素。比較有趣的是,與Hosoya等[7]的結果一致,CLSM觀察結果發現,24 h時,Bt 4.0718細胞在相差模式下還有完整的細胞形態,但FM4-64染色結果顯示細胞膜已破裂,這表明在Bt細胞程序性死亡過程中,細胞膜破裂是早于細胞壁裂解的生物學事件。并且在Bt ΔleuB菌株中,雖然細胞壁裂解受阻斷,但在40 h時,細胞已開啟死亡程序(膜破裂)。
選擇一株具有特殊表型的Bt突變株及其出發菌株作為研究對象,利用熒光染料進行細胞發育過程的形態學觀察,為研究芽胞發育、晶體形成以及母細胞裂解三者之間的關系提供了一些線索,但具體的分子機制還有待進一步深入研究。
