斑馬魚miR-196a-1基因敲除品系的構建
2020-05-28 07:40:16付貴芳謝繽靈謝華平印遇龍
激光生物學報 2020年2期
曾 婷,付貴芳,謝繽靈,杜 涵,謝華平,3*,印遇龍,3,4
(1.動物腸道功能調控湖南省重點實驗室, 長沙 410081; 2.湖南師范大學生命科學學院, 動物營養與人體健康實驗室, 長沙 410081; 3.淡水魚類發育生物學國家重點實驗室, 長沙 410081; 4.中國科學院亞熱帶農業生態研究所, 中國科學院亞熱帶農業生態過程重點實驗室, 長沙 410125)
斑馬魚(Daniorerio),俗稱“zebrafish”,屬硬骨魚類,原產印度和孟加拉,為一種性情溫和、雜食性的熱帶淡水魚[1,2]。斑馬魚飼養成本低、繁殖力強、易大規模飼養,3個月可性成熟、胚胎易得、便于觀察,且斑馬魚和人類基因有高達87%的同源性,這些優點使斑馬魚成為研究人體疾病和發育機理的理想模式生物[3,4]。
miRNA是一類內生的、長度為19~23個堿基的非編碼RNA。一個miRNA可以調節多個基因[5],也可以通過幾個miRNA的組合來精細調控某個基因的表達[6]。miRNA存在多種形式,其被RNA聚合酶II轉錄為有帽子結構的和多聚腺苷酸尾巴(AAAAA)的初級轉錄本(pri-miRNA),該轉錄本可以是蛋白質編碼的,也可以是非蛋白質編碼的。初級轉錄物在核糖核酸酶Drosha和輔助因子Pasha的共同作用下切割產生約70個堿基的莖環前體miRNA(pre-miRNA),再被細胞質核糖核酸酶Dicer切割生成成熟的miRNA和反義miRNA(miRNA*)[7]。成熟的miRNA被整合到RNA誘導的沉默復合物(RNA-induced silencing complex,RISC)中,該復合物通過與miRNA的堿基配對識別靶mRNA,最常見的是抑制靶mRNA的翻譯或使其翻譯不穩定[8]。
miRNA最初被發現是在線蟲中參與lin-4的發育,而在哺乳動物中miRNA參與造血譜系分化和同源框基因調控[9,10]。隨后Muy等[11]研究發現microRNA表達水平的紊亂與腫瘤的發生、發展密切相關。Zhang等[12]的報道表明miR-196a在宮頸癌細胞中靶向調控netrin4基因,通過靶向同源基因團來調節哺乳動物的發育[13,14];miR-196a在胃癌的預后過程中表達上升[15];還有研究表明,miR-196a在食管鱗癌細胞中表達顯著上調,同時,在食管鱗癌患者唾液中miR-196a的表達比正常人高27倍,因此,miR-196a可能作為潛在的食管鱗狀細胞癌診斷標志[16]。在非洲爪蟾中,miR-196a的劑量不同會導致特定的眼部發育異常[17]。通過生物信息學分析比對發現miR-196a-1基因在各個物種中十分保守,說明miR-196a-1基因在調控生物體的發育、腫瘤發生和發展中起著非常重要的作用。
CRISPR/Cas9基因編輯技術是近年來火熱的基因編輯技術。CRISPR/Cas9基因編輯技術由兩部分組成,CRISPR結構骨架序列和Cas9蛋白。CRISPR結構骨架序列由前導序列、Cas基因、CRISPR基因座共同組成,具有招募Cas蛋白作用[18]。Cas9蛋白具有能特異性切割DNA雙鏈的作用。CRISPR基因座經過轉錄加工后,得到小片段的CRISPR RNA即crRNA,而crRNA與反式激活的crRNA即tracrRNA能形成一種嵌合RNA,該嵌合RNA與Cas9蛋白形成復合物,從而切斷基因組DNA[19,20]。后來科學家進一步改造,將crRNA和tracrRNA改造成一個向導RNA(sequence-specific guide RNA,sgRNA)[21]。目的基因靶位置序列的3’端有原間隔序列相鄰基序PAM(protospacer adjacent motifs),為5’-NGG-3’序列,這是Cas9蛋白的識別位點。最后,通過改變sgRNA序列,體外合成sgRNA,Cas9蛋白可以靶向切割目的基因,從而達到基因敲除的目的[22,23]。
目前,miRNA調控生物發育、腫瘤發生和發展機制越來越受到人們的關注,但是,許多miRNA的功能仍然未知。為了研究miR-196a-1基因在斑馬魚腸道發育過程中的作用,本文利用CRISPR/Cas9基因編輯技術構建斑馬魚miR-196a-1基因敲除品系,為探究miR-196a-1在腸道發育中的作用奠定了基礎。
1 材料與方法
1.1 試驗材料
1.1.1 試驗動物
本文用于試驗的TU品系斑馬魚來自本實驗室養殖。水溫28 ℃,pH值在6.5~7.5,14 h光照/10 h黑暗交替循環。1對斑馬魚1周產1次卵,1次產卵約200個胚胎,胚胎在恒溫28.5 ℃的E3水中培養,胚胎2 d后破膜,第5天開始喂食草履蟲至2周左右,之后轉移至養殖系統架上喂食豐年蟲,養殖系統上的斑馬魚1天喂食2次,早晚各1次。本文試驗一般在斑馬魚胚胎發育至1細胞期時進行顯微注射。
1.1.2 主要試劑
引物由擎科生物合成,PCR高保真酶購自擎科生物,DNA marker購自TAKARA公司,瓊脂糖購自BBI公司,PCR產物純化試劑盒購自生工生物,體外轉錄試劑盒購自Thermo Fisher Scientific公司,RNA純化試劑盒購自Qiagen公司,Cas9蛋白購自Thermo Fisher Scientific公司。
1.2 試驗方法
1.2.1 sgRNA靶位點設計
負反轉構造的存在說明研究區構造演化經歷了由早期擠壓應力到晚期拉張應力的轉換。早期擠壓形成的北西向構造帶在晚期拉張應力作用下使研究區具備了良好的油氣成藏條件。
首先在Ensembl網站上(http://www.ensembl.org/index.html)獲取目標基因的完整序列,各個轉錄本以及內外顯子等信息。然后找出所有緊鄰5’-NGG-3’(PAM)的候選靶序列,靶序列一般大小為18~20 bp。sgRNA引物序列F基本結構:靶序列前加保護堿基(tg)和T7啟動子(TAATACGACTCACTATA)或Sp6啟動子(ATTTAGGTGACACTATA),靶序列后加sgRNA骨架序列上游序列(GTTTTAGAGGCTAGAA ATAGG),sgRNA引物序列R:AAGCACCGACTCGGTGCCACT,根據miR-196a-1基因靶位點通過Primer3.0設計基因組正反檢測引物G196a-1-F和G196a-1-R(表1)。

表1 本文涉及到的引物和模板序列Tab.1 Primers and template sequences involved in this study
1.2.2 sgRNA的合成
使用通用模板(見表1),以sgRNA1-F/sgRNA-R或sgRNA2-F/sgRNA-R為引物,在退火溫度為60 ℃,延伸時間為10 s的條件下,用高保真酶進行PCR擴增,獲得攜帶Sp6啟動子的miR-196a-1基因靶位點序列1以及T7啟動子的miR-196a-1基因靶位點序列2的sgRNA模板序列;PCR產物進行瓊脂糖凝膠電泳并切膠回收,以回收的PCR產物作為模板,用Sp6(或T7)體外轉錄試劑盒合成sgRNA,轉錄產物用RNA純化試劑盒進行純化回收,瓊脂糖凝膠電泳及測定濃度后-80 ℃保存。
1.2.3 顯微注射以及靶位點有效性檢測
收集15 min內產的斑馬魚受精卵,待發育至1細胞期時排列在注射板上,將miR-196a-1基因靶位點的sgRNA1(30 ng/μL)、sgRNA2(35 ng/μL)和Cas9蛋白(150~300 ng/μL)按1∶1∶0.7比例混勻;用定量顯微注射系統將混合溶液(約1 nL)共注射到斑馬魚受精卵中,注射后放置到28.5 ℃的恒溫箱中;受精卵培養至36 h時,分別收集野生型和部分注射后胚胎進行有效性鑒定,剩余胚胎繼續培養至成魚。
1.2.4 可穩定遺傳突變體的篩選
將注射后的胚胎培養至2個月左右發育為幼魚,對幼魚逐條剪尾進行基因型鑒定。由于兩個靶位點之間相距132 bp,如果兩個靶位點都有效,就會造成較大片段的缺失。用基因組檢測引物對基因組進行PCR擴增,隨后進行瓊脂糖凝膠電泳,若PCR產物會同時出現野生型目的條帶以及比野生型目的條帶小100 bp左右的條帶,則這些幼魚為攜帶突變的F0代魚;將F0代幼魚繼續飼養1個月左右至成魚,與野生型進行雜交,用同樣的基因型鑒定方法篩選出可穩定遺傳的魚,這些魚為F1代突變體。基因型鑒定后,根據瓊脂糖凝膠電泳結果,將F1代突變體中比野生型目的條帶小的條帶進行切膠回收,并送公司測序,根據測序結果分析突變位點和突變的堿基數。
2 結果與分析
2.1 miR-196a-1基因生物保守性分析
首先在NCBI網站上(https://www.ncbi.nlm.nih.gov)查找miR-196a-1基因在各個物種中的序列,并將序列下載,利用blast比對各個物種的miR-196a-1基因序列,并找出保守序列。在miRBase網站上(https://www.mirbase.org)查找種子序列,與各個物種的miR-196a-1基因序列比對,發現各個物種的miR-196a-1基因的種子序列高度保守(見圖1)。
2.2 miR-196a-1基因靶位點確定
按1.2.1所述的方法在miR-196a-1基因序列上選擇基因敲除位點(見圖2),靶位點序列前加保護堿基和啟動子序列,靶位點序列后加sgRNA骨架上游序列,將此作為正向引物sgRNA1-F和sgRNA2-F。sgRNA骨架下游序列作為反向引物sgRNA-R。

圖1 各個物種間miR-196a-1基因生物保守性分析Fig.1 Bioconservative analysis of miR-196a-1 gene among species加粗部分為保守序列區,序列標紅處為種子序列。miR-196a-1種子序列區高度保守The bold part showed the conserved sequence region, and the red part of the sequence is the seed sequence. The miR-196a-1 seed sequence is highly conserved

圖2 miR-196a-1基因靶位點示意圖Fig.2 The design diagram of miR-196a-1 gene targeting site以上序列是miR-196a-1基因序列的一部分,大寫加粗部分為外顯子,小寫未加粗部分為內含子;下劃線為直線的表示靶位點序列,下劃線為雙直線的表示為PAM序列;下劃線為波浪線的則是檢測引物Above is partial sequence of the miR-196a-1 gene. The capitalized with bold sequences are exons, the lower case with bold parts are introns;The sequence with single underline indicated the target site1 and target site2 sequence, respectively;The doubled underlines means PAM sequence. Underlined by wavy lines are primers for genotyping
2.3 miR-196a-1基因注射有效性檢測與分析
為了確認靶位點有效,本文按方法1.2.3中所述將注射后胚胎培養至36 hpf (hours post fertilization),隨機挑選5管,每管2顆胚胎,提取基因組DNA進行PCR擴增(見圖3)。結果顯示,3號和4號泳道除了一條210 bp的野生型條帶外,下方有一條較弱的小帶,證明選擇的兩個靶位點都有效,達到基因敲除效果。
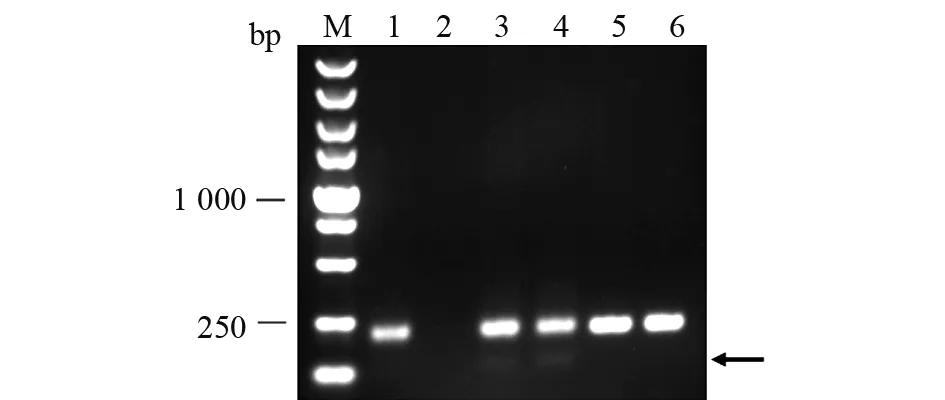
圖3 胚胎注射有效性分析Fig.3 Analysis of effectiveness of embryo injection胚胎注射有效性分析PCR產物凝膠電泳圖。M:DNA標準;1~5:注射胚胎基因組DNA進行PCR擴增的產物;6:野生型對照。3和4泳道除了有一條明亮的210 bp的野生型條帶外,下方有一條較弱的小帶(黑色箭頭指示), 這說明兩個靶位點都有效Gel electrophoresis of PCR products for embryo injection effectiveness analysis. M:DNA marker;1~5:PCR amplification products of genomic DNA with embryo injection;6:Wild type control group. In addition to a bright 210 bp wild-type band in lanes 3 and 4, there is a weaker band (indicated by a black arrow) below, indicating that both target sites are valid
2.4 F0代突變體篩選
將注射有效的剩余胚胎養至成魚,對成魚逐條剪尾提取基因組DNA進行PCR檢測分析(見圖4)。結果顯示第8號泳道除了一條210 bp的野生型條帶外,下方有一條較弱的小帶。

圖4 F0代兩個月幼魚篩選Fig.4 F0 generation two months juvenile screeningM:DNA標準;1~9:兩個月幼魚剪尾提取基因組DNA進行PCR擴增的產物;10:野生型對照。第8號泳道除了有一條明亮的210 bp的野生型條帶外,下方有一條較弱的小帶(黑色箭頭指示)M:DNA marker;1~9:PCR amplified products using two months juvenile genomic DNA injected with miR-196 guide RNAs;10:Wild type control. In addition to a bright 210 bp wildtype band, lane 8 has a weaker band below (Indicated by the black arrow)
2.5 穩定遺傳突變體的篩選
本試驗將篩選到的第8號F0代突變體與野生型斑馬魚雜交,得到F1代魚,收集5管F1代胚胎,每管2顆,提取基因組DNA做PCR擴增并凝膠鑒定,結果如圖5a所示,發現除了210 bp的野生型條帶外,下方有一條100 bp左右的條帶,說明第8號F0代突變體可穩定遺傳。將該100 bp左右的條帶進行切膠回收并送公司測序,峰圖顯示兩個靶位點都出現缺失,結果如圖5b所示。測序序列在NCBI blast 數據庫中進行比對,發現突變體的兩個靶位點之間缺失了103 bp堿基,證明兩個靶位點都有效,并造成大片段的缺失,結果如圖5c所示。將剩余的F1代胚胎養成2個月的幼魚,再逐條剪尾提取基因組DNA進行PCR檢測鑒定,結果如圖5d所示,獲得可穩定遺傳的F1代魚。
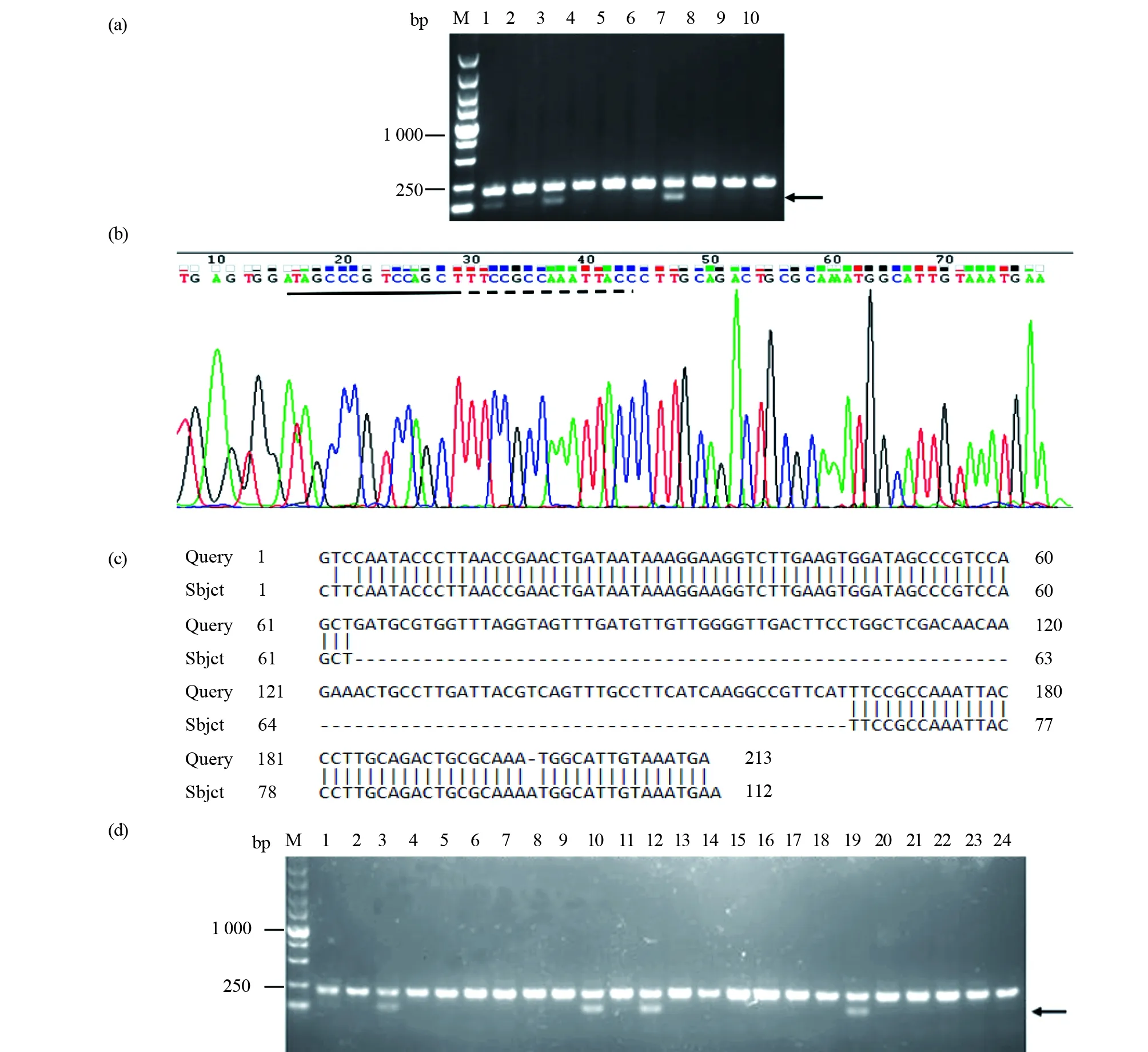
圖5 F1代突變體篩選Fig.5 F1 generation mutant screening(a)F1代胚胎PCR產物凝膠電泳圖。M:DNA標準;1~9:注射胚胎sgRNA后,提取基因組DNA,進行PCR擴增的產物;10:野生型對照。第1、3、7號泳道除了有一條明亮的210 bp的野生型條帶外,下方有一條較弱的小帶(黑色箭頭指示)。證明F0代突變體篩選的8號魚可產生能穩定遺傳的突變后代;(b)F1代胚胎PCR產物測序峰圖。將基因敲除條帶的PCR產物送公司測序,峰值圖中:“----”代表第一號靶位點序列,基因敲除后,第一號靶位點缺失了7個堿基。“”代表第二號靶位點序列,第二號靶位點缺失了8個堿基;(c)測序序列比對結果圖。發現第一號靶位點和第二號靶位點之間缺失103 bp,包括第一號靶位點缺失了7個堿基,第二號靶位點缺失了8個堿基以及第一號與第二號靶位點中間的堿基;(d)F1代突變體成魚PCR產物凝膠電泳圖。M:DNA標準;1~23泳道:注射胚胎基因組DNA進行PCR擴增的產物;24泳道:野生型對照。第1、3、10、12、19號泳道除了有一條明亮的210 bp的野生型條帶外,下方有一條較弱的小帶(黑色箭頭指示)(a)Gel electrophoresis of PCR products using F1 embryos as genomic DNA. M:DNA marker;1~9:PCR products using genomic DNA injected with sgRNA;10:Wild type control. In addition to a bright 210 bp wild-type band,there was a weaker band below in lanes 1, 3, and 7 (indicated by the black arrow). The fish No.8 selected by the F0 mutant can produce mutant offspring that can be stably inherited;(b) The F1 generation mutant embryo PCR product sequencing peak map, the small band of PCR product was purified and sent to the company for sequencing, “----”indicated the first target site sequence,7 bases missing from target site 1;“”marked the second target site sequence,7 bases missing at target 2;(c)Mutant sequence was compared with the control in the NCBI blast datebase,103 bp was missing between the first and second target sites;(d)Gel electrophoresis of PCR products of F1 adult mutant fish. M:DNA marker;1~23:PCR amplified product of genomic DNA with embryo injection;24:Wildtype control. Inaddition to a bright 210 bp wildtype band, there was a weaker band below in lanes 1, 3, 10, 12, 19 (indicated by the black arrow)
3 討論
CRISPR/Cas9基因編輯技術面世之前,一般采用鋅指核酸酶技術(Zinc-finger nucleases,ZFN)和基因打靶技術(transcription activator-like effector nucleases,TALEN)進行基因編輯,但ZFN和TALEN基因打靶技術存在很大的局限性。ZFN基因打靶技術操作過程十分復雜,并且一次不能對多個靶位點進行切割,打靶效率不高。在構建動物模型的過程中,需要多次構建表達載體,構建載體過程繁瑣,試驗周期太長[24-26]。CRISPR/Cas9基因編輯技術相比前兩種技術設計簡單、高效,大大縮短了試驗周期,一經面世,就被廣泛應用。本文用斑馬魚作為模式動物,使用CRISPR/Cas9基因編輯技術敲除miR-196a-1基因,斑馬魚作為模式動物的優勢結合CRISPR/Cas9基因編輯技術的簡單操作,采用Cloning free技術,不需要進行載體的構建,只需以寡聚核苷酸作為模板進行PCR,將PCR產物純化回收后直接作為合成sgRNA的模板,簡化了基因敲除的過程,大大提高了試驗效率,降低了試驗過程中潛在的風險。本文選用了兩個靶位點,采取了注射一對sgRNA的方式,如果兩個靶位點都有效,可以造成大片段的缺失,對斑馬魚易回復突變的特點進行補償作用,敲除結果也可在瓊脂糖凝膠電泳中直觀地觀察到,無需單個檢測。但CRISPR/Cas9基因編輯技術有易脫靶的局限性,這就需要對目的基因的敲除蛋白結構域和靶位點的選擇有更高的要求。
因為顯微注射后的F0代胚胎為嵌合體,其突變不一定能夠遺傳到下一代,因此不直接對F0代突變體測序。篩選到的F0代突變體只進行PCR擴增鑒定,將鑒定有突變的個體與野生型斑馬魚雜交產生F1代,提取F1代胚胎基因組DNA做PCR擴增,瓊脂糖凝膠電泳驗證,隨后送公司進行Sanger測序,確定敲除位點和數量。
F1代突變體能夠穩定遺傳,將F1代雜合突變體自交,經基因型鑒定,出現25%的純合子,與預期結果相符合。將F1代miR-196a-1基因雜合突變體成魚自交,觀察F2代miR-196a-1基因純合突變胚胎,其外表發育正常,未出現明顯表型(圖6),這說明缺失后不影響整體胚胎發育,導致miR-196a-1突變后斑馬魚胚胎整體未出現明顯缺陷,但是該基因是否引起細微結構異常尚不清楚,需要進一步研究。該基因可能還有其他的miRNAs起著代償作用,共同降解靶基因。在后續的研究中,應在miR-196a-1基因敲除模型上進一步敲除其它相關的miRNAs,觀察這些基因在調控斑馬魚腸道發育中的功能。本研究第一次成功構建了miR-196a-1基因敲除斑馬魚模型,為后續的miR-196a-1基因功能研究提供了良好的研究基礎。
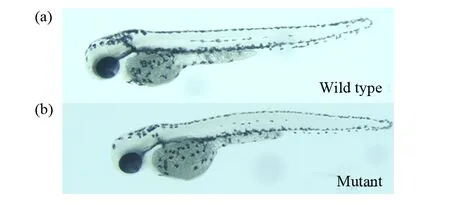
圖6 miR-196a-1基因純合突變體表型 (50 ×)Fig.6 Phenotype of miR-196a-1 null mutant (50 ×)(a)野生型對照組;(b)miR-196a-1基因純合突變體。結果顯示miR-196a-1基因純合突變體與野生型相比,外表沒有明顯變化(a)Wild-type control group;(b)Homozygous mutant of the miR-196a-1 gene. The results showed that the miR-196a-1 homozygous mutant had no obvious difference compared to the wild type
