miR-100-5p下調mTOR抑制前列腺癌細胞的增殖、遷移與侵襲
2020-06-06 06:49:22李蘇亮袁小華閆紅林西安醫學院第一附屬醫院輸血科西安70077西安醫學院第一附屬醫院消化內科通訊作者mailyanhonglin66663com
山西醫科大學學報 2020年5期
葉 蕓,李蘇亮,袁小華,劉 冰,相 蓮,閆紅林(西安醫學院第一附屬醫院輸血科,西安 70077;西安醫學院第一附屬醫院消化內科;通訊作者,E-mail:yanhonglin666@63.com)
最新統計數據顯示,2016年美國前列腺癌新發病161 360例,占男性腫瘤新發病例19%;死亡人數26 730例,占男性腫瘤死亡病例8%[1]。隨著我國人口老齡化的發展以及飲食結構的改變,前列腺癌的發病率逐年升高[2]。前列腺癌在全世界范圍內已成為備受關注的公共健康問題。microRNA(miRNA)是一類由21-24個核苷酸組成的高度保守的非編碼單鏈RNA,通過與靶基因不完全或完全互補結合從而調控靶基因的表達,在機體基因調控網絡中發揮著重要的作用[3,4]。近年來研究發現miRNA作為癌基因或抑癌基因在惡性腫瘤發生進展中發揮重要作用[5]。大量研究證實miRNA在前列腺癌發生及發展中表達失調,且前列腺癌患者與正常人群之間存在差異表達[6,7]。miR-100-5p是miRNA家族重要成員之一,位于染色體11q24.1上,呈高度保守性。miR-100-5p在包括前列腺癌在內許多惡性腫瘤中表達異常[8],參與腫瘤細胞的增殖、遷移及侵襲等生物學行為,但作用機制尚不明確。本研究擬觀察miR-100-5p在前列腺癌細胞系中的表達變化以及其對細胞增殖、遷移及侵襲能力的影響,并探討其作用機制。
1 材料與方法
1.1 主要試劑
miR-100-5p mimics、陰性對照NC-mimics(購自廣州銳博生物公司),Trizol試劑、LipofectamineTM2000購自美國Invitrogen公司,RNA逆轉錄試劑盒、qRT-PCR試劑盒購自大連寶生生物公司,Transwell小室購自美國Corning公司,iBlot半干電轉印試劑盒購自Invitrogen公司,兔抗人單克隆抗體mTOR及GAPDH購自Abcam公司。
1.2 細胞系
人前列腺癌細胞LNCaP和人前列腺上皮細胞RWPE-1購自ATCC(Manassas, VA, USA)。RWPE-1細胞使用含有L-谷氨酰胺的F12K細胞培養基(Gibco BRL Co. Ltd., USA);LNCaP細胞使用 Roswell Park Memorial Institute-1640培養基(HyClone, Logan, UT, USA)。所有培養基均補充青霉素100 U/ml和鏈霉素100 U/ml(HyClone, Logan, UT, USA)和10% exosomes-free FBS(Gibco BRL Co. Ltd., USA)。均置于37 ℃,5% CO2的飽和濕度恒溫箱中常規培養,細胞用0.05% EDTA消化傳代,取生長狀態良好的對數生長期細胞進行后續實驗。
1.3 qRT-PCR檢測miR-100-5p及mTOR的表達
按照TRIzol試劑說明書提取前列腺癌細胞LNCaP的總RNA,鑒定純度和含量后,逆轉錄合成cDNA。cDNA合成參照逆轉錄試劑盒說明書操作。以cDNA為模板,按照qRT-PCR試劑盒說明書配置反應體系進行PCR反應,miR-100-5p基因PCR反應條件:94 ℃ 2 min;40個循環(94 ℃ 20 s,55 ℃ 30 s);mTOR基因PCR反應條件:94 ℃ 2 min;40個循環(94 ℃ 30 s,58 ℃ 30 s,72 ℃ 30 s)。miR-100-5p以U6作為內參基因,mTOR以GAPDH作為內參基因,相對表達水平采用2-ΔΔCt法計算,實驗重復3次。
1.4 細胞轉染
取對數生長期LNCaP細胞分為NC-mimics組和miR-100-5p mimics組,接種于6孔細胞培養板,常規調節下培養,細胞融合度達60%-70%時進行細胞轉染操作。嚴格按LipofectamineTM2000說明書操作分別將NC-mimics和miR-100-5p mimics轉染至LNCaP細胞,轉染后24 h,qRT-PCR檢測miR-100-5p的表達,確定轉染效率后再進行后續細胞實驗。
1.5 CCK-8檢測細胞的增殖活性
收集轉染后的細胞按5 000/孔的密度分別種于96孔板,每組設3個復孔,每24 h加入10 μl CCK-8反應液37 ℃繼續孵育,在Multiskan FC酶標儀上測定450 nm波長處各孔不同時點的吸光度(OD)值,繪制增殖曲線。
1.6 細胞劃痕實驗檢測細胞遷移能力
取轉染后24 h的各組細胞,接種培養于6孔板中,孔板底部預先畫好標記線,待細胞長到完全融合后,用10 μl的滅菌槍頭垂直培養孔底面中央用力劃直線沿直線劃痕,PBS沖洗2次后繼續培養,記錄劃痕后0 h和24 h顯微鏡劃痕愈合情況,劃痕愈合率(%)=[(0 h劃痕寬度-24 h劃痕寬度)/0 h劃痕寬度]×100%,實驗重復3次。
1.7 Transwell侵襲實驗
基質膠均勻涂抹Transwell小室上室,過夜成膜。次日收集轉染24 h的LNCaP細胞以無血清培養基重懸制備單細胞懸液,取200 μl含2×105個細胞的單細胞懸液加入Transwell小室上室,小室下室加入500 μl含10%胎牛血清的DMEM培養基,繼續培養24 h后取出Transwell小室,棉簽輕輕擦棄小室上室的細胞,4%多聚甲醛固定,染色,風干,顯微鏡下觀察,并隨機取5個高倍視野計算穿膜細胞數。實驗重復3次。
1.8 Western blot檢測mTOR的表達
取對數生長期LNCaP細胞,常規提取細胞總蛋白,以BCA法定量蛋白濃度。每個樣本取200 μl樣品,10% SDS-PAGE分離,將電泳產物轉至PVDF膜,5%封閉液4 ℃封閉1 h后,TBS緩沖液漂洗3遍,加入一抗mTOR和GAPDH(1∶1 000),4 ℃孵育過夜,TBS緩沖液漂洗3遍,加入二抗(1∶5 000),室溫孵育45 min后,ECL顯影液顯影,Quality One軟件分析條帶灰度,以mTOR與GAPDH灰度的比值表示相對表達量。實驗重復3次。
1.9 統計學方法
SPSS 20.0進行數據分析,計量資料以均數±標準差表示,采用Mann-WhitneyU檢驗分析非參數數據;P<0.05為差異有統計學意義。
2 結果
2.1 轉染前后miR-100-5p在LNCaP和RWPE-1細胞中的表達
轉染前qRT-PCR結果顯示miR-100-5p在前列腺癌細胞LNCaP的表達水平較RWPE-1細胞低(P<0.01,見圖1A)。miR-100-5p mimics轉染后LNCaP細胞中miR-100-5p的表達明顯高于NC-mimics組(P<0.01,見圖1),表明miR-100-5p mimics轉染效率較高。

與各自對照組相比較,**P<0.01
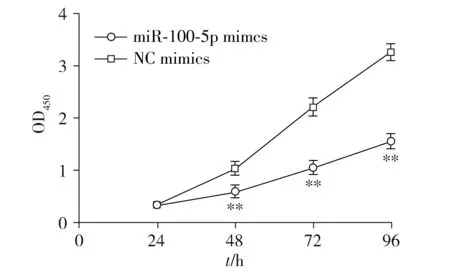
2.2 轉染后LNCaP細胞的增殖活性
CCK-8增殖實驗顯示LNCaP細胞轉染mimics組48,72,96 h時OD值明顯低于NC-mimics組(P<0.01,見圖2)。
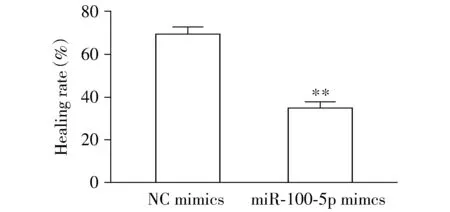
2.3 轉染后LNCaP細胞的遷移能力
與NC-mimics組比較,miR-100-5p mimics組轉染后細胞的劃痕愈合率明顯下降(P<0.01,見圖3,4),提示轉染后LNCaP細胞的遷移能力明顯降低。
2.4 轉染后LNCaP細胞的侵襲能力
Transwell侵襲實驗結果顯示,miR-100-5p mimics組穿膜細胞數明顯低于NC-mimics組(P<0.01,見圖5),提示轉染后細胞的侵襲能力明顯下降。
2.5 生物學信息法預測miR-100-5p的靶基因
應用生物學信息法預測miR-100-5p可能靶基因,查閱miRand(http://www.microrna.org),預測mTOR可能是miR-100-5p的作用靶基因,miR-100-5p與mTOR的3′UTR區存在種子序列互補的結合位點(見圖6)。
與NC-mimics組比較,**P<0.01
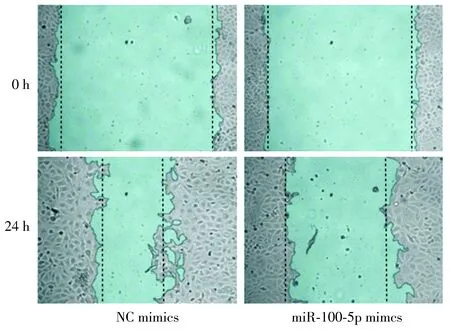
圖3 細胞劃痕實驗檢測細胞的遷移能力Figure 3 Cell migration ability detected by cell scratch experiment
與NC-mimics組相比較,**P<0.01
2.6 轉染后LNCaP細胞的mTOR mRNA和蛋白的表達
qRT-PCR結果顯示miR-100-5p mimics轉染LNCaP細胞mTOR mRNA的表達明顯低于轉染NC-mimics細胞(P<0.01,見圖7)。Western blot 顯示100-5p mimics轉染后LNCaP細胞mTOR 蛋白的表達明顯低于轉染NC-mimics細胞(P<0.01,見圖7)。

圖5 轉染后LNCaP細胞侵襲能力的改變Figure 5 Invasion ability of cells detected by Transwell invasion assay
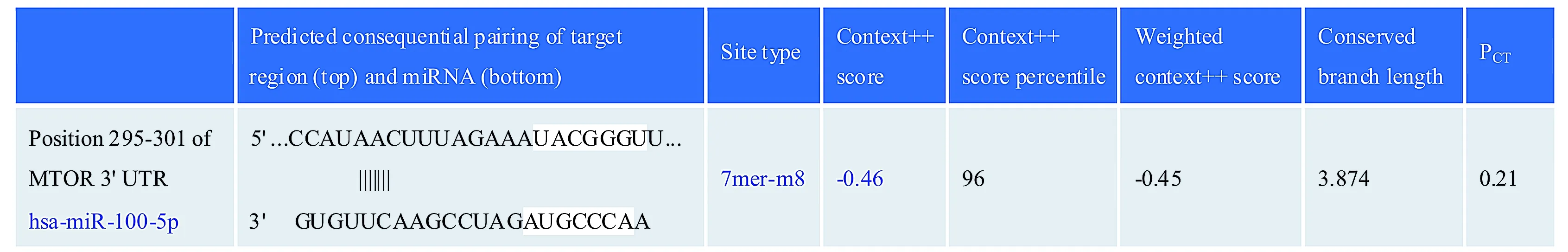
圖6 miR-100-5p與mTOR的3′UTR區 的結合序列Figure 6 The binding sequence of miR-100-5p with the 3′UTR region of mTOR
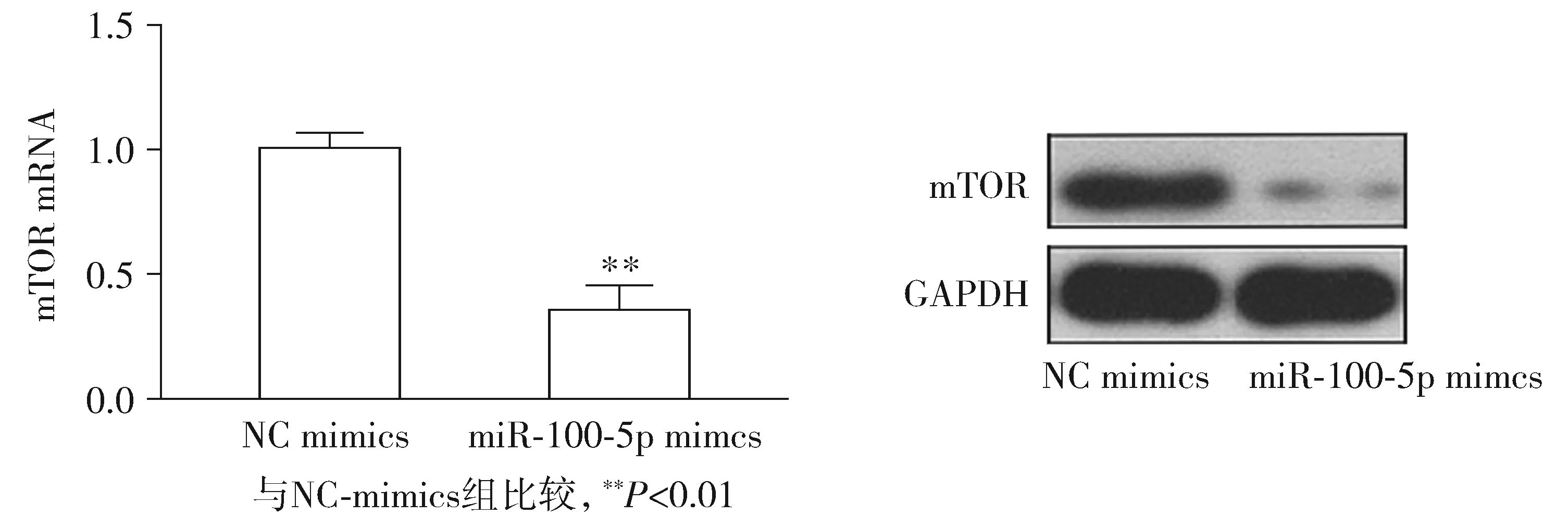
圖7 轉染后LNCaP細胞中mTOR mRNA和蛋白水平的表達Figure 7 Expression of mTOR mRNA and protein in LNCaP cells after transfected with miR-100-5p mimics
3 討論
miRNA是一類19-24 nt左右的ncRNA。miRNA可與多個靶mRNA3′UTR結合并調控基因表達,導致靶基因異常表達[9,10]。miRNA的異常表達與腫瘤的發生發展存在關聯,研究證實miRNA在腫瘤組織中差異性表達,可以作為腫瘤早期檢測、分型及預后的生物標志物[11,12]。研究發現miR-100-5p等在前列腺癌中表達下調,被認為是抑癌基因在前列腺癌的發生進展中發揮重要作用[13]。有報道稱,miR-100-5p的缺失導致AGO2表達水平上調,進而發生癌細胞的遷移、侵襲、EMT,從而促進前列腺癌的轉移[14]。本研究通過qRT-PCR檢測前列腺癌細胞系LNCaP和人正常前列腺上皮細胞系RWPE-1中miR-100-5p的表達,miR-100-5p在LNCaP細胞中表達明顯降低,提示miR-100-5p在前列腺癌的發生發展中可能也起著抑癌基因的作用。細胞無限增殖、侵襲與轉移是惡性腫瘤的基本生物學特性,在對多種惡性腫瘤的研究中均發現miR-100-5p能夠抑制腫瘤細胞的增殖、遷移及侵襲等腫瘤生物學行為[15,16]。
為了進一步研究miR-100-5p對LNCaP細胞增殖、侵襲和遷移能力的影響,通過脂質體轉染法將miR-100-5p mimics轉染至LNCaP細胞,成功上調細胞中miR-100-5p的表達;CCK-8增殖實驗、細胞劃痕實驗和Transwell侵襲實驗結果顯示miR-100-5p的上調能夠明顯抑制腫瘤細胞的增殖活性、遷移和侵襲能力,進一步證實miR-100-5p作為抑癌基因能夠抑制PCa的進展。
miR-100-5p抑制腫瘤細胞增殖、侵襲及遷移能力的下游分子通路尚不完全明確,為了研究miR-100-5p調控LNCaP細胞的下游靶基因,應用生物學信息法預測mTOR可能是miR-100-5p的作用靶基因,因為miR-100-5p與mTOR的3′UTR區存在種子序列互補的結合位點。mTOR是一種絲氨酸/蘇氨酸蛋白激酶,其通過調控蛋白質合成參與生理進程和病理反應,并與癌癥的發病機制有關[17]。mTOR信號通路主要調控參與腫瘤發生發展的細胞增殖和代謝,是僅次于p53通路的人類癌癥中第二大通路[18]。本研究中,應用qRT-PCR和Western blot發現,上調LNCaP細胞中miR-100-5p表達后,細胞中mTOR mRNA和蛋白的表達水平明顯下降,提示miR-100-5p在前列腺癌的發生發展中的作用與靶向調控mTOR基因有關,上調miR-100-5p的表達能夠明顯抑制mTOR基因的表達。mTOR 是自噬信號通路中的主要靶標,在細胞生長、增殖、分化、細胞周期調控等方面起重要作用。基于本研究結果推測miR-100-5p可能通過靶向mTOR誘導腫瘤細胞的自噬。
綜上所述,本研究結果表明,miR-100-5p在前列腺癌細胞系LNCaP中低表達,且miR-100-5p能夠抑制LNCaP細胞的增殖、遷移及侵襲能力,其機制與下調mTOR基因的表達有關,可能成為未來PCa靶向治療的分子靶點。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
發明與創新(2022年30期)2022-10-03 08:40:56
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:02
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:00
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:00
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年7期)2019-01-06 09:27:34
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
發明與創新(2016年38期)2016-08-22 03:02:52
