
當量生物素控制對谷氨酸棒桿菌發酵產L-異亮氨酸的影響
2020-05-29 10:10:22熊海波梅漫莉徐慶陽
中國調味品 2020年5期
熊海波,梅漫莉,徐慶陽,2,3*
(1.天津科技大學 生物工程學院,天津 300457;2.天津市氨基酸高效綠色制造工程實驗室, 天津 300457;3.代謝控制發酵技術國家地方聯合工程實驗室,天津 300457)
L-異亮氨酸是人體必需的氨基酸,同時又是3種支鏈氨基酸之一[1],因其特殊的結構和功能,在人類生命代謝中具有極其重要的地位[2,3]。異亮氨酸是人體激素、蛋白質合成和能量生成的原料,在細胞膜的跨膜運輸中發揮著至關重要的作用[4]。同時作為食品添加劑,可調節食品中的氨基酸平衡,因此,被廣泛應用于食品與醫藥等行業,具有巨大的商業價值[5]。日本在L-異亮氨酸產量、品質和技術水平上均居世界領先地位,2018年,全球生產L-異亮氨酸的主要廠家日本的味之素、協和發酵、田邊制藥以及德國的Degussa均以發酵方式為主,其中,日本味之素占據了全球醫藥及食品用氨基酸市場份額的60%[6-8]。中國L-異亮氨酸發酵產業起步較晚,技術水平落后,糖酸轉化率低,發酵副產物較多,后期分離提取困難,因此在國際上行業競爭能力相對較弱[9],但隨著20世紀90年代中國氨基酸市場的開放,國內發酵行業迅猛發展,目前中國已經成為氨基酸發酵產業大國[10,11]。微生物要大量積累L-異亮氨酸,必須使代謝調節異常化,這種代謝調節異常化的細胞對環境條件是敏感的,所以需要優化發酵條件[12]。當前,微生物發酵法產異亮氨酸主要以大腸桿菌作為工程菌株,但大腸桿菌發酵控制條件復雜,易受噬菌體侵染,且副產物纈氨酸產量較高,分離提取困難。為解決上述問題,提高L-異亮氨酸產量及糖酸轉化率,保證菌種產酸的穩定性,本研究以谷氨酸棒桿菌YILM 1504為供試菌株,以生物素為唯一變量,確定了發酵產L-異亮氨酸過程中最適生物素添加量及中后期補料工藝。
1 材料與方法
1.1 材料與試劑
1.1.1 菌種
谷氨酸棒桿菌(Corynebacteriumglutamate)YILM 1504:由天津科技大學代謝工程實驗室保存。
1.1.2 培養基配方
種子培養基配方:葡萄糖25 g/L,MgSO40.5 g/L,KH2PO45 g/L,(NH4)2SO45 g/L,酵母浸粉5 g/L[13],玉米漿干粉20 g/L,VH0.1 mg/L,VB10.3 mg/L,消泡劑 0.2 g/L,用NaOH調節pH到4.5~5之間,種子發酵罐培養用氨水調節并維持pH到6.7~7.0之間。
發酵培養基配方:葡萄糖80 g/L,MgSO40.8 g/L,KH2PO41.6 g/L,(NH4)2SO45 g/L, 玉米漿干粉(10,20,30,40 g/L,即15,30,45,60 μg/L),VB10.3 g/L,豆餅水解液10 mL/L,賴氨酸0.2 g/L,蛋氨酸0.2 g/L,培養基pH用NaOH調節到4.5~5之間,發酵培養過程中pH用氨水調節并維持到6.7~7.0之間。
1.2 儀器與設備
LDZH-100KBS型全自動立式蒸汽滅菌器 天津博鑫生物科技有限公司;5 L自動控制發酵罐 上海保興生物設備工程有限公司;SBA-40E 生物傳感分析儀 山東省科學院生物研究所;Agilent 1200高效液相色譜儀 Agilent Technologies公司;KQ-C 高壓蒸汽發生器 上海奉賢協新機電廠;752分光光度計 上海分析儀器廠;Olympus生物顯微鏡 日本Olympus 會社。
1.3 方法
1.3.1 菌種培養
從-80 ℃冰箱中取出谷氨酸棒桿菌Corynebacteriumglutamate)YILM 1504保菌管,在無菌間傳兩根斜面試管,32 ℃培養箱培養活化24 h,活化后的菌株再傳2個200 mL茄形瓶,繼續32 ℃培養24 h,保持菌株旺盛的活力[14]。
2 L種子發酵培養,發酵罐接種3 L,32 ℃,維持pH在 6.7~7.0。初始通風2.0 L/min,初始罐壓小于0.05 MPa,初始轉速200 r/min,種子培養時間16 h,期間轉速逐步調節到400 r/min,其他條件不變,OD600大于20×0.8可接發酵培養。
3 L發酵培養,種子罐留600 mL種子液,添加發酵培養基,定容到3 L,發酵液連續培養46 h。溫度32 ℃,pH 6.7~7.0,溶氧保持在30~40之間,轉速由200 r/min逐步提高到900 r/min。通風量由2.0 L/min逐步提升到6.0 L/min。16 h后測殘糖,流加80%(體積分數)的葡萄糖,維持罐內殘糖在10~20 g/L之間。中間添加適量消泡劑消除泡沫[15]。
1.3.2 梯度濃度生物素添加對比實驗
5 L發酵罐中,發酵培養基其他條件不變,10,20,30,40 g/L玉米漿換算成生物素添加量分別為15,30,45,60 μg/L。設置4個生物素添加量梯度,每個梯度3批實驗,探究生物素濃度對菌體量、產酸和副產物的影響。
1.3.3 直接添加與分批流加生物素對比實驗
5 L發酵罐中,分批流加實驗的發酵培養基成分及培養條件與底料直接添加生物素相同,但分批流加生物素實驗,在發酵培養期間,分別在18,32,42 h添加10 g/L的玉米漿(15 μg/L的生物素)。每個實驗設置3個批次,探究流加生物素對菌體量及產酸的影響。
L-異亮氨酸發酵初期為0~18 h,中期為18~32 h,后期為42~56 h,發酵間期指整個發酵過程,為0~56 h。圖6中初期添加指直接添加實驗,間期補加指分批流加實驗。
1.3.4 玉米漿干粉處理方法
用鐵鍋稱取800 g玉米漿干粉,添加2 g消泡劑,溶于2.1 L水中,電磁爐240 ℃煮40 min,得到的玉米漿濃液,冷卻后定容到2 L,制備成40%玉米漿液體備用。
普通玉米漿本身富含糖、氮和微生物生長無機微量元素和有機因子,保存不當容易滋生雜菌,導致酸敗變質,使其成分有較大的波動。而玉米漿干粉為超低溫噴霧干燥的固體顆粒,內在質量比普通液體玉米漿質量穩定(有效活性成分均一而穩定)可控,從而使生物發酵的穩定性和重現性保持良好。該實驗使用玉米漿干粉為保齡寶生物股份有限公司生產,每1 g 40%玉米漿溶液中含有1.5 μg生物素。
1.3.5 異亮氨酸及副產物檢測
發酵液中只檢測到丙氨酸一種主要副產物,因此只需測L-異亮氨酸和丙氨酸含量。
發酵期間,用紙層析法初步檢測產酸量,發酵結束后用高效液相色譜精密檢測產酸量及副產物含量。
紙層析法:點樣1 μL樣品與標品,展開劑為正丁醇∶冰醋酸∶水為4∶1∶2,展開時間20 min,結束后晾干,茚三酮顯色,然后置于80 ℃烘箱中干燥30 s即呈現紅色斑點,對比顏色深淺、斑點大小,得出濃度。
高效液相色譜:發酵液中L-異亮氨酸和丙氨酸濃度用高效液相色譜法測定。采用Agilent C18( 15 mm×4.6 mm,3.5 μm) 色譜柱,衍生劑為2,4-二硝基氟苯,柱前衍生,流動相為50%的乙腈、4.1 g/L 的醋酸鈉溶液,柱溫33 ℃,流速1 mL/min,檢測波長360 nm[16]。
1.3.6 pH的測定
采用pH 6.4~8.0精密pH試紙測定。
1.3.7 殘糖含量檢測
每2 h取樣離心,留上清。用SBA生物傳感分析儀檢測殘糖含量。
1.3.8 菌體量檢測
每2 h取樣,分別稀釋10,20,100倍,用紫外可見分光光度計測量OD600。
菌體生物量=吸光值OD600×稀釋倍數。
注:吸光值范圍在0.2~0.8,超過量程后需換下一個稀釋倍數。
1.3.9 pH在線檢測
發酵罐自帶的梅特勒pH在線檢測,用pH試劑進行放液驗證檢測。培養基pH控制,可用梅特勒電導率儀精密檢測。
1.3.10 計算方法
糖酸轉化率(%)=發酵液中L-異亮氨酸質量濃度(g/L)×發酵液總體積(L)/總耗糖量(g)×100%。
每個實驗數據分別取3個數的平均值,進行單因素方差分析。
2 結果與分析
2.1 生物素濃度對菌體濃度及產酸的影響

圖1 生物素濃度對菌體濃度的影響Fig.1 Effect of biotin concentration on thallus concentration
對于L-異亮氨酸而言,較高生物素有利于菌體生長和L-異亮氨酸積累。生物素是磷酸烯醇式丙酮酸羧化酶的輔基,通過促進CO2的固定能把碳架流導向合成L-異亮氨酸的途徑,但過高的生物素對產酸也會產生不利的效果。玉米漿中含有豐富的生物素及速效氮源,通過添加玉米漿控制生物素含量。在5 L發酵罐中,選擇在發酵初期分別一次性添加10,20,30,40 g/L玉米漿(15,30,45,60 μg/L的生物素),通過多批次生物素濃度對比實驗,在忽略初始接種種子濃度的情況下,生物素濃度越高,菌體生長越迅速。由圖1可知,當生物素濃度處于低適量的情況下,生物素濃度的提高對菌體濃度的增長呈階梯式增長,在生物素濃度為15,30,45 μg/L的5 L發酵罐中,菌體最高濃度OD600分別是101.5,108,124。而當生物素濃度提高到60 μg/L時,菌體量爆發式增長,最高OD600可達到368,高生物素的發酵環境下,菌體生長速度激增,可見生物素濃度對菌體的生長影響很大。
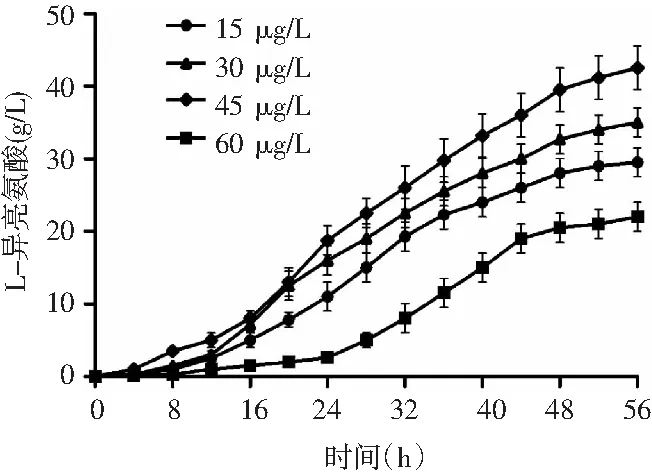
圖2 生物素濃度對L-異亮氨酸產量的影響Fig.2 Effect of biotin concentration on L-isoleucine yield
低適量生物素濃度下,生物素濃度的增加與菌體濃度、異亮氨酸產量呈正相關。在15~45 μg/L的生物素濃度范圍內,產酸量分別是29.5,35,42.5 g/L。在高濃度的生物素發酵液中,微生物的產酸量較低。由圖2可知,當生物素濃度達到60 μg/L時,發酵產酸量為22 g/L。在第24 h才開始快速產酸,高速產酸期延后了8 h,同時菌體量的快速增長消耗了培養基中大部分的營養物質,再加上后期不利物質的積累,第42 h時,菌體提前進入衰退期,產酸進入停滯階段。高濃度的生物素為細胞膜的生長提供了動力,在其他微生物所必需的營養物質都豐富的條件下,生物素濃度越高,微生物的生長情況越好,菌體量越高,但微生物細胞膜的完整性越好,細胞膜的通透性越低,異亮氨酸的分泌越困難,胞內終產物阻遏越強,對代謝途徑中關鍵酶的反饋越強,抑制異亮氨酸的合成,最終導致胞外發酵液中異亮氨酸濃度較低。異亮氨酸作為必需氨基酸的一種,同樣是微生物生長的必要成分,谷棒菌株在合成異亮氨酸的同時,同樣從發酵液中吸收并利用異亮氨酸合成自身所需的物質。
2.2 生物素濃度對耗糖速率及糖酸轉化率的影響

圖3 生物素濃度對耗糖速率的影響Fig.3 Effect of biotin concentration on sugar consumption rate
谷氨酸棒桿菌發酵產L-異亮氨酸,葡萄糖為主要碳源,其消耗主要有兩部分組成:一部分提供細胞生長與自身繁殖的能量與碳骨架,另一部分由細胞合成與分解成各種氨基酸、核苷酸、脂肪酸等中間代謝體。由圖3可知,在10,20,30,40 g/L玉米漿(15,30,45,60 μg/L的生物素)的5 L發酵罐中,初始培養基中糖濃度都是80 g/L,隨著發酵開始,微生物耗糖速率隨生物素添加量的增加而加快,在18~24 h之間達到谷氨酸棒桿菌產酸的最佳糖濃度。谷氨酸棒桿菌的生長與產酸是非同步關系,在8 h以前微生物主要以自身生長與繁殖為主,隨著微生物濃度的增加,耗糖速率緩慢增加;在發酵8 h左右,微生物進入分裂對數期,同時開始產酸,耗糖快速上升;當達到20 h后,細胞濃度達到最大值并保持穩定不變,產酸進入穩定期,耗糖速率基本保持穩定;40 h后,隨著發酵液中營養物質的消耗和副產物的積累,發酵進入后期,微生物濃度和產酸量下降,耗糖速率降低。
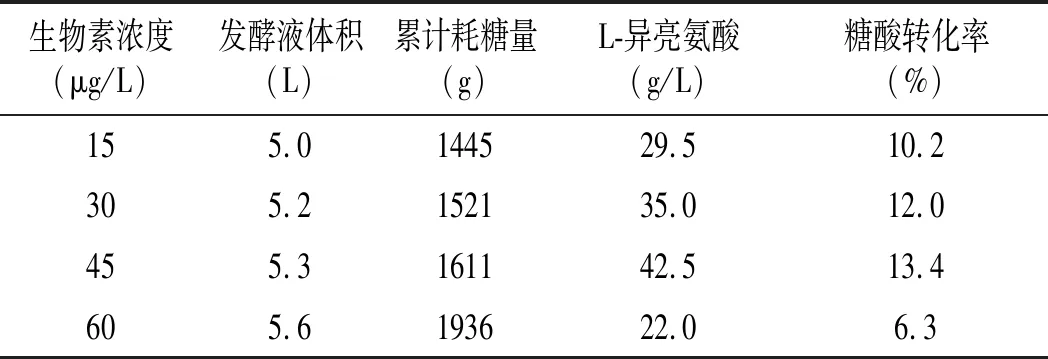
表1 不同生物素發酵參數對比Table 1 Comparison of fermentation parameters of different biotin
由表1可知,生物素的添加量影響微生物的菌體濃度和產酸量,在低適量下,生物素濃度越高,細胞生長越快,菌體量越高,產酸越快,耗糖速率越快,糖酸轉化率越高;在高濃度生物素添加量下,隨著生物素的增加,營養物質大部分用于微生物的生長,產酸期延后,產酸低,糖耗高,糖酸轉化率底。
2.3 生物素濃度對副產物的影響
異亮氨酸合成流程圖見圖4。

圖4 異亮氨酸合成流程圖Fig.4 Flow chart of isoleucine synthesis
注:PC為磷酸烯醇式丙酮酸酶;AK為天冬氨酸激酶;HD為高絲氨酸脫氫酶;TD為蘇氨酸脫水酶;AS為酰羥基酸合成酶;HT為高絲氨酸激酶。

圖5 不同生物素濃度發酵對丙氨酸的影響Fig.5 Effects of fermentation with different biotin concentration on alanine
本實驗使用的谷氨酸棒桿菌為賴氨酸、蛋氨酸缺陷菌株,從液相檢測可以看出,副產物主要為丙氨酸。從代謝流分析得出,葡萄糖一部分經過EMP途徑分解合成丙酮酸再合成丙氨酸,另一部分經過EMP途徑合成磷酸烯醇式丙酮酸,再經過TCA循環合成天冬氨酸,再合成丙氨酸。在整個異亮氨酸合成途徑中,合成天冬氨酸之前的所有途徑相同,所以低適量生物素發酵過程中,隨著生物素濃度的增加,異亮氨酸的合成增加,丙氨酸含量也隨之增加。L-異亮氨酸的生產和丙氨酸的生成為協同增效。在高濃度生物素添加量發酵過程中,細胞膜完整,對氨基酸運出細胞膜也有一定的影響。由圖5可知丙氨酸最高為9 g/L。
2.4 分批流加玉米漿對產酸的影響

圖6 發酵間期補加玉米漿對菌體濃度的影響Fig.6 Effect of corn syrup supplementation on thallus concentration during fermentation

圖7 發酵間期補加玉米漿對L-異亮氨酸的影響Fig.7 effect of corn syrup supplementation on L-isoleucine during fermentation
從上述分析可以得出,高濃度生物素添加量發酵不利于菌體產酸,對于這種情況,本實驗通過分批流加生物素技術,在發酵期間補加生物素,解決了這個難題。分批式添加生物素,在總體玉米漿濃度50 g/L(生物素濃度75 μg/L)不變的情況下,在初始發酵培養基中添加20 g/L玉米漿(30 μg/L生物素),在發酵18,32,42 h分別補加10 g/L玉米漿(15 μg/L生物素)。由圖6和圖7可知,在發酵18 h以前,玉米漿中的速效氮源使菌體生長迅速,同時生物素濃度為30 μg/L,處于低適量生物素添加量,菌體轉型時間點提前,轉型時間由14~18 h提前到10~14 h,菌體轉型快,產酸量高;到18 h以后,大部分菌體已經轉型成功,細胞膜完整性差,轉變成有利于異亮氨酸的積累和胞外分泌的新模式,底料生物素基本被消耗,此時添加少量玉米漿,補充生物素及速效氮源,細胞菌體量進一步增加,同時低適量生物素無法使細胞膜修復完整,細胞膜仍處于缺陷狀態,而補加的速效氮源,有利于菌體生長和異亮氨酸分泌;32 h和42 h生物素再次被消耗完,此時添加玉米漿可保持菌體量濃度不變或下降速度變緩,延長產酸時間,提高轉化率。產量達到了44.5 g/L,比初始生物素濃度為30 μg/L,后期不補料產酸量提高了49.8%。
異亮氨酸發酵的高效產酸關鍵在于控制發酵前中期谷氨酸棒桿菌細胞膜在結構和功能上的特異性轉變[17],增強細胞膜的通透性,從而形成有利于異亮氨酸胞內積累與胞外分泌的新模式[18-21]。這就需要掌握異亮氨酸高產菌株的發酵特性以及該菌株特異型轉變的周期與細胞轉型的關鍵時間點。生物素小分子在參與羧化作用時,能夠結合羧基,參與菌體內固定 CO2的羧化反應;作為乙酰CoA的輔酶,參與磷脂的合成和細胞膜的組成,最終影響目的產物的合成與分泌[22]。在發酵初期,罐內生物素對于菌體來說是過量的,在營養成分豐富的情況下,細胞進入快速生長分裂階段。當發酵進入中期(18 h)后,罐內生物素對于菌體處于亞適量狀態,菌體生長緩慢,細胞膜合成不足,封閉性減弱,通透性增加,細胞內外物質交換能力增強,有利于L-異亮氨酸分泌于胞外[23]。細胞內L-異亮氨酸濃度降低,解除終產物反饋抑制,細胞大量合成L-異亮氨酸。玉米漿在氨基酸發酵中為菌體提供氮源和生長因子,尤其是其中的生物素和限制性氨基酸的含量對發酵結果影響巨大[24]。本研究通過在培養基中添加不同濃度玉米漿的方式控制生物素的添加量,同時,玉米漿中豐富的氨基酸為菌體的轉氨作用提供了原料,促進了異亮氨酸的合成。生物素的亞適量工藝目前已普遍應用于谷氨酸、纈氨酸、亮氨酸等各類氨基酸的發酵生產。
3 結論
本研究通過限制生物素濃度探究谷氨酸棒桿菌YILM 1504產L-異亮氨酸的最適生物素濃度。5 L發酵罐中,在發酵液中生物素處于低適量條件下,隨著生物素濃度的增加,菌體濃度增加,耗糖速率加快,產酸量高,糖酸轉化率高;經過多批次對比實驗,得出最適生物素濃度為45 μg/L,此時產酸達到42.5 g/L,糖酸轉化率達到最高,為13.4%;在高濃度生物素發酵液中,生物素的增加只會增加菌體濃度,不利于異亮氨酸發酵。為了解決了高濃度生物素發酵中后期菌體活力低而造成產酸低的問題,在發酵前期控制生物素濃度為30 μg/L,保持低適量生物素,有利于發酵前期細胞轉型;中期在18 h向發酵液中添加10 g/L玉米漿(15 μg/L生物素),保持少量的生物素,有利于增加轉型細胞濃度,提高產酸速率;后期在32,42 h繼續補加玉米漿,維持細胞濃度,延長高速產酸時間,提高糖酸轉化率,降低成本;54 h產酸量達到了44.5 g/L,比初始生物素濃度為30 μg/L,后期不補料產酸量提高了49.8%。
