大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因的鑒定及生物信息學(xué)分析
2020-06-04 03:21:36張迎春孫天歌張小雪梁其干李艷軍
西北農(nóng)業(yè)學(xué)報(bào) 2020年5期
張迎春,孫天歌,張小雪,梁其干,李艷軍,孫 杰
(石河子大學(xué) 農(nóng)學(xué)院,新疆生產(chǎn)建設(shè)兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,新疆石河子 832003)
黃萎病被稱為棉花的“癌癥”,是制約中國(guó)棉花生產(chǎn)的主要因素之一。中國(guó)棉花黃萎病主要是大麗輪枝菌(Verticilliumdahliae)和黑白輪枝菌(Verticilliumalboatrum)侵染所致。大麗輪枝菌是一種土傳性病原真菌,可侵染200多種植物,其寄主主要是雙子葉植物[1-2]。大麗輪枝菌的微菌核能在土壤中存活30~50年,當(dāng)感應(yīng)到寄主植物根際釋放的根系分泌物后,微菌核開始萌發(fā)產(chǎn)生感染性菌絲可定植于根皮層,進(jìn)而擴(kuò)散到木質(zhì)部產(chǎn)生孢子,導(dǎo)致作物的維管束受到阻塞,出現(xiàn)枯萎癥狀,發(fā)育遲緩,黃化和壞死[3-5],嚴(yán)重影響棉花的產(chǎn)量和品質(zhì)。因中國(guó)氣候變化多樣,大麗輪枝菌在不同環(huán)境條件下產(chǎn)生了許多具有豐富遺傳多樣性的生理小種,田間致病性易發(fā)生變異,出現(xiàn)強(qiáng)致病力的類型[6-7],大麗輪枝菌的防治目前仍無有效的生物及化學(xué)措施。大麗輪枝菌全基因組數(shù)據(jù)的公布為其致病分子機(jī)制的研究提供了基礎(chǔ),為抗病新品種的開發(fā)提供了新路線[8]。
植物細(xì)胞壁由纖維素、半纖維素以及果膠等多糖組成,這些組分與木質(zhì)素一起形成致密的網(wǎng)狀結(jié)構(gòu),形成植物的支撐結(jié)構(gòu)[9]。在大麗輪枝菌侵染植物過程中分泌大量的細(xì)胞壁降解酶(cell wall degrading enzymes, CWDEs)[10]。細(xì)胞壁降解酶不僅可降解植物細(xì)胞壁中的多糖,而且也破壞植物細(xì)胞壁結(jié)構(gòu),為病原微生物的入侵和攝取營(yíng)養(yǎng)提供條件[11]。細(xì)胞壁降解酶包括角質(zhì)酶、多聚半乳糖醛酸酶、果膠酯酶、果膠裂合酶、木聚糖酶、內(nèi)葡聚糖酶G1、β-1,4-葡萄糖苷酶和蔗糖非發(fā)酵蛋白激酶等[11]。角質(zhì)酶VdCUT11水解植物細(xì)胞壁的角質(zhì)層促進(jìn)大麗輪枝菌的侵染[12]。果膠裂解酶VdPL1可誘導(dǎo)植物免疫反應(yīng),同時(shí)作為一個(gè)毒力因子參與大麗輪枝菌的侵染[13]。除角質(zhì)酶和果膠裂解酶外,纖維素酶、聚半乳糖醛酸酶、木聚糖酶等在大麗輪枝菌侵染植物的過程中也發(fā)揮了重要作用[13]。
植物細(xì)胞的感染需要半纖維素的降解[14],木聚糖是植物細(xì)胞壁中半纖維素的主要成分。在蘋果樹爛腐病研究中,木聚糖酶被證明與致病性或毒力有關(guān)[15]。在灰霉病菌研究中,木聚糖酶基因xyn11A缺失突變體的致病力顯著降低,但未完全喪失致病力[16]。木聚糖酶屬于水解酶類,主要包括有β-1,4-內(nèi)切木聚糖酶、β-1,4-外切木聚糖酶、β-1,4-木糖苷酶、α-L-呋喃阿拉伯糖苷酶、α-葡萄糖醛酸酶和乙酰木聚糖酯酶等[17]。其中,β-1,4-內(nèi)切木聚糖酶是使木聚糖完全水解最關(guān)鍵的酶,其作用于木聚糖主鏈的木聚糖苷鍵,隨機(jī)切斷主鏈內(nèi)的糖苷鍵,水解木聚糖而生成分子量大小不同的寡糖[18]。β-1,4-內(nèi)切木聚糖酶能使組成細(xì)胞壁的不溶性木聚糖變成可溶性木聚糖,從而迅速降解細(xì)胞壁[19]。在病原菌侵染植物的過程中,β-1,4-內(nèi)切木聚糖酶參與了細(xì)胞壁的降解,表明它在致病過程中發(fā)揮著重要作用[20]。然而,目前大麗輪枝菌中木聚糖酶研究仍較少,本研究從大麗輪枝菌基因組中鑒定β-1,4-內(nèi)切木聚糖酶基因并進(jìn)行生物信息學(xué)分析,同時(shí)對(duì)這些基因在棉花不同抗感品種根系分泌物中的表達(dá)模式進(jìn)行分析,旨在揭示β-1,4-內(nèi)切木聚糖酶基因與大麗輪枝菌致病性的聯(lián)系,為進(jìn)一步研究β-1,4-內(nèi)切木聚糖酶基因在大麗輪枝菌致病過程中的作用以及揭示致病的分子機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1 材料與處理
大麗輪枝菌V991菌株由石河子大學(xué)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室保存,2個(gè)陸地棉品種‘新陸早7號(hào)’(感黃萎病)和‘中植棉2號(hào)’(耐黃萎病)均由石河子大學(xué)棉花研究所提供。
1.1.1 棉花根系分泌物的收集 將‘新陸早7號(hào)’和‘中植棉2號(hào)’品種的種子浸泡在1%(1 g NaClO加入100 mL H2O)NaClO溶液中進(jìn)行表面消毒,用無菌蒸餾水反復(fù)沖洗后種在經(jīng)過高溫高壓(121 ℃,60 min)滅菌處理的沙土中。每個(gè)品種種植2盆,每盆9株苗,光周期為16 h/8 h,溫度28 ℃,每3 d用霍格蘭營(yíng)養(yǎng)液澆灌1次。 45 d后,從沙土中取出棉苗,用2 L滅菌水浸泡砂土,使根系分泌物充分溶解在水中,用細(xì)菌過濾器 (0.22 μm)過濾水溶液,在冷凍干燥機(jī)中濃縮至0.5 L。
1.1.2 大麗輪枝菌的培養(yǎng)與取樣 將貯存的大麗輪枝菌V991分生孢子培養(yǎng)于察氏培養(yǎng)基上,25 ℃黑暗條件下培養(yǎng)5 d。收集新鮮的分生孢子,分別稱取0.5 g分生孢子接菌于5 mL不同抗感棉花品種的根系分泌物中。25 ℃ 220 r/min搖培0、6和12 h后,收集V991的分生孢子。根系分泌物培養(yǎng)前0 h的大麗輪枝菌樣本,標(biāo)記為CK;感病品種‘新陸早7號(hào)’根系分泌物培養(yǎng)6 h和12 h的樣本,分別標(biāo)記為VDX6和VDX12;耐病品種‘中植棉2號(hào)’根系分泌物培養(yǎng)6 h和12 h的樣本,分別標(biāo)記為VDZ6和VDZ12;同時(shí)用水培養(yǎng)大麗輪枝菌,培養(yǎng)6 h和12 h的樣本分別標(biāo)記為VDW6和VDW12。每處理2個(gè)重復(fù)用于轉(zhuǎn)錄組測(cè)序,共構(gòu)建14個(gè)文庫(kù)。
1.2 方 法
1.2.1 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因的鑒定 從http://fungi.ensembl.org/Verticillium_dahliae/Info/Index/ 下載大麗輪枝菌全基因組編碼序列CDS(Coding sequence) GCF_000150675.1_ASM15067v2-rna序列文件、GCF_000150671_ASM15067v2-protein 蛋白質(zhì)序列文件、GCF_000150675.1_ASM15067v2_ genomic全基因組序列文件,以及Verticillium_dahliaeASM15067v2.42 GFF3文件。利用TBtools v 0.66443軟件對(duì)GFF3文件中所有β-1,4-內(nèi)切木聚糖酶基因進(jìn)行搜索,同時(shí)在CDS、蛋白質(zhì)、全基因組序列文件中獲取基因?qū)?yīng)的cDNA序列、蛋白質(zhì)序列以及基因組序列。
1.2.2 β-1,4-內(nèi)切木聚糖酶基因的氨基酸序列分析 利用ExPAsy(http://web.expasy.org)工具在線分析β-1,4-內(nèi)切木聚糖酶基因一級(jí)結(jié)構(gòu)的基本理化性質(zhì),包括氨基酸序列大小、分子質(zhì)量和等電點(diǎn)等。利用在線工具WOLFPSORT(https://wolfpsort.hgc.jp)和 TMHMM(http://www.cBMs.dtu.dk/services/TMHMM/)分別對(duì)編碼蛋白質(zhì)的亞細(xì)胞定位情況和跨膜結(jié)構(gòu)域進(jìn)行預(yù)測(cè)[21]。
1.2.3 多重序列比對(duì)和進(jìn)化樹分析 采用DNAMAN 6.0和ClustalX 2.0對(duì)β-1,4-內(nèi)切木聚糖酶氨基酸序列進(jìn)行多重序列比對(duì)。利用SMART工具在線預(yù)測(cè)11個(gè)β-1,4-內(nèi)切木聚糖酶蛋白序列的信號(hào)肽和結(jié)構(gòu)域,并用DOG 1.0軟件進(jìn)行繪制。以11個(gè)β-1,4-內(nèi)切木聚糖酶蛋白序列的主結(jié)構(gòu)域?yàn)樘结樞蛄校贑aZy數(shù)據(jù)庫(kù)中(http://www.cazy.org/)查找其他物種中的相應(yīng)蛋白[22],去除重復(fù)以及不完整序列后,挑選9條其他物種中的β-1,4-內(nèi)切木聚糖酶蛋白序列用于進(jìn)化樹分析。利用MEGA 6.0提供的ClustalW程序?qū)Υ篼愝喼捌渌锓N中的β-1,4-內(nèi)切木聚糖酶基因進(jìn)行多重序列比對(duì),然后通過鄰位相連法構(gòu)建基因的復(fù)合進(jìn)化樹。
1.2.4 β-1,4-內(nèi)切木聚糖酶基因的染色體分布 從大麗輪枝菌全基因組數(shù)據(jù)庫(kù)中獲得β-1,4-內(nèi)切木聚糖酶基因染色體分布以及長(zhǎng)度信息,利用TBtools v0.66443軟件繪制β-1,4-內(nèi)切木聚糖酶基因的染色體分布圖。
1.2.5 β-1,4-內(nèi)切木聚糖酶基因結(jié)構(gòu)分析 從全基因組數(shù)據(jù)庫(kù)下載該基因的外顯子和內(nèi)含子信息,利用在線軟件Gene Structure Display Server (GSDS)(http://gsds.cBMi.pku .edu.cn/)分析大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因外顯子-內(nèi)含子的數(shù)目及分布。
1.2.6 大麗輪枝菌樣本RNA的提取及轉(zhuǎn)錄組測(cè)序 采用RNA simple total kit試劑盒(Tiagen,中國(guó)北京)提取大麗輪枝菌總RNA,利用Nanodrop○R2000分光光度計(jì)(Thermo Scientific,Wilmington,de,USA)和Agilent 2100生物分析儀(Agilent TeGHnologies,Santa Clara,CA,USA)分別檢測(cè)RNA的純度和完整性,利用Qubit○R2.0熒光計(jì)(Thermo Scientific,Wilmington,de,USA)對(duì)RNA的濃度進(jìn)行精確測(cè)量,最終獲得高質(zhì)量的RNA樣本用于RNA測(cè)序。采用Illumina HiSeqTM4000測(cè)序平臺(tái)進(jìn)行轉(zhuǎn)錄組測(cè)序,由北京諾禾致源科技股份有限公司完成,轉(zhuǎn)錄組數(shù)據(jù)已上傳至NCBI SRA數(shù)據(jù)庫(kù)中,登錄號(hào)為PRJNA545805。
1.2.7 β-1,4-內(nèi)切木聚糖酶基因?qū)Σ煌藁垢衅贩N根系分泌物的響應(yīng)分析 對(duì)轉(zhuǎn)錄組測(cè)序獲得的raw reads進(jìn)行質(zhì)量控制后,利用HISAT軟件將得到的clean reads比對(duì)到大麗輪枝菌的參考基因組數(shù)據(jù)(http//www.broadinstitute.org/annotation/genome/Verti- cillium dahlia /Blast.html)。采用HTSeq軟件對(duì)各樣品進(jìn)行基因表達(dá)水平分析,使用的模型為union,分別統(tǒng)計(jì)單個(gè)基因的表達(dá)水平。FPKM(fragments per kilobase of exon per millon fragments mapped)為每百萬fragments中來自某一基因每千堿基長(zhǎng)度的fragments數(shù)目,同時(shí)考慮測(cè)序深度和基因長(zhǎng)度對(duì)fragments計(jì)數(shù)的影響,是目前最為常用的基因表達(dá)水平估算方法[22]。根據(jù)轉(zhuǎn)錄組測(cè)序結(jié)果獲得β-1,4-內(nèi)切木聚糖酶基因的FPKM值,并采用HemI軟件繪制基因在不同抗感棉花品種根系分泌物中的表達(dá)熱圖,使用Excel 2010制作基因表達(dá)折線圖。
2 結(jié)果與分析
2.1 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因的鑒定
通過TBtools軟件查找GFF3文件中所有 β-1,4-內(nèi)切木聚糖酶基因,共獲得11條關(guān)于β-1,4-內(nèi)切木聚糖酶基因的記錄,分別為VDAG_03385~VDAG_08273(表1)。這些基因編碼序列(coding sequence, CDS)長(zhǎng)度為666~1 314 bp,編碼221~437個(gè)氨基酸,蛋白質(zhì)分子質(zhì)量為23.90~47.64 ku,理論等電點(diǎn)為5.09~10.00。對(duì)各成員序列的亞細(xì)胞定位預(yù)測(cè)表明,這些基因大多(7個(gè))為細(xì)胞膜外蛋白,部分定位于細(xì)胞膜、線粒體、細(xì)胞質(zhì)基質(zhì)和細(xì)胞核中(表1)。其結(jié)果與角毛殼菌生物信息學(xué)分析一致,說明該酶蛋白主要在胞外發(fā)揮水解木聚糖的生物學(xué)作用[23]。
2.2 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶的保守域分析
利用蛋白跨膜結(jié)構(gòu)域在線分析工具TMHMM(http://www.cBMs.dtu.dk/services/)發(fā)現(xiàn),β-1,4-內(nèi)切木聚糖酶蛋白不含跨膜結(jié)構(gòu)域(transmembrane domain, TM)。利用在線工具SMART(http://smart.embl-heidelberg.de/smart/batGH.pl)對(duì)11個(gè)β-1,4-內(nèi)切木聚糖酶蛋白序列進(jìn)行信號(hào)肽和結(jié)構(gòu)域預(yù)測(cè),分析發(fā)現(xiàn)11個(gè)酶中 VDAG_03790、 VDAG_05042、 VDAG_05043、 VDAG_05294和 VDAG_06165均含有糖苷水解酶Glyco_hydro11結(jié)構(gòu)域,其中 VDAG_06165還含有一個(gè)fCBD(cellulse biding domain;真菌纖維素結(jié)合域)結(jié)構(gòu)域; VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744均含有糖苷水解酶Glyco_hydro10結(jié)構(gòu)域; VDAG_05509和 VDAG_03808均含有糖苷水解酶 Glyco_Hydro 43結(jié)構(gòu)域,其中 VDAG_03808還含有一個(gè)CBM(carbohydrate binding modules;碳水化合物結(jié)合域)結(jié)構(gòu)域(圖1)。VDAG_03385、 VDAG_08273、 VDAG_04744、 VDAG_06165V、DAG_03809和 VDAG_05509含有不同長(zhǎng)度的信號(hào)肽(圖1)。
將 VDAG_03790、 VDAG_05042、 VDAG_05043、 VDAG_05294和 VDAG_06165蛋白的Glyco_hydro11結(jié)構(gòu)域進(jìn)行多重序列比對(duì),發(fā)現(xiàn)GGKGW、YGW、EYY和STR區(qū)域具有較強(qiáng)的保守性(圖2-A)。將 VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744蛋白的Glyco_hydro10的結(jié)構(gòu)域進(jìn)行多重序列比對(duì),保守區(qū)域?yàn)閃DVVNE、ADP、NDY和TELD(圖2-B)。對(duì)VDAG_03808和VDAG_05509蛋白的Glyco_Hydro 43結(jié)構(gòu)域進(jìn)行多重序列比對(duì),KGVRD和IATDL區(qū)域具有較強(qiáng)的保守性(圖2-C)。這些保守性區(qū)域可能與木聚糖酶的催化相關(guān)[24-25]。
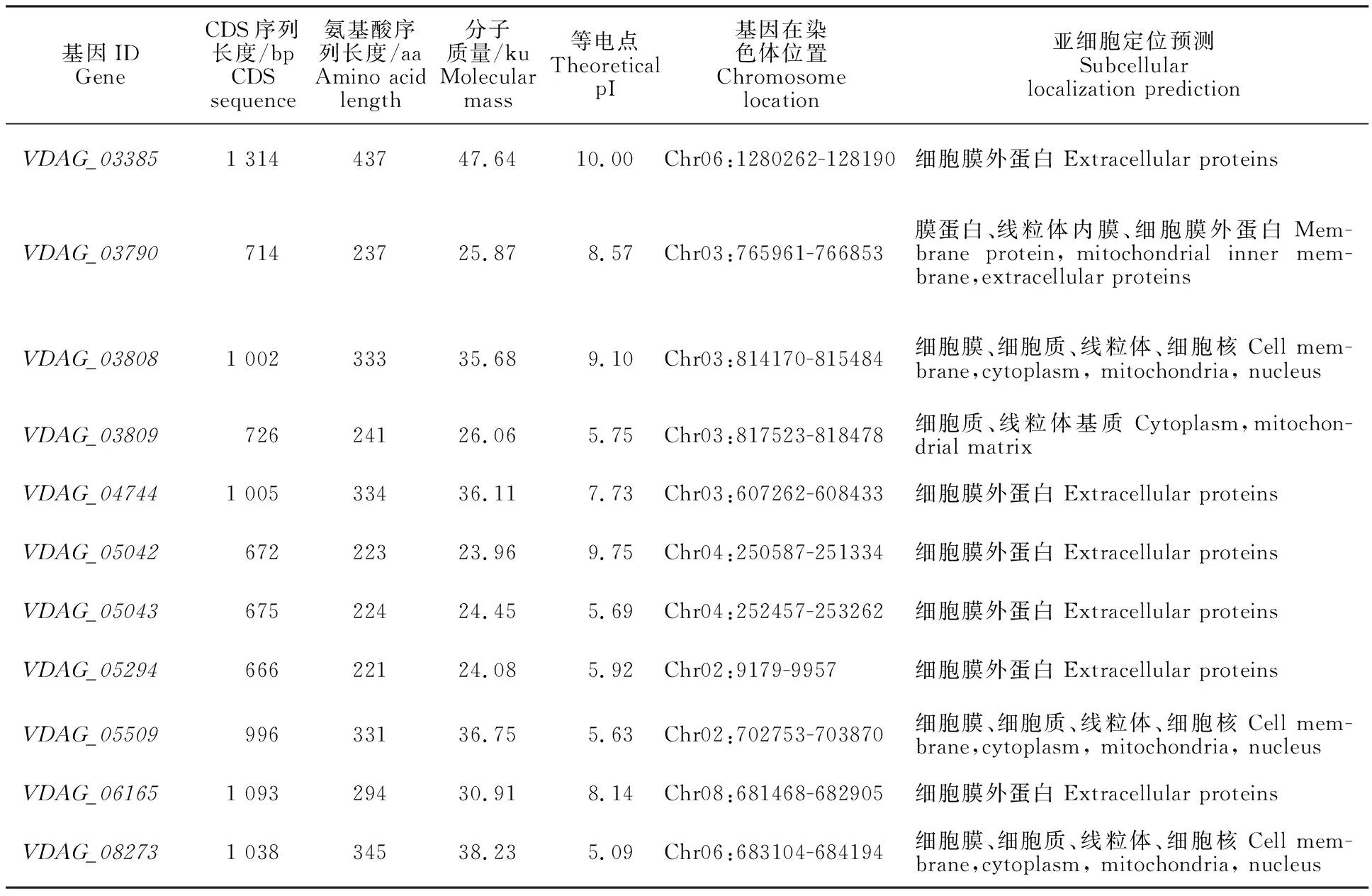
表1 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因的特征Table 1 Characterization of Endo-β-1,4-xylanase genes in V.dahliae
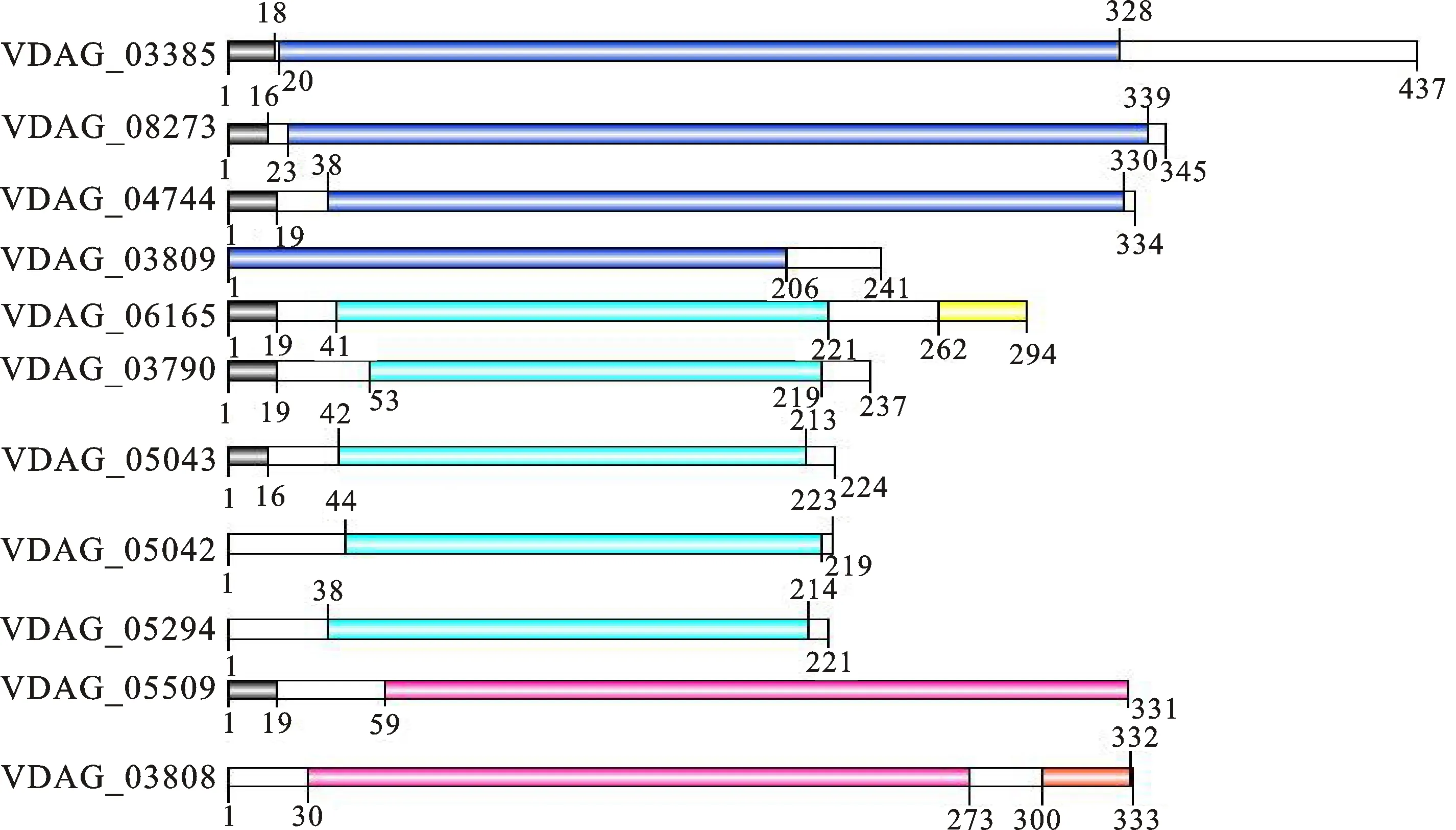
黑色表示信號(hào)肽;深藍(lán)色表示Glyco_hydro10結(jié)構(gòu)域;淺藍(lán)色表示Glyco_hydro11結(jié)構(gòu)域;粉色表示Glyco_Hydro 43結(jié)構(gòu)域;黃色表示fCBD結(jié)構(gòu)域;橙紅色表示CBM結(jié)構(gòu)域
The black indicates signal peptide;Dark blue indicates the Glyco_hydro10 domain; Light blue indicates the Glyco_hydro11 domain; Pink indicates the Glyco_Hydro 43 domain; Yellow indicates the fCBD domain;Orange red indicates the CBM domain
圖1 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶蛋白結(jié)構(gòu)域分析
Fig.1 Domains analysis of Endo-β-1,4-xylanase proteins inV.dahliae

A為VDAG_03790、VDAG_05042、VDAG_05043、VDAG_05294和VDAG_06165蛋白Glyco_hydro11結(jié)構(gòu)域多重序列比對(duì);B為VDAG_08273、VDAG_03385、VDAG_03809和VDAG_04744蛋白Glyco_hydro10結(jié)構(gòu)域多重序列比對(duì);C為VDAG_03808和VDAG_05509蛋白Glyco_Hydro 43結(jié)構(gòu)域多重序列比對(duì)。灰色和黑色區(qū)域分別代表 75%以上和 100%的保守區(qū)。黑色方框表示氨基酸保守區(qū)域
A indicates a multiple sequence alignment of Glyco_hydro11 domain in VDAG_03790, VDAG_05042, VDAG_05043, VDAG_05294, and VDAG_06165 proteins; B indicates a multiple sequence alignment of Glyco_hydro10 domain in VDAG_08273, VDAG_03385, VDAG_03809 and VDAG_04744 proteins; C indicates a multiple sequence alignment of Glyco_Hydro 43 domain in VDAG_03808 and VDAG_05509 proteins.The gray and black areas represent more than 75% and 100% of conservative regions, respectively.Black boxes indicate regions of amino acid conservation
圖2 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶蛋白結(jié)構(gòu)域多重序列比對(duì)
Fig.2 Multiple sequence alignment of domains in Endo-β-1,4-xylanase proteins fromVerticilliumdahliae
2.3 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶蛋白的進(jìn)化關(guān)系分析
在CaZy數(shù)據(jù)庫(kù)中查找并隨機(jī)選取9個(gè)來自其他物種的β-1,4-內(nèi)切木聚糖酶蛋白記錄,其中XP_018153852.1(十字花科炭疽病菌)、EPS36346.1(捕食線蟲真菌)、ABR14629.1(宇佐美曲霉)和RGP60782.1(擬枝孢鐮刀菌)屬于糖苷水解酶(glycoside hydrolase,GH)10家族成員,BAE71410.1(黑色素短梗霉)、ACS96449.1(嗜酸真菌)和ACR83565.1(黑曲霉)屬于11家族成員,XP_013423968.1(出芽短梗霉)和THY53895.1(出芽短梗霉)屬于43家族成員。利用MEGA 6.0軟件將上述9個(gè)蛋白與11個(gè)大麗輪枝菌β-1,4-內(nèi)切木聚糖酶蛋白進(jìn)行進(jìn)化關(guān)系分析,結(jié)果如圖3所示,20個(gè)糖苷水解酶基因被清晰地分為3組。 VDAG_06165、 VDAG_03790、 VDAG_05294、VDAG_0 5042和 VDAG_05043與其他物種中11家族成員聚為一類; VDAG_08273、 VDAG_03385、 VDAG_03809和 VDAG_04744與其他物種中10家族成員聚為一類; VDAG_05509和 VDAG_03808與其他物種中43家族成員聚為一類。該結(jié)果與結(jié)構(gòu)域預(yù)測(cè)結(jié)果相一致,表明大麗輪枝菌11個(gè)β-1,4-內(nèi)切木聚糖酶蛋白中,含有5個(gè)糖苷水解酶11家族成員,4個(gè)10家族成員和2個(gè)43家族成員。
2.4 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因結(jié)構(gòu)分析
利用在線軟件GSDS對(duì)獲得的11個(gè)β-1, 4-內(nèi)切木聚糖基因的結(jié)構(gòu)進(jìn)行分析,結(jié)果如圖4所示:VDAG_06165基因外顯子和內(nèi)含子最多,含有7個(gè)外顯子和6個(gè)內(nèi)含子;VDAG_05042和VDAG_08273基因外顯子和內(nèi)含子較少,分別含有2個(gè)外顯子和1個(gè)內(nèi)含子。其余8個(gè)基因中,4個(gè)基因(VDAG_03385、VDAG_03809、VDAG_04744和VDAG_03808)含有4個(gè)外顯子和3個(gè)內(nèi)含子,4個(gè)基因(VDAG_05294、VDAG_05043、VDAG_05042和VDAG_05509)含有3個(gè)外顯子和2個(gè)內(nèi)含子。

10、11和43分別指糖苷水解酶10、11和43家族
10、11 and 43 refer to glycoside hydrolase families of 10、 11 and 43, respectively
圖3 大麗輪枝菌和其他物種中β-1,4-內(nèi)切木聚糖酶蛋白進(jìn)化樹分析
Fig.3 Phylogenetic tree analysis of Endo-β-1,4-xylanase proteins inV.dahliaeand other species
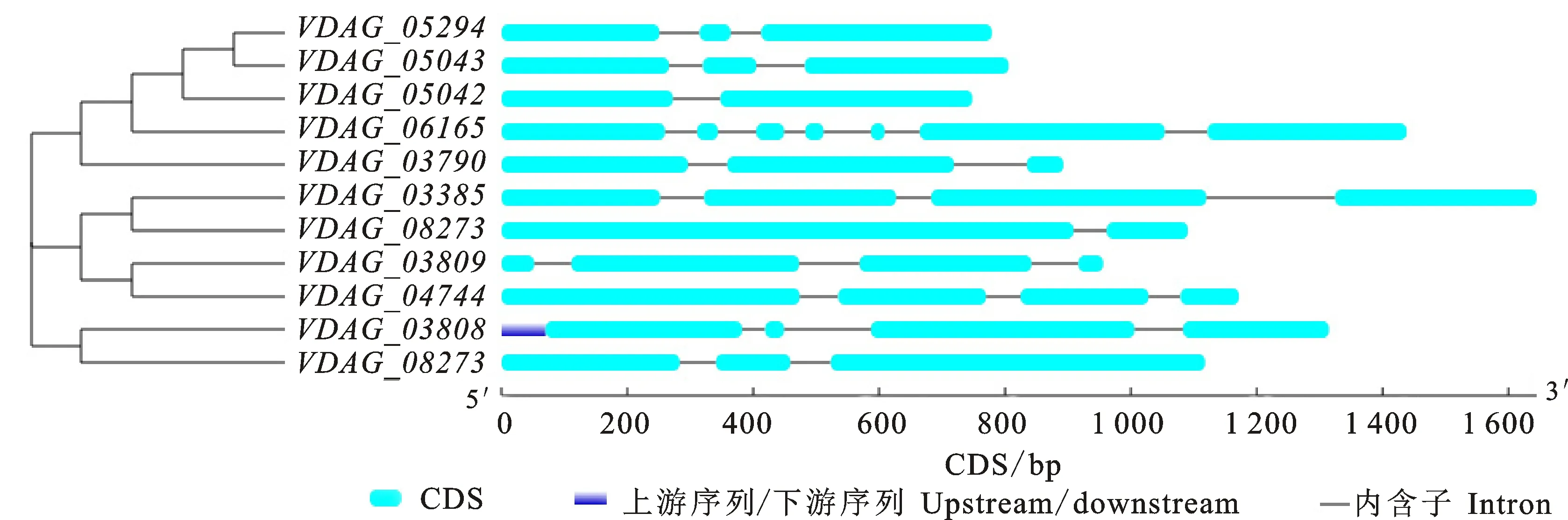
藍(lán)色表示UTR(上下游序列),綠色表示編碼區(qū),灰線表示內(nèi)含子區(qū)
The blueindicates UTR, the green indicates the exon, and the gray indicates the intron
圖4 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因結(jié)構(gòu)分析
Fig.4 Gene structures analysis of Endo-β-1,4-xylanase genes inV.dahliae
2.5 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因的染色體分布
大麗輪枝菌有8條染色體,Chr01長(zhǎng)度為 5 243 713 bp,Chr02為5 264 584 bp,Chr03為 5 534 790 bp,Chr04為3 754 323 bp,Chr05為 3 249 347 bp,Chr06為2 537 490 bp,Chr07為 3 147 721bp,Chr08為2 984 914 bp。11個(gè)β-1,4-內(nèi)切木聚糖酶基因分布在其中5條染色體上(圖5)。3號(hào)染色體上分布的β-1,4-內(nèi)切木聚糖酶基因最多,包括VDAG_03790、VDAG_03808、VDAG_03809和VDAG_04744四個(gè)基因,它們的基因組序列長(zhǎng)度分別為893 bp、1 315 bp、956 bp和1 172 bp。8號(hào)染色體上分布的基因最少,僅含VDAG_06165基因,其基因組長(zhǎng)度為1 438 bp。2號(hào)、4號(hào)和6號(hào)染色體上分別分布2個(gè)基因,VDAG_05294與VDAG_05509分布于2號(hào)染色體上,它們的基因組序列大小分別為779 bp和1 118 bp;VDAG_05042和VDAG_05043分布于4號(hào)染色體上,它們的基因組序列大小分別為748 bp和806 bp;VDAG_03385和VDAG_08273分布于6號(hào)染色體上,它們的基因組序列大小分別為1 645 bp和1 091 bp。根據(jù)200 kb的核苷酸中含有3個(gè)以上基因即為1個(gè)基因簇[26],3號(hào)染色體上的4個(gè)基因形成1個(gè)基因簇,其他染色體上未形成基因簇。
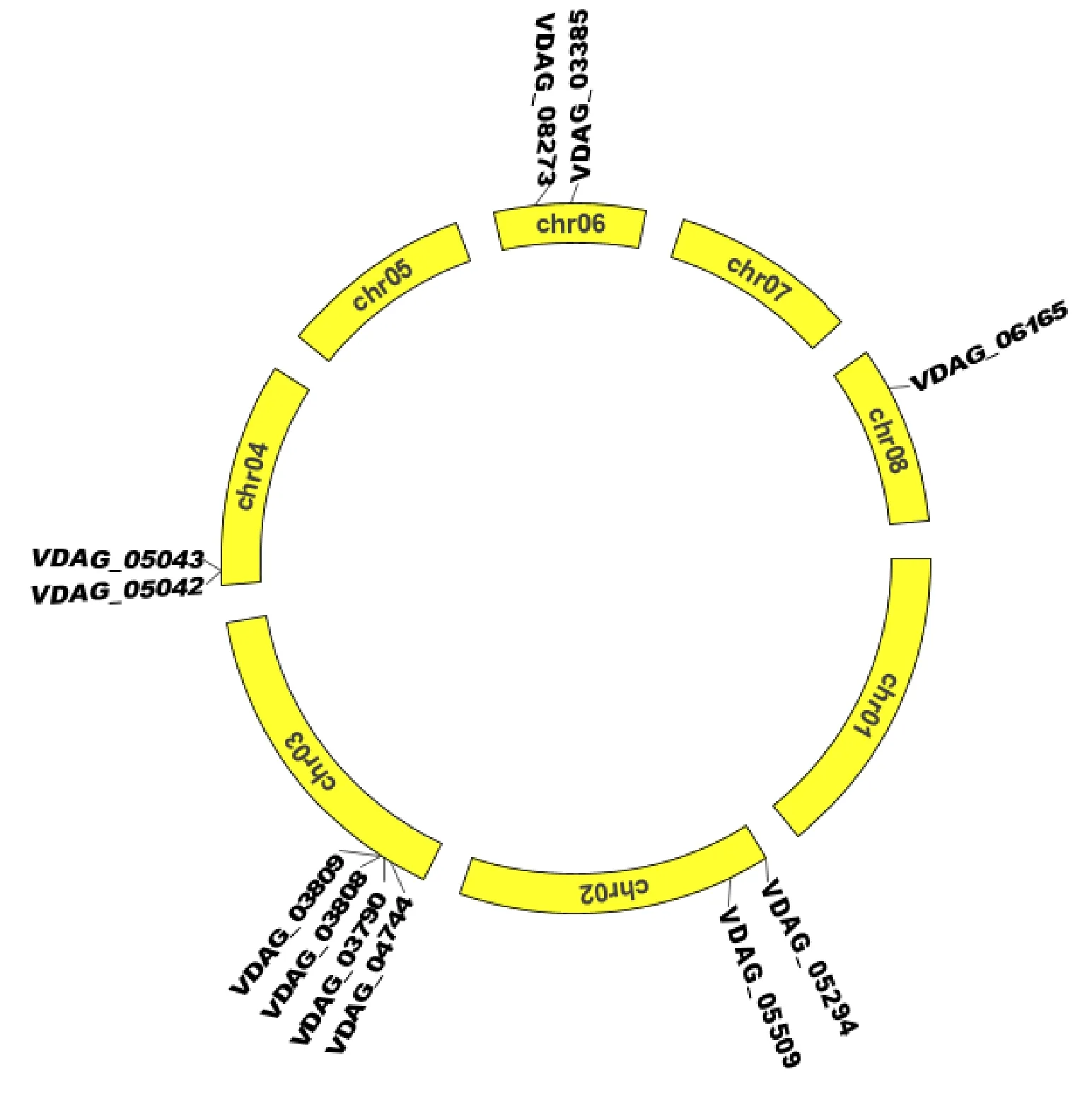
圖5 大麗輪枝菌β-1,4-內(nèi)切木聚糖酶基因在染色體上的分布Fig.5 Chromosomal distribution of Endo-β-1,4-xylanase genes in V.dahliae
2.6 β-1,4-內(nèi)切木聚糖酶基因?qū)Σ煌垢忻藁ㄆ贩N根系分泌物的響應(yīng)分析
抗病品種根系分泌物對(duì)病菌的孢子萌發(fā)和菌絲生長(zhǎng)有一定的抑制作用,而感病品種根系分泌物則能刺激病菌生長(zhǎng)[27-28]。根據(jù)不同抗感棉花品種根系分泌物培養(yǎng)6 h和12 h的大麗輪枝菌的轉(zhuǎn)錄組測(cè)序結(jié)果,獲得β-1,4-內(nèi)切木聚糖酶基因培養(yǎng)后每個(gè)時(shí)間點(diǎn)的FPKM值,用HemI軟件繪制基因在不同抗感棉花根系分泌物中的表達(dá)熱圖。根據(jù)β-1,4-內(nèi)切木聚糖酶基因在不同抗感根系分泌物中表達(dá)模式的差異(圖6),11個(gè)基因被劃分為3組,第3組基因(VDAG_03790、VDAG_03808、VDAG_03809、VDAG_05043和VDAG_06165)的表達(dá)量在感病品種根系分泌物培養(yǎng)的大麗輪枝菌(VDX6和VDX12)中明顯較高,而在耐病品種根系分泌物培養(yǎng)的大麗輪枝菌(VDZ6和VDZ12)和水培養(yǎng)的大麗輪枝菌(VDW6和VDW12)中表達(dá)量較低。結(jié)合5個(gè)基因的FPKM折線圖(圖7)可見,VDAG_03790在感病品種根系分泌物培養(yǎng)6 h的樣本中表達(dá)量明顯升高;VDAG_03808在感病品種根系分泌物培養(yǎng)6 h和12 h的樣本中表達(dá)量均較高,VDAG_03809、VDAG_05043和VDAG_06165在感病品種根系分泌物培養(yǎng)12 h的樣本中表達(dá)量明顯升高。而5個(gè)基因在耐病品種根系分泌物和水培養(yǎng)的樣本表達(dá)量未見明顯升高。上述分析表明5個(gè)基因均對(duì)感病品種根系分泌物產(chǎn)生響應(yīng),暗示它們與大麗輪枝菌的致病性相關(guān)。

紅色和藍(lán)色分別代表基因表達(dá)水平的高低
Red and blue bands represent high and low gene expression levels, respectively
圖6 β-1,4-內(nèi)切木聚糖酶基因在不同抗感棉花
根系分泌物培養(yǎng)的大麗輪枝菌中的表達(dá)熱圖
Fig.6 Heat map of Endo-β-1,4-xylanase genes
inV.dahliaecultured root exudates
from different resistant cottons
3 結(jié)論與討論
糖苷水解酶(glycoside hydrolase,GH)亦稱糖苷酶,是作用于各種糖苷或寡糖使糖苷鍵水解的酶的總稱。CAZy(Carbohydrate-Active enzymes Database)數(shù)據(jù)庫(kù)以糖苷水解酶與唯一性底物特異性結(jié)合原理,將糖苷水解酶家族分為130個(gè)家族[29-30]。內(nèi)切木聚糖酶主要存在于兩個(gè)離散的序列家族第10和11家族[31]。本研究通過對(duì)大麗輪枝菌中11個(gè)β-1,4-內(nèi)切木聚糖酶基因進(jìn)行生物信息學(xué)分析發(fā)現(xiàn),5個(gè)基因含有Glyco_hydro11結(jié)構(gòu)域,與其他物種中11家族成員聚為一類,屬于糖苷水解酶11家族;4個(gè)基因含有Glyco_hydro10結(jié)構(gòu)域,與其他物種中10家族成員聚為一類,屬于10家族;2個(gè)基因含有Glyco_hydro43結(jié)構(gòu)域,與其他物種中43家族成員聚為一類,屬于43家族。
蛋白結(jié)構(gòu)域分析還發(fā)現(xiàn)VDAG_06165中含有一個(gè)fCBD結(jié)構(gòu)域,該結(jié)構(gòu)域被認(rèn)為存在于大多數(shù)真菌纖維素酶和半纖維素酶中[32]。CBD結(jié)構(gòu)域在木聚糖酶降解天然植物細(xì)胞壁木聚糖中起著重要的作用[32-33],它是由33~36個(gè)氨基酸組成的具有高度保守性的區(qū)域。由于VDAG_06165含有fCBD結(jié)構(gòu)域,推測(cè)該基因可能與降解木聚糖有關(guān)。VDAG_03808中含有一個(gè)CBM結(jié)構(gòu)域,該結(jié)構(gòu)域是碳水化合物結(jié)合模塊(CBMS),通常存在于較長(zhǎng)的蛋白質(zhì)序列中,與碳水化合物活性酶的催化模塊(例如糖苷水解酶和多糖裂合酶)共存,在那里它們可增強(qiáng)酶模塊的催化活性[34]。CBM通過使酶接近纖維素表面來影響酶的活性[30,34]。

VDX表示β-1,4-內(nèi)切木聚糖酶基因在感病棉花品種根系分泌物中表達(dá)量;VDZ表示β-1,4-內(nèi)切木聚糖酶基因在抗病棉花品種根系分泌物中表達(dá)量;VDW表示β-1,4-內(nèi)切木聚糖酶基因在水培養(yǎng)中表達(dá)量
VDX represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with root exudates from susceptible cultivars;VDZ represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with root exudates from tolerance cultivars;VDW represents the expression level of Endo-β-1,4-xylanase genes inV.dahliaecultured with water
圖7 β-1,4-內(nèi)切木聚糖酶基因在不同抗感棉花品種根系分泌物培養(yǎng)的
大麗輪枝菌中FPKM值的變化趨勢(shì)圖
Fig.7 Change trend of FPKM value of Endo-β-1,4-xylanase genes inV.dahliae
cultured with root exudates from different resistant cottons
糖苷水解酶43家族成員均含有Glyco_hydro43結(jié)構(gòu)域,催化果膠降解的果膠酶一般含有該結(jié)構(gòu)域[25]。本研究中VDAG_03808和VDAG_05509均含有糖苷水解酶43家族結(jié)構(gòu)域,推測(cè)這兩個(gè)基因可能參與果膠的降解。在所有含有糖苷水解酶43主結(jié)構(gòu)域的蛋白質(zhì)中,69%的蛋白質(zhì)還含有一個(gè)信號(hào)肽,將翻譯后的蛋白質(zhì)導(dǎo)向細(xì)胞質(zhì)外。VDAG_03808中無信號(hào)肽;VDAG_05509含有1~19 bp長(zhǎng)度分泌信號(hào)肽,推測(cè)該基因編碼的分泌蛋白在細(xì)胞外發(fā)揮作用。
根系分泌物是植物根系在生長(zhǎng)過程中釋放到介質(zhì)中的全部有機(jī)和無機(jī)物質(zhì)。大麗輪枝菌通過根系入侵棉花,因此根系分泌物的生物學(xué)效應(yīng)對(duì)大麗輪枝菌的成功侵染至關(guān)重要[28,35-36]。棉花根系分泌物富含氨基酸和糖類,抗病品種和感病品種的根系分泌物成分有明顯差異。與抗病棉花品種根系分泌物相比,感病品種根系分泌物中富含天冬氨酸、蘇氨酸、賴氨酸和脯氨酸等大量氨基酸,葡萄糖、果糖和蔗糖含量明顯較高[37]。抗病品種根系分泌物抑制大麗輪枝菌的生長(zhǎng),而感病品種根系分泌物促進(jìn)其生長(zhǎng)[22,28]。目前棉花根系分泌物對(duì)基因表達(dá)的影響研究仍較少,VdRGS1在根系分泌物處理的大麗輪枝菌中的表達(dá)量明顯升高,基因敲除和回補(bǔ)試驗(yàn)發(fā)現(xiàn)該基因與大麗輪枝菌孢子生產(chǎn)、菌絲發(fā)育、微菌核的形成和致病性相關(guān)[13]。本研究分析不同抗感棉花品種根系分泌物對(duì)11個(gè)β-1,4-內(nèi)切木聚糖酶基因的表達(dá)的影響。
植物病原真菌可以產(chǎn)生一系列細(xì)胞壁降解酶,以促進(jìn)感染和定植,包括纖維素酶、半纖維素酶、果膠酶等。近年來,研究人員已利用基因敲除手段對(duì)大麗輪枝菌中細(xì)胞壁降解相關(guān)基因的功能進(jìn)行研究,但由于這些基因都是以基因家族的形式存在,導(dǎo)致基因的功能研究未獲得確定性的結(jié)果。因此,確定基因家族中對(duì)致病性起關(guān)鍵作用的基因?qū)τ诖篼愝喼虏》肿訖C(jī)制的解析是十分必要的。本研究通過制作11個(gè)基因在不同抗感棉花根系分泌物中的表達(dá)熱圖和部分基因的表達(dá)折線圖,發(fā)現(xiàn)5個(gè)β-1,4-內(nèi)切木聚糖酶基因在感病品種根系分泌物培養(yǎng)的大麗輪枝菌中明顯上調(diào),而在耐病品種根系分泌物和水培養(yǎng)的樣本中未明顯上調(diào),表明它們對(duì)感病品種的根系分泌物有明顯的響應(yīng),暗示它們可能與大麗輪枝菌的致病性相關(guān),這些基因可用作今后利用基因敲除技術(shù)研究基因功能與大麗輪枝菌致病分子機(jī)制的目標(biāo)基因。
