異育銀鯽內參基因的篩選
2020-06-07 13:14:30費越越南星羽高鐘元
水產科學 2020年3期
關鍵詞:分析
費越越,南星羽,余 路,羅 揚,高鐘元,許 丹,3
( 1.上海海洋大學,國家水生動物病原庫,上海 201306; 2.上海海洋大學,農業農村部水生種質資源重點實驗室,上海 201306; 3.上海海洋大學,國家漁業科學教育示范中心,上海 201306 )
實時熒光定量PCR(qRT-PCR)是一種實時監控PCR擴增產物并進行解析的定量方法,因其靈敏度高、操作簡單等優點,目前已在基因表達檢測中廣泛應用[1]。在常用的相對定量PCR試驗中,為準確獲得目的基因的相對表達量,通常引入合適的內參基因來校正RNA質量和反應條件等存在的誤差。理想的內參基因應該在不同組織、不同發育階段和不同處理條件下均保持相對恒定的表達水平[2]。試驗中常選擇管家基因作為內參基因,如甘油醛-3-磷酸脫氫酶(GAPDH)、β-肌動蛋白(β-actin)、18S核糖體RNA(18S rRNA)和延伸因子(EF-1α)。然而,有研究報道稱,絕對穩定的基因并不存在,管家基因也只是在特定條件下表現出相對穩定的表達[3],如鱖魚(Sinipercachuasti)胚胎發育階段β-actin、18S rRNA和GAPDH基因的穩定性研究表明,18S rRNA在不同發育階段表達水平有顯著差異[4]。達氏鱘(Acipenserdabryanus)14種不同組織中EF-1α、β-actin和GAPDH基因的表達量研究表明,GAPDH基因表達有顯著差異[5]。因此,即便是管家基因也要在特定條件下進行特定分析。為了提高qRT-PCR數據結果準確性,通常需要根據試驗處理條件選擇合適的內參基因。
異育銀鯽(Carassiusauratusgibelio)是以興國紅鯉(Cyprinuscarpiovar.singuonensis)為父本,天然雌核發育的方正銀鯽(C.auratusgibelio)為母本,經人工授精和異精雌核發育而獲得的子代,因其生長迅速、適應力強、肉質鮮美且易于養殖,頗受人們喜愛[6]。據統計,目前,我國養殖的異育銀鯽產量超過3.0×107t,在水產養殖中占有非常重要的地位[7-8]。但是近幾年來,大規模的集約化養殖導致其病害問題日益嚴重[9]。其中由鯉皰疹病毒Ⅱ型(CyprinidherpesvirusⅡ,CyHV-2)引發的造血器官壞死癥導致異育銀鯽大規模死亡,這給養殖戶造成巨大的經濟損失[10]。目前,針對鯉皰疹病毒Ⅱ型引發異育銀鯽大規模死亡的致病機理還不是十分清楚。Lu等[11]為研究鯉皰疹病毒Ⅱ型的致病機理選取感染后的腎臟組織進行轉錄組測序分析,利用qRT-PCR技術驗證并分析基因的表達情況;Xia等[12]為探究干擾素對造血器官壞死癥的防治作用,利用qRT-PCR技術研究干擾素相關基因的表達情況。然而利用qRT-PCR技術進行異育銀鯽內參基因系統篩選的研究國內外尚未見報道。在生物體中各組織均有其獨特的生理機能,研究基因在不同組織中的不同表達量,可為研究基因的功能提供參考[13]。據報道,異育銀鯽的腎臟和脾臟是鯉皰疹病毒Ⅱ型病毒富集最主要的組織,篩選出感染腎臟和脾臟在不同時間點均較穩定的內參基因可以為研究病毒致病機理獲得最準確的基因表達結果[14]。魚類細胞系的構建為魚類疾病的研究等提供強有力的基礎材料[15]。細胞水平qRT-PCR內參基因的研究將為確保基因表達分析的準確性提供依據,從而為細胞水平探究致病機理奠定基礎。
筆者選用GAPDH、EF-1α、18S rRNA和β-actin 4個管家基因,利用內參基因評估軟件geNorm[16]、Norm Finder[17]、Best Keeper[18]、Delta Ct[19]以及RefFinder[20]對這4個內參基因分別在健康異育銀鯽的不同組織、鯉皰疹病毒Ⅱ型感染腎臟、脾臟以及尾鰭細胞在不同時間點進行內參基因表達穩定性分析,并從異育銀鯽腎臟組織cDNA轉錄組文庫中篩選出差異表達的基因PIN1[21]在腎臟組織不同感染時間點,對4個候選內參基因的穩定性進行驗證。最終自4個候選內參基因中篩選出在健康異育銀鯽的不同組織、鯉皰疹病毒Ⅱ型感染腎臟、脾臟以及尾鰭細胞不同時間點均適用的內參基因。
1 材料與方法
1.1 試驗材料
健康異育銀鯽 [體長(10.1±0.2) cm,質量(90.2±0.12) g] 購于江蘇四大家魚原種場,在上海海洋大學動物房(23±0.5) ℃暫養,暫養期間每日投喂顆粒飼料1次(投喂量為魚體質量的3%)。經過7 d的暫養,隨機分為5組進行腹腔注射鯉皰疹病毒Ⅱ型病毒懸液(每組注射量為7×103.9±0.22pfu/mL)[22],對照組注射1 mL磷酸緩沖鹽溶液。注射后,分別在0、6、12、24 h和120 h時間點取6尾魚,解剖取腦、脾臟、腎臟、肌肉、鰓、腸、肝臟和心臟組織塊,迅速放置于液氮中速凍后暫存于-80 ℃冰箱中。
本試驗的尾鰭細胞[22]和鯉皰疹病毒Ⅱ型由國家水生動物病原庫實驗室保存[23]。尾鰭細胞用規格為T75培養瓶培養18瓶,傳代48 h(細胞融合度至80%~90%),棄除原來培養基(10%FBS,M199培養基),加入2 mL鯉皰疹病毒Ⅱ型病懸液(7×103.9±0.22pfu/mL)[22],對照組加入2 mL磷酸緩沖鹽溶液孵育2 h之后,注入8 mL新鮮培養基(2%FBS,M199培養基),放置27 ℃培養箱。分別在0、1、2、3、4 d和5 d各時間點收集3瓶細胞,放置冰箱-80 ℃保存。
1.2 總RNA的提取和cDNA 的合成
健康異育銀鯽的不同組織、鯉皰疹病毒Ⅱ型感染異育銀鯽的腎臟、脾臟以及細胞不同時間點的總RNA均使用試劑盒TRIzol?Reagent (Thermo Scientific, 美國)提取。用1%瓊脂糖凝膠電泳檢測RNA完整性。使用NanoDrop?2000c核酸分析儀(Thermo Scientific, 美國)檢測RNA的含量及純度,稀釋后保持其質量濃度為500~1000 ng/μL。分別從不同處理條件下的樣品中取2 μg RNA,使用Recombinant DNase Ⅰ(RNase-free) (TaKaRa)操作進行去DNA處理,使用PrimeScriptTMⅡ 1st Strand cDNA Synthesis Kit(TaKaRa)反轉錄成cDNA,存儲于-20 ℃備用。
1.3 引物來源
自異育銀鯽腎臟組織轉錄組數據庫中獲得GAPDH基因的部分序列,用Primer 5.0設計引物用于qRT-PCR(表1)。另外β-actin[24]、18S rRNA[25]、EF-1α[26]、PIN1[21]基因的引物序列來源于已發表的文章。
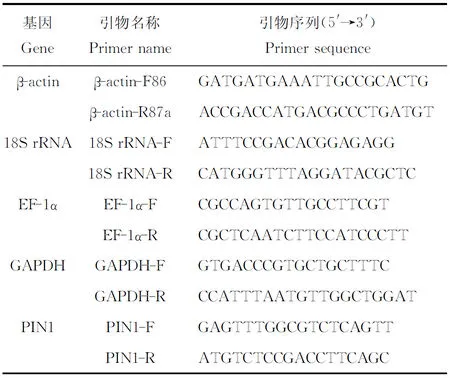
表1 4種候選內參基因的引物信息
1.4 qRT-PCR試驗
qRT-PCR使用 TB GreenTMPremix Ex TaqTMⅡ (Tli RNaseH Plus)試劑盒(TaKaRa)按說明書操作。反應在BIO RAD CFX96TMReal-Time System定量PCR儀完成,25 μL PCR擴增反應體系: TB GreenTMPremix Ex TaqTMⅡ (2×) 12.5 μL,cDNA (<100 ng) 2.0 μL,ddH2O 8.5 μL,PCR Forward/Reverse Primer(10 μmol/L) 1 μL,每個樣品重復測定3次。實時熒光定量PCR反應條件為95 ℃預變性30 s;95 ℃變性5 s,60 ℃復性30 s,共40個循環。
1.5 數據分析
試驗選用geNorm、Norm Finder、Best Keeper和Delta Ct對qRT-PCR產生的原始Ct值進行數據分析。運用Ref Finder對4種軟件的分析結果進行綜合排名,最終給出一個穩定性排名。
1.6 內參基因的驗證
取感染鯉皰疹病毒Ⅱ型病毒的異育銀鯽在0、6 h和72 h時的腎臟組織提取RNA(每個時間點3個生物學重復),為使研究更能區分內參基因的穩定性,選取已驗證在病毒感染不同時間點的腎臟組織表達差異較大的PIN1[21]基因為目的基因,分別以EF-1α、β-actin、18S rRNA和GAPDH為內參基因,按照1.3中qRT-PCR試驗步驟進行操作,引物見表1。內參基因驗證的相對表達量用平均值±標準誤來表示,使用SPSS 19.0進行單因素方差分析,P<0.01表示具有極顯著性差異。
2 結 果
2.1 引物的特異性評估
cDNA初始模板質量濃度控制在200 ng/μL,以10倍梯度稀釋的對數函數與循環閾值(Ct值)作標準曲線。計算擴增效率,r2變化為0.993~0.997。所有基因的溶解曲線均為明顯單一峰(圖1),且樣品PCR重復性較好,表明引物特異性較好,不存在引物二聚體。
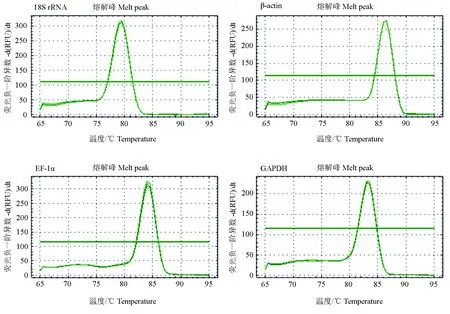
圖1 4對引物的熔解曲線
2.2 健康的異育銀鯽不同組織候選內參基因的穩定性評估
檢測健康異育銀鯽8種組織(腦、脾臟、腎臟、肌肉、鰓、腸、肝臟和心臟)中4個候選內參基因的表達量,所得Ct值分布見圖2。不同組織中4個候選內參基因的Ct值為7.45~26.27,其中GAPDH表達水平受組織影響差異較大,最大值和最小值相差10.17個循環。18S rRNA的Ct值變化較小,最大值和最小值相差2.18個循環。18S rRNA的表達Ct值過小,這可能是因為18S rRNA是一種核糖體RNA,約占RNA總量的80%,呈現高豐度表達[27-28]。EF-1α和β-actin在這8種組織中穩定性較好,均穩定在17~18個循環。
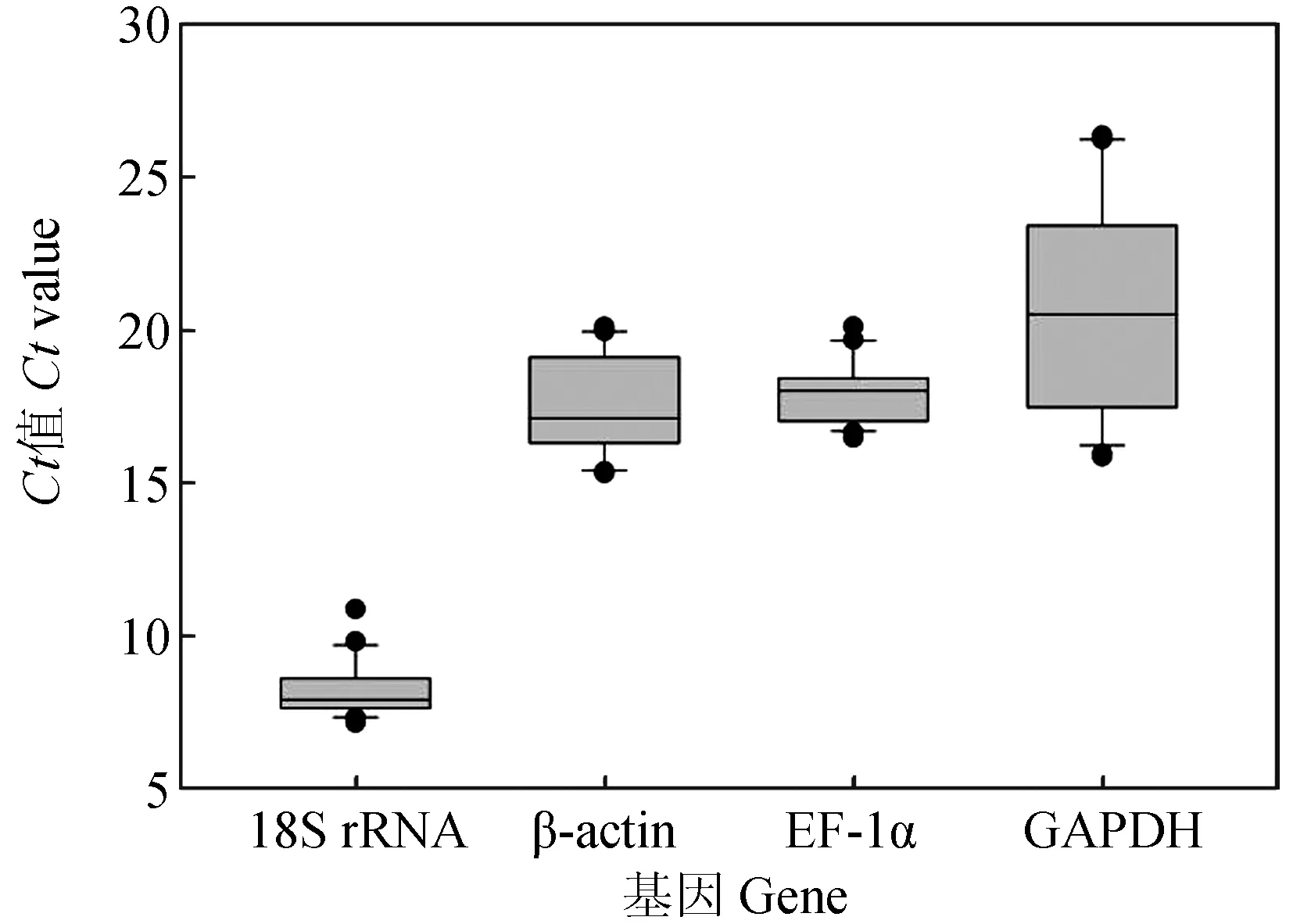
圖2 健康異育銀鯽的不同組織候選內參基因Ct值分布
健康異育銀鯽的不同組織內參基因穩定性排序結果顯示,Delta Ct不需要限定起始的總RNA的含量,可直接對同一處理條件下不同基因進行Ct值兩兩比較,獲得更精確的結果。Delta Ct對異育銀鯽的不同組織4個候選內參基因的評估結果顯示,各基因穩定性排序為EF-1α>β-actin>18S rRNA>GAPDH。
BestKeepe主要通過內參基因和目標基因表達量的配對相關系數、標準偏差和變異系數確定內參基因。其分析結果顯示,各基因穩定性排序為β-actin>EF-1α>GAPDH>18S rRNA。
Normfinder通過一個群體計算不同候選基因得到穩定值,這個數值越小基因則越穩定。Normfinder的分析結果與Delta Ct和BestKeeper的分析結果幾乎相同,與Delta Ct的分析結果完全一致,各基因穩定性排序為EF-1α>β-actin>18S rRNA>GAPDH。
geNorm與Normfinder算法類似,不同的是geNorm對候選內參基因進行兩兩比對計算得到各基因geNorm穩定值(亦稱M值),并通過geNorm穩定值對基因穩定性進行排序,geNorm穩定值越小,表達越穩定。其分析結果表明,EF-1α 和β-actin在異育銀鯽的不同組織中的表達同樣較穩定。由圖3可見,EF-1α和β-actin的geNorm穩定值均為0.304。
Ref Finder是一種綜合分析的軟件可以根據每個軟件的排名分配適當的權重并計算其權重的幾何平均值為最終的總排名。其分析結果顯示,各基因穩定性排序為EF-1α>β-actin>18S rRNA>GAPDH。
綜合4種軟件(Ref Finder)分析結果,結合geNorm穩定值以及4個內參基因的表達量Ct值,確定EF-1α和β-actin為健康異育銀鯽的不同組織穩定的內參基因。
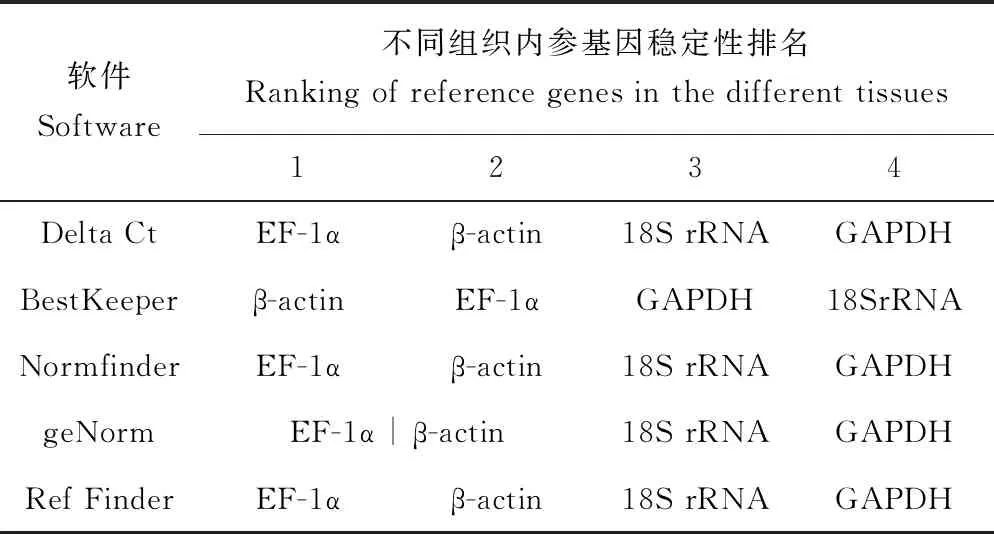
表2 健康異育銀鯽的不同組織內參基因穩定性排序
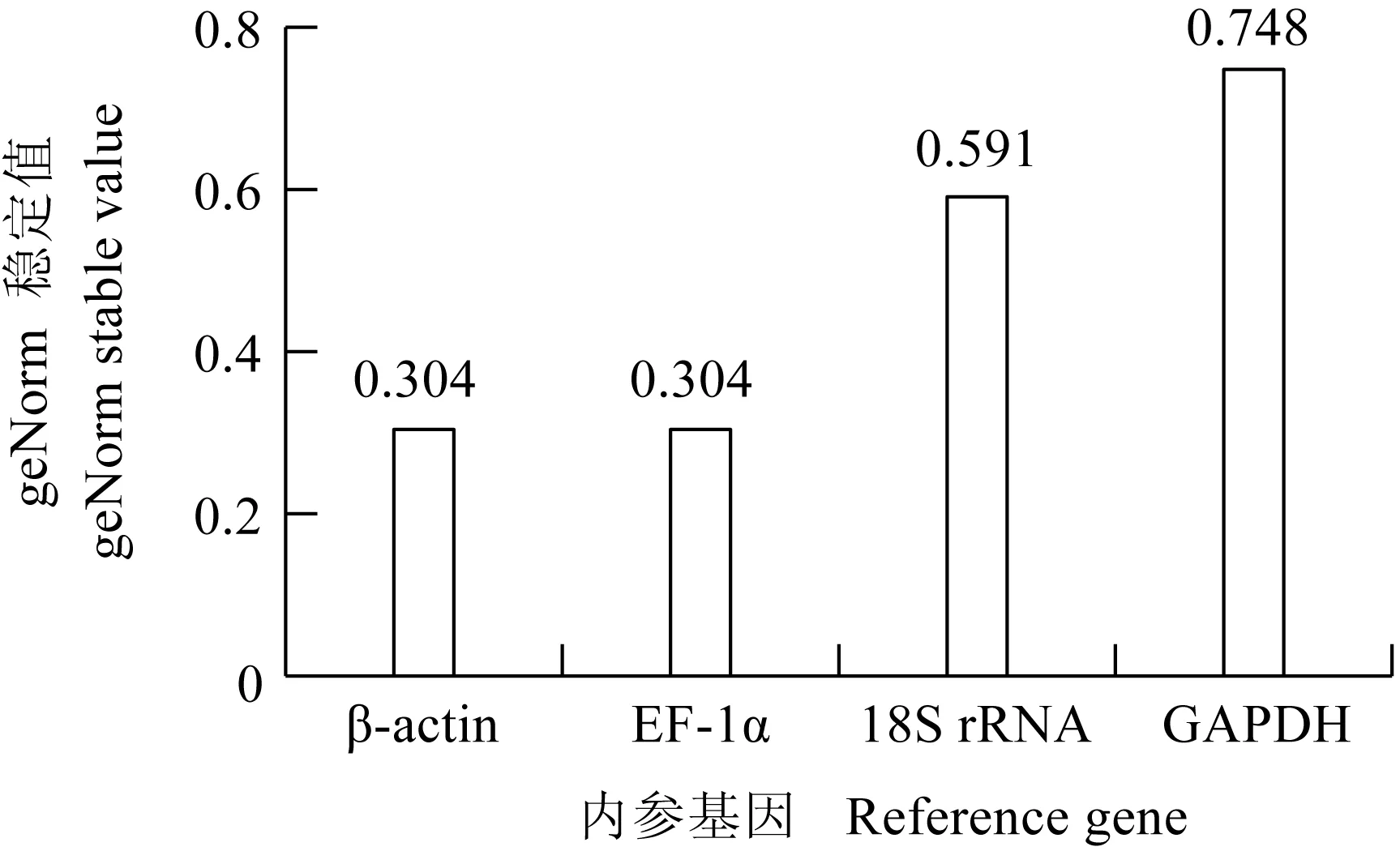
圖3 健康異育銀鯽的不同組織geNorm穩定值
2.3 鯉皰疹病毒Ⅱ型感染異育銀鯽腎臟組織在不同時間點候選內參基因的穩定性評估
檢測鯉皰疹病毒Ⅱ型感染異育銀鯽腎臟組織在不同時間點(0、6、12、24、48 h和120 h)4個候選內參基因的表達Ct值分布見圖4。4個候選內參基因的Ct值為8.79~24.58,其中GAPDH表達水平受感染時間影響差異較大,最大值和最小值相差3.21個循環。18S rRNA的Ct值變化較小,最大值和最小值相差1.53個循環。18S rRNA在不同感染時間點的表達量都過高,GAPDH在不同感染時間點的表達量都較β-actin和EF-1α的表達量低。相對而言,β-actin和EF-1α在4個候選內參基因中表達量較穩定。
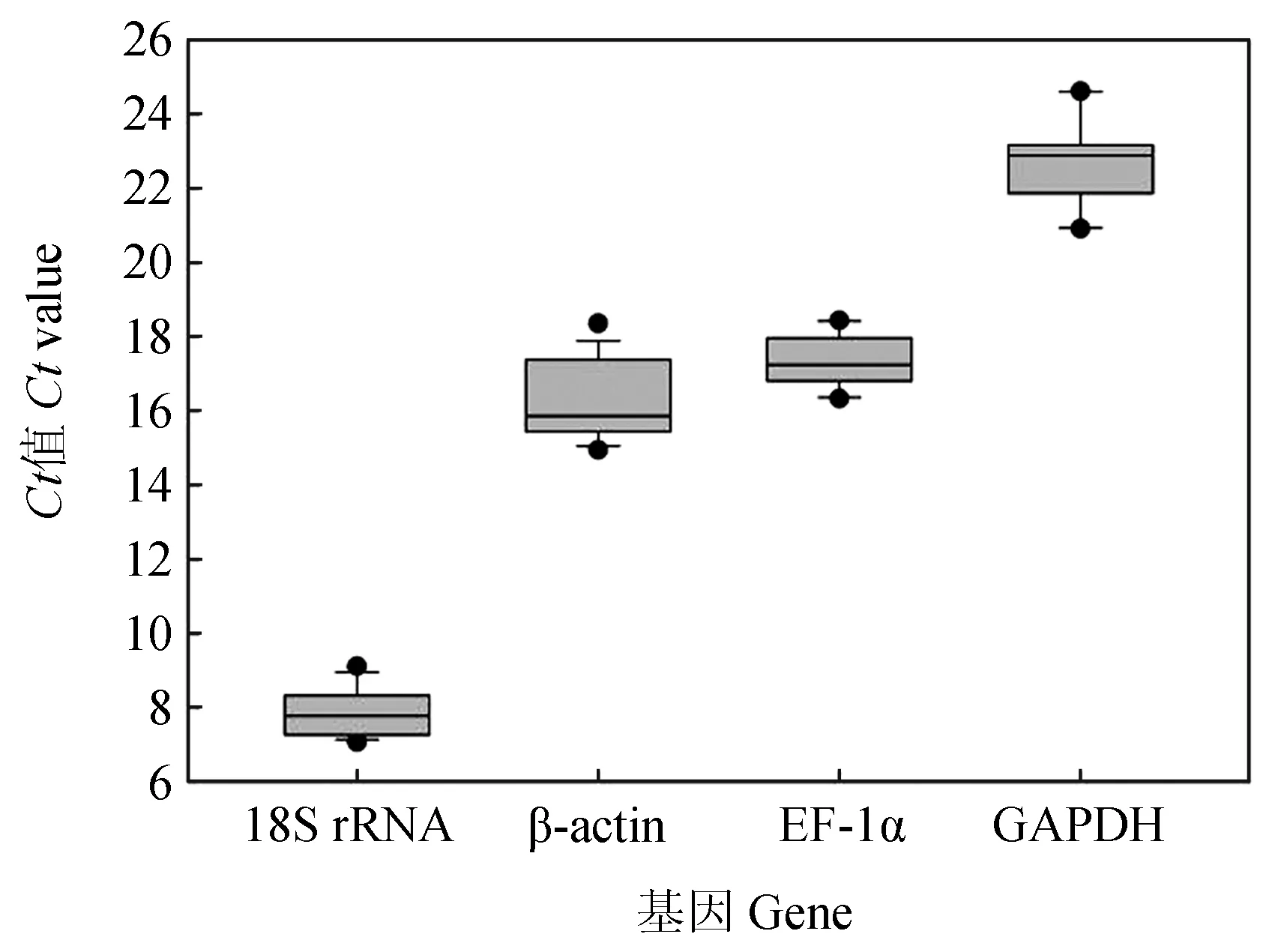
圖4 CyHV-2感染異育銀鯽腎臟組織不同時間點候選內參基因Ct值分布
Delta Ct的分析結果與Normfinder的分析結果一致,為β-actin>18S rRNA>GAPDH>EF-1α。BestKeeper的分析結果與前者略有不同,為EF-1α>18S rRNA>β-actin>GAPDH。geNorm的分析結果表明,在鯉皰疹病毒Ⅱ型感染異育銀鯽腎臟組織不同時間點18S rRNA和β-actin的表達穩定性一致(表3)。18S rRNA和β-actin的geNorm穩定值均為0.535(圖5)。由于18S rRNA的Ct值過小,容易在試驗中造成誤差[29],所以綜合4種軟件分析結果,結合geNorm穩定值以及4個內參基因的表達Ct值,最終確定β-actin最穩定。
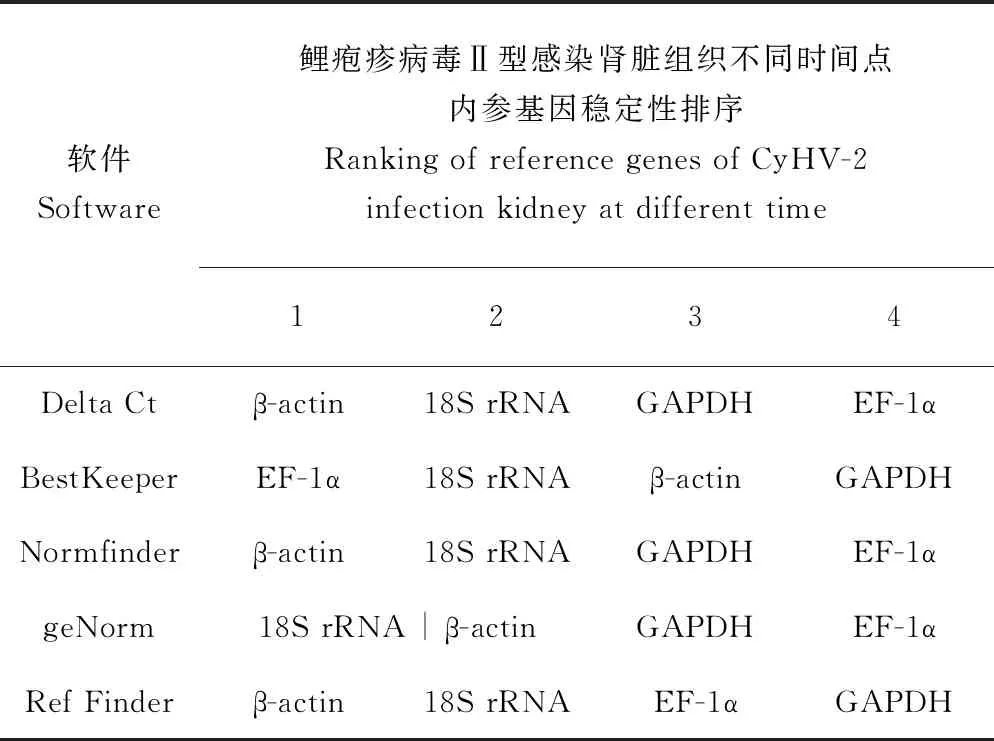
表3 鯉皰疹病毒Ⅱ型感染腎臟組織不同時間點內參基因穩定性排序
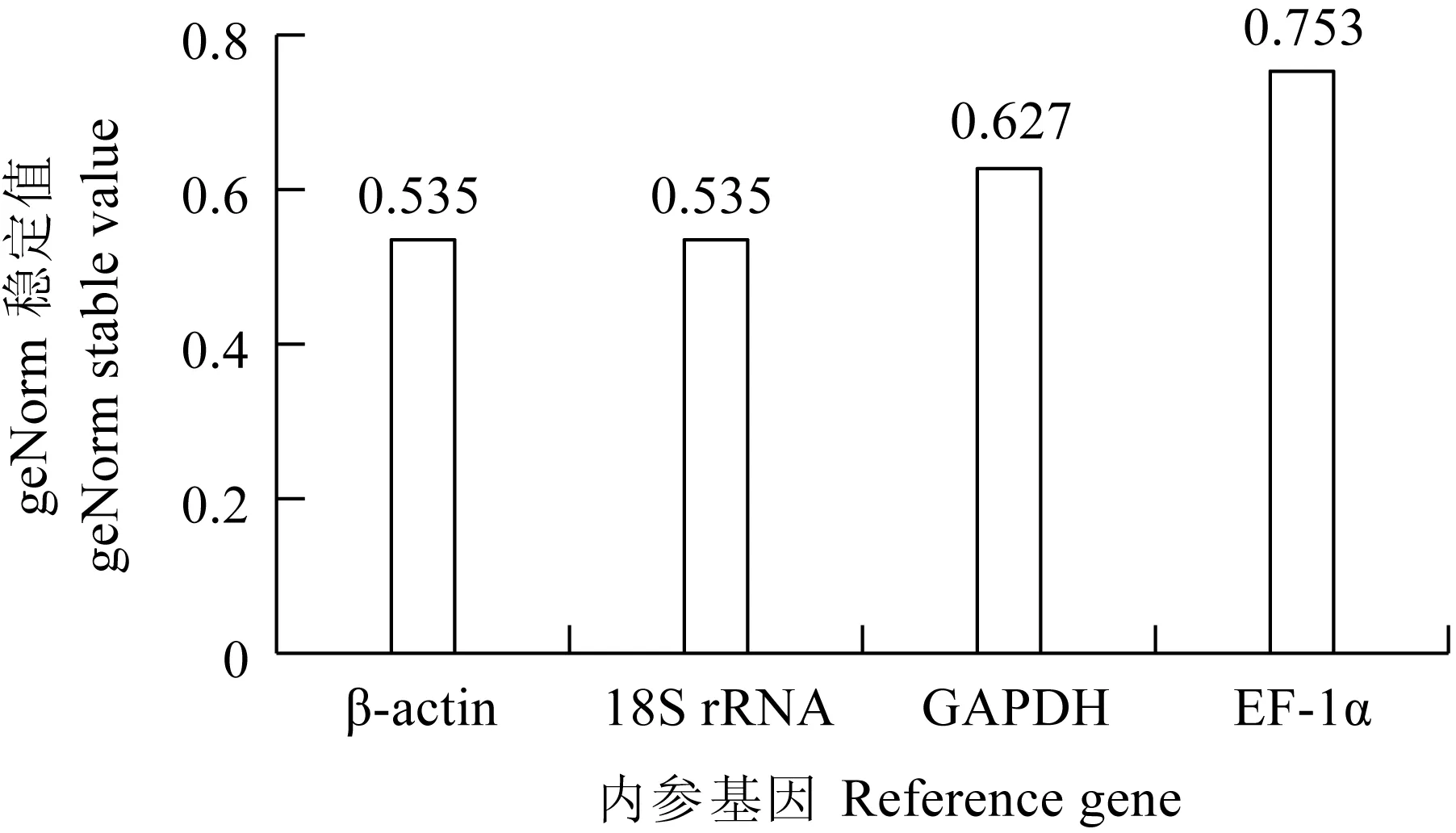
圖5 CyHV-2感染異育銀鯽腎臟組織不同時間點geNorm穩定值
2.4 鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織在不同時間點候選內參基因的穩定性評估
檢測鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織在不同時間點(0、6、12、24、48 h和120 h)候選內參基因的表達量Ct值分布見圖6。4個候選內參基因的Ct值為9.63~31.34,其中18S rRNA表達水平受感染時間影響差異較大,最大值和最小值相差21.71個循環。GAPDH的Ct值變化較小,最大值和最小值相差6.04個循環,但GAPDH的表達量都較EF-1α和β-actin的表達低。而EF-1α和β-actin的表達量均穩定在15~23。

圖6 CyHV-2感染異育銀鯽脾臟組織不同時間點候選內參基因Ct值分布
Delta Ct的分析結果與geNorm的分析結果幾乎一致,在鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織不同時間點EF-1α和β-actin穩定性較好。其中geNorm的穩定值結果顯示,EF-1α和β-actin的穩定性相差不大(圖7)。與Delta Ct 和geNorm的分析結果不同,BestKeeper與Normfinder的分析結果表明,在鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織中EF-1α和GAPDH的表達相對較穩定(表4)。綜合4種軟件分析結果,結合geNorm穩定值以及4個候選內參基因的表達Ct值,最終確定,在鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織不同時間點EF-1α相對比較穩定。
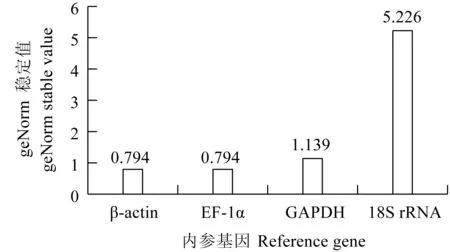
圖7 CyHV-2感染異育銀鯽脾臟組織不同時間點geNorm穩定值
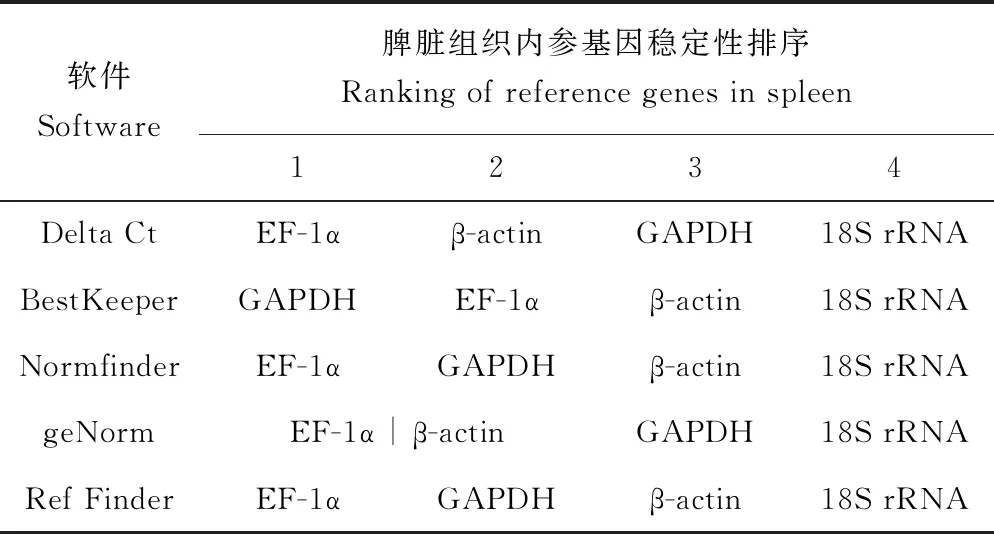
表4 鯉皰疹病毒Ⅱ型感染異育銀鯽脾臟組織不同時間點內參基因穩定性排序
2.5 鯉皰疹病毒Ⅱ型感染異育銀鯽尾鰭細胞在不同時間點候選內參基因的穩定性評估
檢測鯉皰疹病毒Ⅱ型感染尾鰭細胞不同時間點(0、1、2、3、4 d和5 d),4個候選內參基因的表達量Ct值分布見圖8。4個候選內參基因的Ct值為7.39~36.54,其中GAPDH表達水平受感染時間影響差異較大,最大值和最小值相差4.67個循環。18S rRNA的Ct值變化較小,最大值和最小值相差2.98個循環。β-actin和EF-1α在細胞感染不同時間點的表達量Ct值均相對穩定(圖9,表5)。
2.6 內參基因的驗證
研究表明,在鯉皰疹病毒Ⅱ型感染異育銀鯽腎臟組織過程中,隨著感染時間延長,病毒拷貝數逐漸升高,PIN1基因的mRNA表達量顯著下降[21]。為驗證測序結果的準確性,本次定量驗證中添加6 h的試驗組。經過SPSS 19.0分析,試驗結果和腎臟組織轉錄組cDNA的測序結果(TPM值)見圖10。PIN1基因以GAPDH為內參基因時,在鯉皰疹病毒Ⅱ型感染腎臟組織0 h和72 h時的表達趨勢與測序結果不符;PIN1基因以18S rRNA為內參基因,目的基因表達在0 h和72 h沒有顯著差異。相反,以β-actin和EF-1α為內參基因時,PIN1基因在鯉皰疹病毒Ⅱ型感染腎臟組織0 h和72 h時的表達趨勢與測序結果一致,且差異極顯著(P<0.01)。隨著感染時間的增加,PIN1基因以β-actin基因為內參分別在鯉皰疹病毒Ⅱ型感染腎臟組織0、6 h和72 h時相對表達量差異極顯著(P<0.01)。而PIN1基因以EF-1α基因為內參在鯉皰疹病毒Ⅱ型感染腎臟組織0 h和72 h時相對表達量差異極顯著(P<0.01)(圖11),但在PIN1基因以EF-1α為內參在鯉皰疹病毒Ⅱ型感染腎臟組織6 h和72 h時相對表達量無極顯著差異(P>0.01)。因此,在鯉皰疹病毒Ⅱ型感染異育銀鯽的腎臟組織中β-actin為其最佳的內參基因。
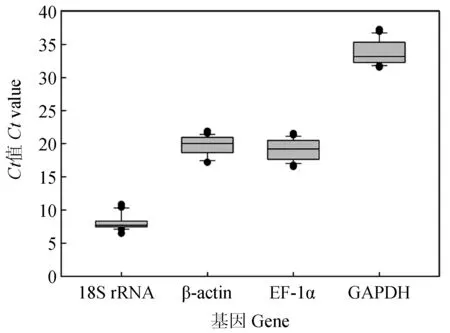
圖8 CyHV-2感染GiCF細胞不同時間點候選內參基因Ct值分布
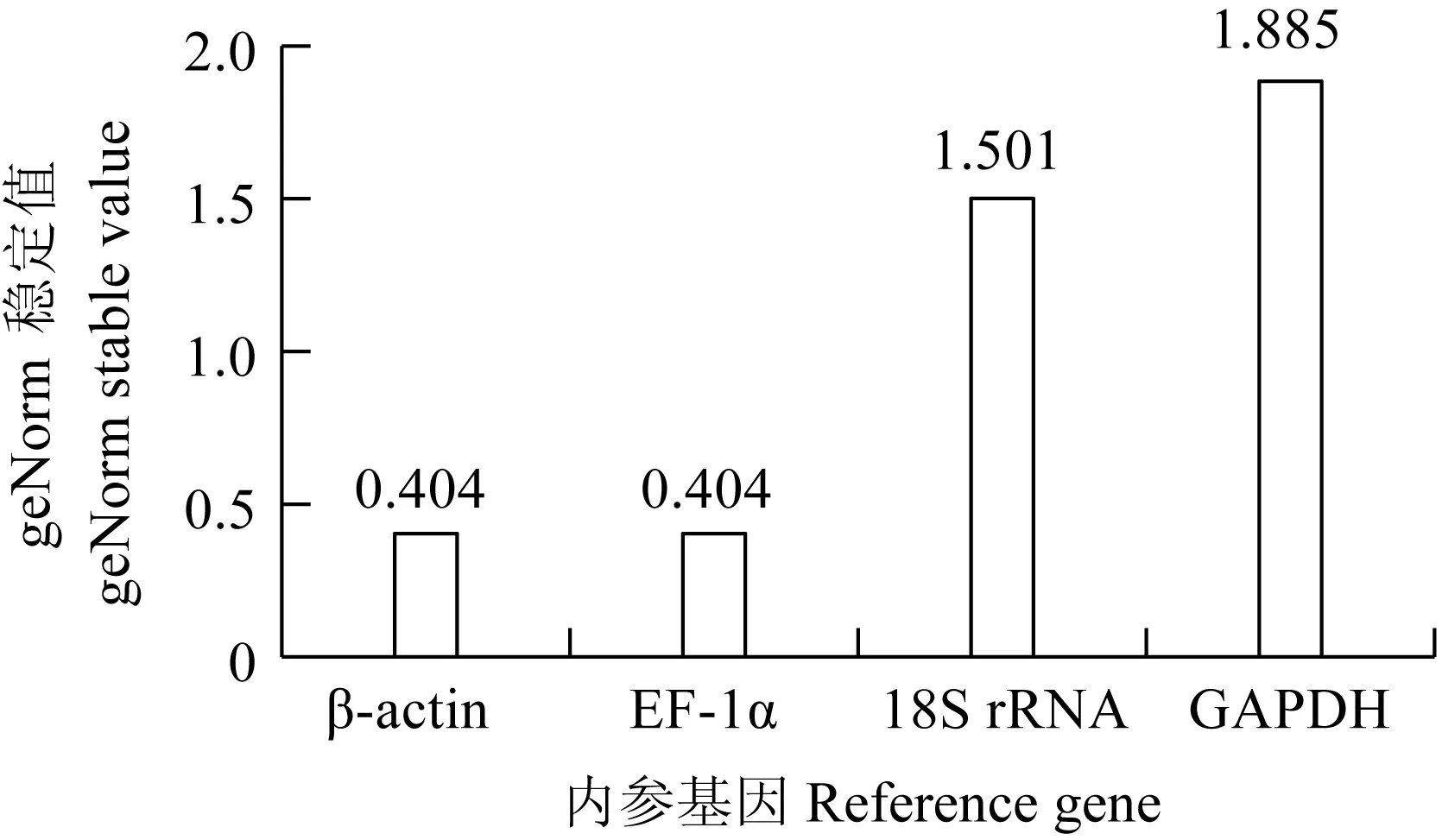
圖9 CyHV-2感染GiCF細胞不同時間點geNorm 穩定值

表5 鯉皰疹病毒Ⅱ型感染細胞不同時間點內參基因的穩定排名情況
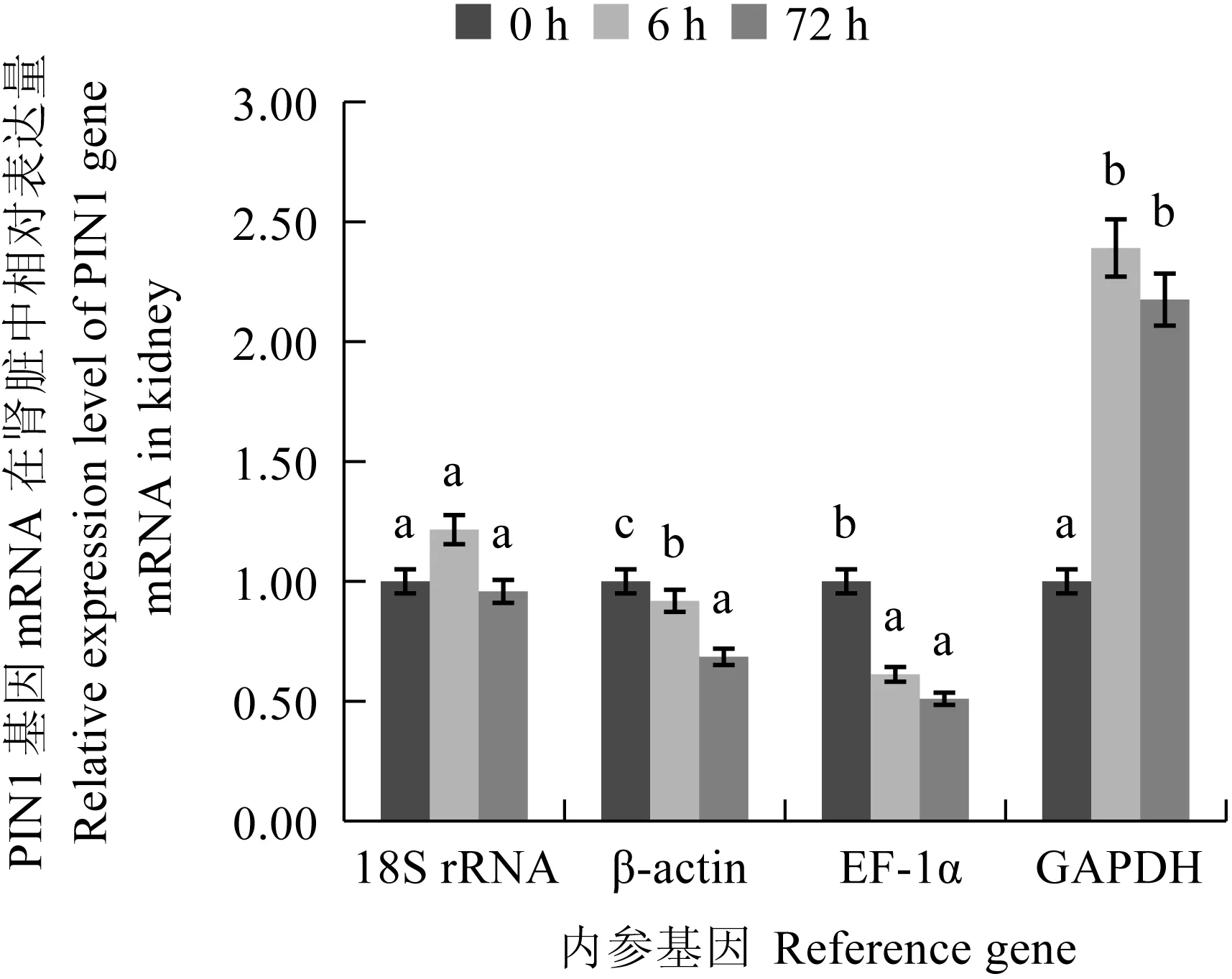
圖10 PIN1基因分別以4個候選內參基因為內參在腎臟中的表達量
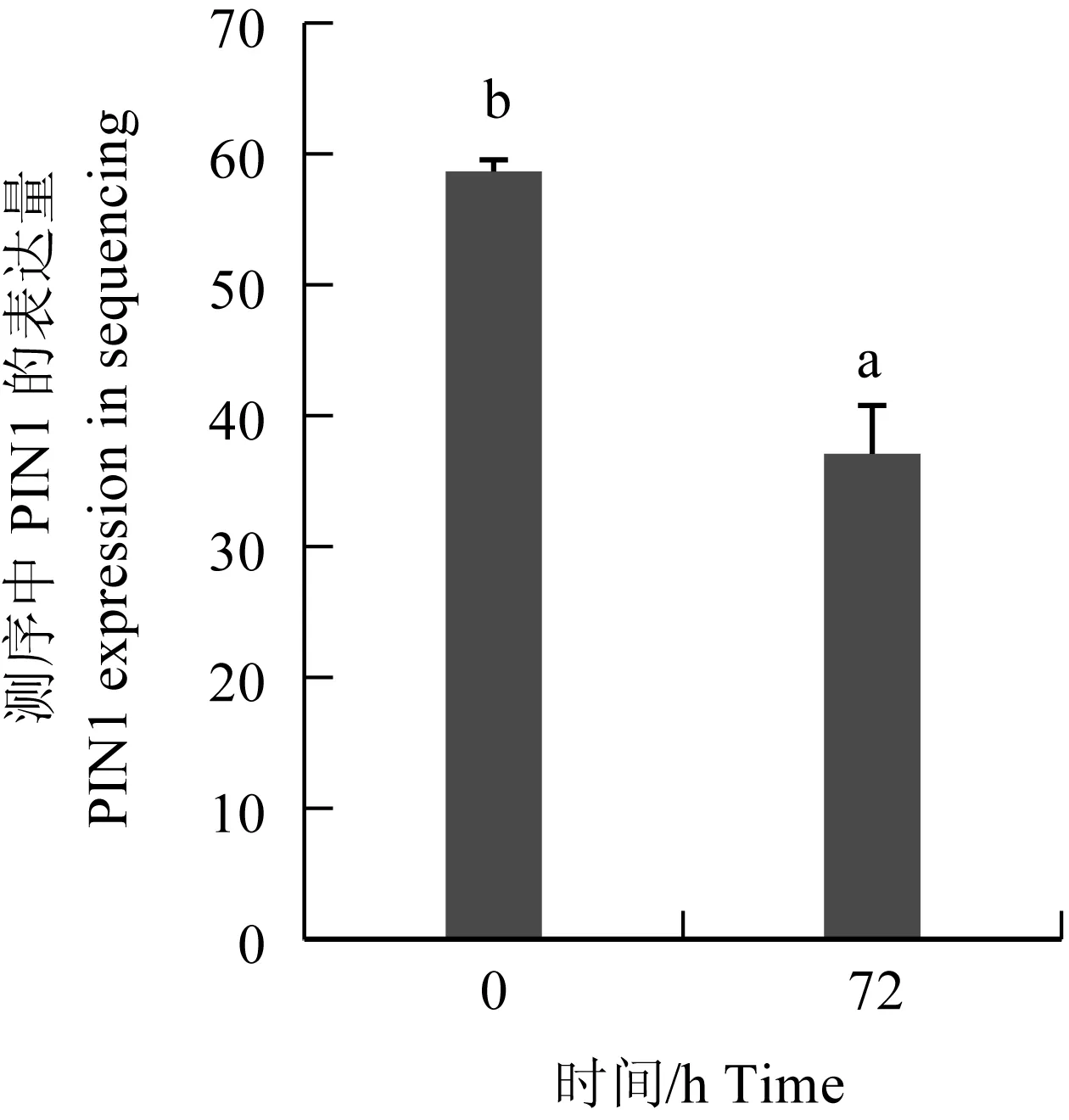
圖11 PIN1在轉錄組數據庫測序中的表達量
3 討 論
3.1 內參基因在不同組織中穩定性分析
為了使基因在qRT-PCR試驗中的相對表達結果更可靠,篩選最佳的內參(管家)基因是必不可少的。研究水產動物常用的管家基因很多,18S rRNA、GAPDH、EF-1α和β-actin是4個較為常用的內參基因。其中,甘油醛-3-磷酸脫氫酶(GAPDH)是一種參與糖酵解和糖異生等循環過程的關鍵酶[30];延伸因子(EF-1α)是參與蛋白質翻譯的重要因子[31];18S核糖體RNA(18S rRNA)是一類編碼核糖體小亞基RNA[32];β-肌動蛋白(β-actin)是構成細胞骨架的主要成分之一。然而不同物種、不同組織、不同發育時期以及不同處理條件,管家基因的穩定性并不是一成不變的,如在褐牙鲆胚胎不同發育時期18S rRNA是其最合適的內參基因[33];鱖魚的不同組織中GAPDH為其內參基因[34]。因此只有篩選出合適的內參基因才能保證qRT-PCR試驗結果的準確性。
本試驗選取健康異育銀鯽的8種不同組織為研究對象,選擇EF-1α、GAPDH、18S rRNA和β-actin 4個候選管家基因,結合4種內參基因分析軟件,geNorm穩定值以及4個內參基因的表達量Ct值,最終分析結果表明,β-actin和EF-1α基因為健康異育銀鯽的不同組織中最穩定的基因。董捷[26]研究草魚(Ctenopharyngodonidella)不同組織基因表達豐度時,發現18S rRNA和β-actin是最穩定的內參基因;Olsvik等[35-36]比較了大西洋鮭(Salmosalar)不同組織中β-actin、RPS20和EF-1α的表達穩定性,發現EF-1α基因的表達最穩定;茅華華等[37]發現斑鱧(Channamaculata)在成魚不同組織中β-actin基因最穩定。健康異育銀鯽的不同組織內參基因β-actin和EF-1α的穩定性與上述研究都有相似之處。這可能因為不同的物種中管家基因的轉錄水平不同,但是其在特定組織的穩定性可能存在一致性。在脊椎動物魚類和無脊椎動物貝類中,雖然不同的物種中β-actin和EF-1α基因表達量不同,如草魚[26]、鱸鯉(Percocyprispingi)[38]、鱖魚[4]、建鯉 (Cyprinuscarpiovar.jian)[39]、皺紋盤鮑(Haliotisdiscushannai)[40]、合浦珠母貝(Pinctadafucata)[41]和三角帆蚌 (Hypriopsiscumingii)[42],但是其仍然具有良好的穩定性,這與本試驗中的結果一致,表明β-actin和EF-1α基因同樣可以作為魚類與海洋無脊椎動物的內參基因。
3.2 內參基因在病毒刺激下穩定性分析
生物體通常都有自我保護的意識,在病毒刺激下會產生相應的免疫應答反應,篩選出穩定的內參基因,為研究病毒刺激下免疫基因的表達提供理論基礎。
Jorgensen等[43]用傳染性鮭魚貧血病毒(Infectioussalmonanemiavirus, ISAV)感染大西洋鮭和腎細胞發現感染不同時間點組織水平18S rRNA和EF-1α最穩定,而細胞水平僅有18S rRNA最穩定。與上述試驗結果類似的是本試驗用鯉皰疹病毒Ⅱ型感染異育銀鯽的脾臟組織不同時間點,EF-1α為其穩定的內參基因。不同的是本試驗選用鯉皰疹病毒Ⅱ型感染異育銀鯽的腎臟和尾鰭細胞不同時間點發現β-actin為其最穩定的基因。這與Julin等[44]用傳染性胰腺壞死病毒(Infectiouspancrieaticnecrosisvirus, IPNV)感染大西洋鮭不同時間點,發現EF-1α和β-actin的穩定性最高的結論相似。有研究表明,異育銀鯽的腎臟是鯉皰疹病毒Ⅱ型的主要復制點,脾臟是淋巴細胞移居和接受抗原刺激后產生免疫應答的主要場所[9]。所以在鯉皰疹病毒Ⅱ型感染異育銀鯽的腎臟、脾臟不同時間點內參基因的穩定性不同,這可能與病毒刺激條件下,基因在組織間的選擇性表達有關。此外本試驗由于18S rRNA的表達過于豐富,可能會比目的基因的表達還要高,容易在試驗過程中和數據分析時造成誤差,且有研究表明,18S rRNA的轉錄容易受到生物因素和藥物因素影響[29]。EF-1α是真核生物的延伸因子,其在組織中穩定性已經被大量的學者研究證明,如大鯢(Andriasdavidianus)在細菌的感染下組織的內參基因的穩定性評價顯示,EF-1α是穩定的內參基因[45]。本試驗的結果分析也表明,EF-1α可作為病毒刺激下穩定的內參基因。β-actin是一種微絲的結構蛋白,在抵御某些病毒和病原微生物的入侵時,β-actin的分泌穩定,可以用作為內參基因對其他基因的標準化處理[46]。因此,在病毒刺激下,β-actin較適合作為異育銀鯽腎臟組織和細胞水平qRT-PCR的內參基因。
PIN1是一種高度保守的特異的多肽脯氨酰基順反異構酶具有促進細胞增殖的功能[47]。數據庫測序結果顯示,PIN1基因表達量呈下調趨勢,本試驗的qRT-PCR結果也顯示,以β-actin為內參時PIN1基因表達量符合測序結果。Lu等[22]在異育銀鯽腎臟組織的cDNA文庫測序過程中發現,PIN1基因表達量在感染魚體過程中呈現組織水平的下調趨勢與本試驗結果相符合,這可能在病毒的刺激下PIN1基因的轉錄水平受到抑制從而導致細胞增殖的功能受阻。
4 結 論
本試驗通過qRT-PCR技術,篩選出異育銀鯽組織水平和細胞水平中均較穩定的內參基因。試驗結果表明,在健康異育銀鯽的不同組織中β-actin和EF-1α均可作為其內參基因。在鯉皰疹病毒Ⅱ型感染腎臟以及細胞系不同時間點,β-actin為其內參基因;脾臟組織中,EF-1α為其內參基因。通過在異育銀鯽腎臟組織不同感染時間點,PIN1基因的表達分析也進一步驗證了其內參基因的適用性,本研究將為探究異育銀鯽基因的qRT-PCR提供理論依據,為深入查明異育銀鯽體內及體外宿主的基因表達與病毒相互作用奠定基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
