青海省櫻桃葉斑病病原菌的分離與鑒定
2020-06-08 09:41:12劉俏寧楠楠馬永強
植物保護 2020年2期
劉俏 寧楠楠 馬永強


摘要 為明確引起青海省櫻桃葉斑病病原菌的種類,本研究從西寧市城北區(qū)、海東市樂都區(qū)和貴德縣的櫻桃上采集有葉斑癥狀的葉片,采用組織分離法分離出病原菌,通過觀察病原菌的形態(tài)特征,結(jié)合rDNA-ITS、EF-1α和Alt a 1基因序列分析對病原菌的種類進行了鑒定,并用柯赫氏法則進行驗證。結(jié)果表明,共獲得279株病原菌菌株,分屬鏈格孢Alternaria alternata、細極鏈格孢A.tenuissima和刺盤孢屬Colletotrichum spp.,分離頻率分別為84.95%、5.02%和10.03%。采用柯赫氏法則進行離體葉片致病性測定,接種葉片100%發(fā)病,說明引起青海省櫻桃葉斑病的病原菌為鏈格孢、細極鏈格孢和刺盤孢屬真菌,且鏈格孢為主要病原菌。該研究結(jié)果可為青海省櫻桃葉斑病的綜合防治提供理論依據(jù)。
關(guān)鍵詞 櫻桃; 葉斑病; 鏈格孢屬; 刺盤孢屬; Alt a 1基因; 系統(tǒng)發(fā)育
中圖分類號: S 436.62 ?文獻標識碼: A ?DOI: 10.16688/j.zwbh.2019567
Abstract In order to confirm the pathogen of cherry leaf spot in Qinghai province, the typical diseased cherry leaves were collected from Chengbei district of Xining city, Ledu district and Guide county of Haidong city. The pathogen was isolated by tissue separation method and identified by morphological observation and sequence analysis of rDNA-ITS, EF-1α and Alt a 1 genes. The pathogenicity of the strains were tested by Kochs postulates. The results showed that 279 strains were obtained, belonging to Alternaria alternata, A.tenuissima and Colletotrichum spp., with the isolation frequencies of 84.95%, 5.02% and 10.03%, respectively. Kochs postulates test demonstrated that 100% incidence was found on the inoculated leaves, indicating that the pathogens causing cherry leaf spot disease in Qinghai province are A.alternata, A.tenuissima and Colletotrichum spp., and A. alternaria is the main pathogen. The results provide a theoretical basis for the comprehensive prevention and control of cherry leaf spot in Qinghai province.
Key words cherry; leaf spot; Alternaria spp.; Colletotrichum spp.; Alt a 1 gene; phylogeny
櫻桃Prunus avium 又名迎慶果、含桃,薔薇科李屬落葉喬木果樹。因果實色澤鮮艷,營養(yǎng)豐富,酸甜可口,而深受人們的喜愛,有“百果第一枝,早春第一果”的美譽[1]。世界上櫻桃主要分布在美國、加拿大、歐洲等北半球國家,在國內(nèi)作為果樹栽培的櫻桃有中國櫻桃、甜櫻桃、酸櫻桃和毛櫻桃,主要分布在山東、陜西、四川等地[2]。由于其果實經(jīng)濟價值高,近年來,在我國各省市得到大面積推廣栽培,我國櫻桃的栽培面積日益增加,目前已居世界首位[3]。隨著栽培面積的不斷擴大,發(fā)生在櫻桃上的病害也呈現(xiàn)出明顯加重的趨勢。目前,國內(nèi)外危害櫻桃產(chǎn)業(yè)的病害主要有櫻桃褐斑病[4]、流膠病[5]、根癌病[6]、灰霉病[7]、褐腐病[8]和葉斑病[9]等。其中櫻桃葉斑病是世界范圍內(nèi)危害櫻桃產(chǎn)業(yè)的真菌性病害,在歐美國家發(fā)生極其嚴重,給當?shù)毓r(nóng)造成了巨大經(jīng)濟損失,嚴重阻礙了櫻桃產(chǎn)業(yè)的健康發(fā)展[10-15]。目前,引起櫻桃葉斑病的病原菌有Blumeriella jaapii[16]、Calonectria ilicicola[17]、Pseudocercospora pruni-persicicola[18]、Cylindrosporium padi[19]和Phoma herbarum[20]。此外, Thomidis 和 Tsipouridis[9]于2006年報道了希臘櫻桃葉斑病的病原菌為鏈格孢屬真菌中的小孢子種鏈格孢Alternaria alternata。我國學(xué)者趙遠征等[21]于2013年在大櫻桃果實上分離出鏈格孢,并證實了該種病原菌人工有傷接種可侵染葉片。但在自然條件下由鏈格孢屬真菌和刺盤孢屬真菌引起櫻桃葉斑病的研究在國內(nèi)尚未見報道。
本研究組在2017年-2019年對青海省櫻桃病害進行系統(tǒng)調(diào)查時發(fā)現(xiàn)兩種新的葉部病害普遍發(fā)生。在發(fā)病最為嚴重的7、8月份,病株率高達100%。發(fā)病葉片形成圓形或近圓形的褐色或紫紅色病斑,大小不等。發(fā)病嚴重時,病斑相互連接,擴展至整個葉面,導(dǎo)致光合作用嚴重受阻,樹勢衰弱,第二年果實產(chǎn)量降低。為明確青海省櫻桃葉斑病的病原菌,本研究采用組織分離技術(shù)對病原菌進行分離,柯赫氏法則進行致病性測定,結(jié)合形態(tài)學(xué)和分子生物學(xué)手段對病原菌進行鑒定,以期為青海省櫻桃葉斑病病原菌的科學(xué)診斷和病害的綜合防治提供理論基礎(chǔ)和技術(shù)支持。
1 材料與方法
1.1 病葉的采集
櫻桃病葉于2017年-2019年采自青海省西寧市城北區(qū)、海東市樂都區(qū)和貴德縣。葉斑病A共采集90片病葉,其中西寧市城北區(qū)3片,海東市樂都區(qū)76片,貴德縣11片;葉斑病B共采集44片病葉,其中海東市樂都區(qū)40片,貴德縣4片。采集發(fā)病癥狀典型的新鮮櫻桃病葉放入采樣袋內(nèi),及時帶回實驗室分離。
1.2 病原菌的分離與純化
采用組織分離法[22]對病原菌進行分離,用 PDA培養(yǎng)基培養(yǎng)3~5 d后,在無菌條件下用打孔器在菌落邊緣切取直徑為5 mm的菌餅接種在PDA平板上,進行純化和轉(zhuǎn)接培養(yǎng)。純化2~3次后,將菌株接種在試管斜面PDA培養(yǎng)基上,置于4℃保存?zhèn)溆谩?/p>
1.3 病原菌的形態(tài)學(xué)鑒定
用滅菌打孔器在菌落邊緣切取直徑為5 mm的菌餅接種在PDA和PCA培養(yǎng)基的中央,每天觀察并記錄菌落的形狀、大小、顏色以及菌絲的生長狀況等形態(tài)特征。為了更清晰地觀察病原菌的形態(tài)特征,在PCA培養(yǎng)基上斜插無菌蓋玻片,待菌落生長7 d后,取出PCA培養(yǎng)基中的插片,置于載玻片上,滴一滴清水,制成臨時玻片,于顯微鏡下觀察記錄病原菌的菌絲形態(tài),分生孢子梗和分生孢子的形狀、大小、顏色、著生方式以及有無隔膜等特征,并參照張?zhí)煊頪23]對鏈格孢屬真菌種級形態(tài)的描述及Sutton[24]對刺盤孢屬的分類方法鑒定病原菌的種類。
1.4 病原菌的分子生物學(xué)鑒定
采用CTAB法[25]提取病原菌的總DNA。分別以通用引物ITS1和ITS4[26]、EF1-728F和EF1-986R[27]、Alt-for和Alt-rev[28]進行PCR擴增,所用引物見表1,引物均由生工生物工程(上海)股份有限公司合成。PCR反應(yīng)體系均為25 μL:10×PCR buffer 2.5 μL(含Mg2+),模板DNA 0.5 μL,dNTPs 1.0 μL,引物各 0.5 μL(10 μmol/L),5 U/μL Taq酶0.2 μL,用ddH2O補齊。94℃ 預(yù)變性4 min;94℃變性45 s,55℃退火45 s,72℃延伸 1 min,30個循環(huán); 72℃延伸10 min,4℃保溫。擴增完成后,在1%瓊脂糖凝膠中通過150 V和100 mA電泳檢測5 μL擴增產(chǎn)物20 min,用SanPrep柱DNA J凝膠回收試劑盒(SK8131,上海生工)回收PCR擴增產(chǎn)物,并送至上海生工進行雙向測序。測序結(jié)果與GenBank數(shù)據(jù)庫中的核酸序列進行BLAST同源性比較分析。利用ClustalX 1.8和MEGA 5.1進行序列間的比對分析,將3條基因序列按照rDNA-ITS、EF-1α和Alt a 1的順序進行多基因序列整合,以鄰接法(NJ)構(gòu)建菌株的系統(tǒng)發(fā)育樹,并用自展法(bootstrap method)進行檢驗,重復(fù)1 000次,分析該菌與同屬菌株間的親緣關(guān)系[1]。
1.5 病原菌的致病性測定
采用離體葉片接種方式進行病原菌的致病性測定。將純化后的病原菌菌株接種在PDA平板上,25℃恒溫,黑暗倒置培養(yǎng)6 d。用滅菌打孔器在菌落邊緣打出直徑為5 mm的菌餅,接種在健康櫻桃葉片上。采用有傷(針刺)和無傷兩種接種方式,先用自來水沖洗掉葉片表面的雜質(zhì),再用75%乙醇消毒30 s,滅菌水沖洗3次,用滅菌濾紙將葉片表面的水分吸干后,進行有傷和無傷接種,每個葉片接種6個菌餅,重復(fù)3次,用空白PDA培養(yǎng)基作為對照。置于25℃的恒溫培養(yǎng)箱中,黑暗培養(yǎng)7 d后調(diào)查發(fā)病率,并對發(fā)病葉片再次進行病菌分離和鑒定。具體方法是用滅菌剪刀在發(fā)病葉片病健交界處剪取0.5 cm×0.5 cm的病組織小塊,于超凈工作臺上用75%乙醇消毒3 min,無菌水沖洗3次,放在滅菌濾紙上晾干后,接種在PDA平板上,置于25℃恒溫培養(yǎng)箱中培養(yǎng),待菌落長出后進行形態(tài)學(xué)和分子生物學(xué)鑒定。發(fā)病率=發(fā)病葉片數(shù)/接種葉片總數(shù)×100%。
2 結(jié)果與分析
2.1 發(fā)病癥狀
葉斑病A發(fā)病初期在葉片上形成大小不一的褐色斑點,后病斑逐漸擴大為圓形或近圓形褐色斑,直徑為2.23~11.67 mm,病斑處干枯、皺縮,略微凹陷,且產(chǎn)生同心輪紋,中間為黃褐色或淺褐色,病健交界處深褐色且隨著病斑的不斷擴大產(chǎn)生離層,常由此脫落形成穿孔。病斑背面直徑為2.30~11.35 mm,顏色較正面淺,為黃褐色,同心輪紋不明顯。發(fā)病嚴重時,病斑相互連接形成不規(guī)則的病斑塊,且擴展范圍不受葉脈限制。發(fā)病嚴重的葉片失水嚴重,萎蔫、皺縮、柔韌性降低,尤其是病斑處因干枯而出現(xiàn)裂痕(圖1)。葉斑病B發(fā)病葉片形成圓形或近圓形的紫褐色病斑,直徑為1.47~4.60 mm,病斑內(nèi)部很小部分為黃褐色,向外為紫色環(huán),且最外層有紫紅色光暈。病健交界處不產(chǎn)生離層,不形成穿孔。病斑背面為紫紅色,直徑為1.13~4.52 mm。發(fā)病嚴重時,病斑相互連接,擴展整個葉面,使葉片干枯、皺縮(圖2)。
2.2 病原菌的分離與純化
對兩種葉斑病的病原菌進行分離和純化,共獲得279株分離物,其中從葉斑病A的病葉中分離出153株,包括西寧3株,海東135株,貴德15株。通過培養(yǎng)形態(tài)和鏡檢分別將西寧3株分類命名為CBX001;海東135株分類命名為HS002(11株)、LD002(3株)、GM001(5株)、JX001(5株)、LJH001(22株)、LJH002(2株)、YR001(2株)、TGW001(78株)、TGW003(7株);貴德15株分類命名為GDX001(4株)、GDX003(2株)、GDD001(5株)和GDH001(4株)。從葉斑病B的病葉中共獲得126株病原菌株,包括海東113株,分別命名為TGT001(82株)、TGT002(28株)和TGT003(3株);貴德13株,均命名為GDY001。
2.3 病原菌的形態(tài)學(xué)鑒定
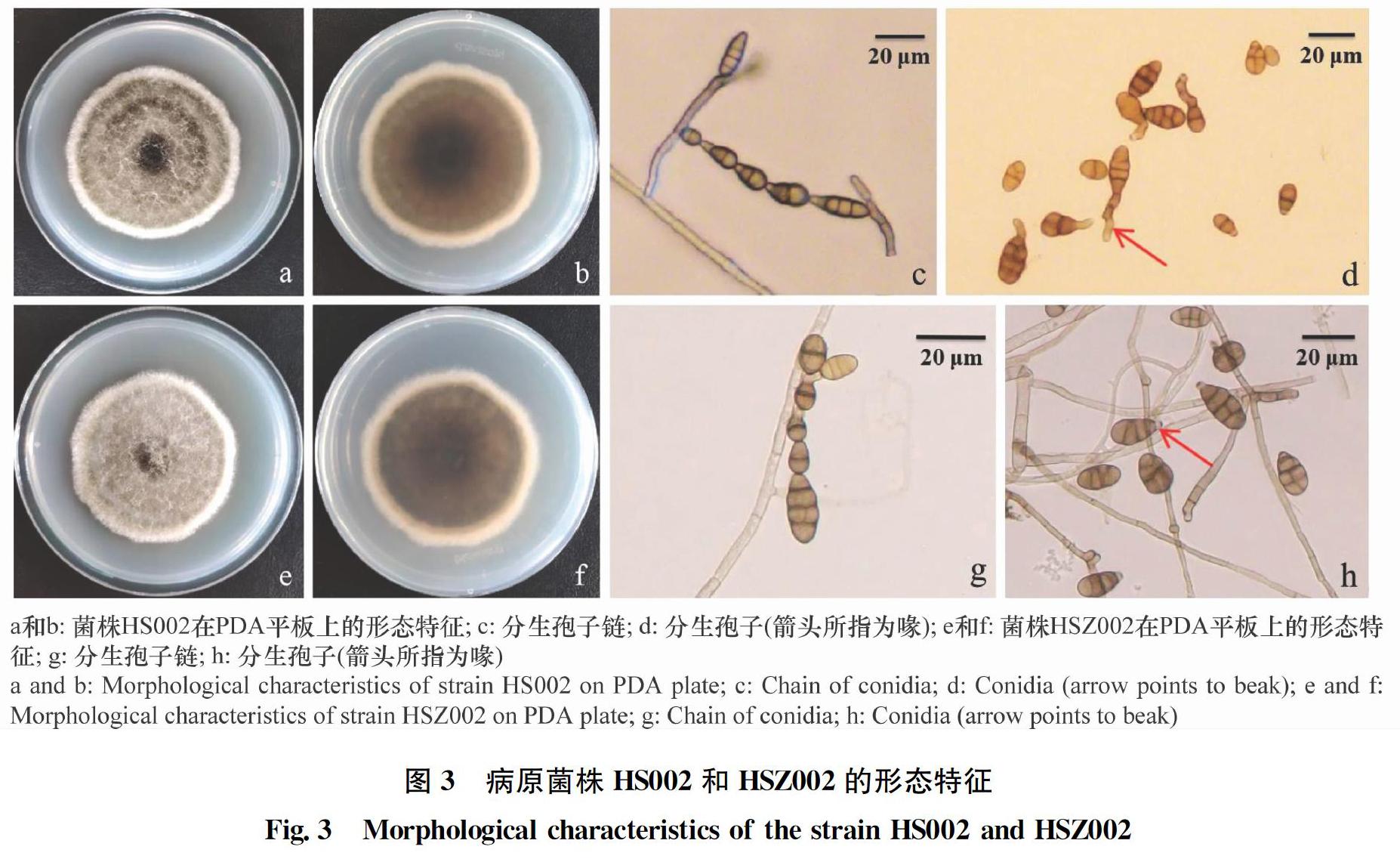
病原菌株CBX001、HS002、LD002、GM001、JX001、LJH001、YR001、TGW001、GDX001、GDD001、GDH001、TGT001和GDY001在PDA平板上菌落為圓形,呈灰黑色或墨綠色絨狀,邊緣整齊,背面深橄欖綠或黑色。培養(yǎng)初期,菌絲呈放射狀生長,為白色,后逐漸變?yōu)榛野咨⒒疑蚝谏嗑砬豢棾山q毛狀。氣生菌絲發(fā)達,致密,生長迅速,7 d后菌落直徑達9 cm(圖3 a和b)。在顯微鏡下,病原菌的菌絲為無色,寬3~5.5 μm,有隔膜。分生孢子梗淺褐色或棕褐色,著生于菌絲末端或側(cè)面,直立或屈膝狀彎曲,有分隔,大小為(42.3~96.5)μm×(3.2~4.7)μm。分生孢子黃褐色或褐色,長橢圓形、卵形、棍棒狀或倒梨形,有1~5個橫隔膜,0~3個豎或斜隔膜,分隔處大多具有縊縮現(xiàn)象,大小為(9.6~35.5)μm×(6.5~15.5)μm。分生孢子著生于分生孢子梗頂端或側(cè)面,呈鏈狀生長,每條鏈上有2~6個孢子。分生孢子鏈具有二次分支現(xiàn)象,形似樹狀。分生孢子多數(shù)具有喙,短喙為淺褐色柱狀或錐狀,長喙棕褐色且具有分隔,大小為(3.2~21.3)μm×(3.4~5.1)μm(圖3 c和d)。
菌株LJH002、TGW003、GDX003和TGT003在PDA上菌落為圓形,呈灰褐色或墨綠色絨狀,邊緣整齊,背面深橄欖綠或黑色。菌絲粗壯,菌落粗糙,培養(yǎng)初期菌絲呈放射狀生長,為白色,后逐漸變?yōu)榛疑⒛G色或黑色。氣生菌絲發(fā)達,致密,生長迅速,9 d后菌落直徑達9 cm(圖4a和b)。分生孢子梗淺褐色,多數(shù)單生于菌絲側(cè)面,偶有分支,直立或略彎曲,有分隔,大小為(26.5~83.9)μm×(3.6~5.2)μm。分生孢子淺褐色或褐色,橢圓形、卵形、棍棒狀,少數(shù)倒梨形,有1~7個橫隔膜,0~5個豎或斜隔膜,分隔處有明顯縊縮現(xiàn)象,成熟的孢子常有1~4個主橫隔膜因較粗而顏色加深,為黑褐色,大小為(15.7~46.8)μm×(7.2~15.7)μm。分生孢子呈鏈狀生長,不分支或少分支,每條鏈上有6~10個孢子。喙或假喙柱狀或錐狀,淺褐色,有隔膜,基部略微膨大,大小為(3.3~3.4)μm×(3.4~4.9)μm(圖4c和d)。
菌株TGT002在PDA平板上菌落圓形,呈粉紅色、灰白色或灰褐色毛絨狀,邊緣整齊,背面粉紅色、橘黃色或紫褐色。培養(yǎng)初期,菌絲呈白色或粉紅色,后逐漸變?yōu)榛野咨蚧疑揖涑霈F(xiàn)同心輪紋。氣生菌絲發(fā)達、致密,生長迅速,6 d后菌落直徑達4.8 cm(圖5 a和b)。病原菌菌絲直徑為1.92~5.79 μm,透明且有隔膜。分生孢子梗無色,著生于菌絲側(cè)面,單生且有隔膜,具有分枝。分生孢子梭形、圓柱狀,光滑透明,無隔膜,大小為(10.72~15.82)μm×(3.75~6.86)μm(圖5 c和d)。
2.4 病原菌的分子生物學(xué)鑒定
提取分類命名后18株病原菌的總DNA,經(jīng)PCR擴增、測序后將結(jié)果提交至NCBI網(wǎng)站進行BLAST同源性比較分析。結(jié)果表明,基于rDNA-ITS基因序列所測長度為545~583 bp,EF-1α基因所測序列長度為253~285 bp,Alt a 1基因所測長度為480~532 bp。將病原菌的rDNA-ITS、EF-1α和Alt a 1基因序列合并構(gòu)建系統(tǒng)發(fā)育樹,結(jié)果表明, 13株菌株以100%支持率與A.alternata聚為同一支,4株菌株與A.tenuissima以100%支持率聚為一支(圖6)。病原菌TGT002構(gòu)建的系統(tǒng)發(fā)育樹結(jié)果顯示,該菌與刺盤孢屬真菌中的多個種聚為一支(圖7)。
2.5 279株病原菌的鑒定結(jié)果和分布
結(jié)合病原菌的形態(tài)特征和系統(tǒng)發(fā)育分析結(jié)果,從葉斑病A病葉中分離出的153株病原菌被鑒定為鏈格孢屬中的兩個小孢子種:鏈格孢142株和細極鏈格孢11株,分離頻率分別為92.8%和7.2%。西寧、海東和貴德三個地區(qū)的不同采集地點均分離出了鏈格孢菌,其中西寧3株、海東126株、貴德13株;而只有在海東和貴德分離出了細極鏈格孢,分別為9株和2株。從葉斑病B病葉中分離出的126株病原菌中,95株被鑒定為鏈格孢,3株被鑒定為細極鏈格孢,28株被鑒定為刺盤孢屬,分離頻率分別為75.4%、2.4%和22.2%。海東和貴德兩個采集地點均分離出了鏈格孢,其中海東82株、貴德13株; 而3株細極鏈格孢和28株Colletotrichum spp.均從海東櫻桃病葉上分離得到。
2.6 病原菌的致病性測定
分別從三類菌株中各選取1株代表性菌株HS002(鏈格孢,樂都)、GDX003(細極鏈格孢,貴德)和TGT002(刺盤孢屬,樂都)進行致病性測定,結(jié)果表明,3株菌株接種的葉片均在接種后第3 天開始發(fā)病,且有傷和無傷接種均可發(fā)病,發(fā)病率為100%。菌株HS002和GDX003接種的葉片發(fā)病初期病斑呈暗褐色,后病斑逐漸擴大為圓形或近圓形褐色斑,病斑處出現(xiàn)同心輪紋,與田間自然發(fā)病癥狀相同;菌株TGT002接種的葉片病斑呈褐色,大小不等,后病斑逐漸擴大為圓形紅褐色或紫褐色斑,病斑處干枯且出現(xiàn)裂痕,而對照組葉片未發(fā)病(圖8)。從菌株HS002、GDX003和TGT002接種發(fā)病葉片上再次進行病菌分離,分別將病原菌命名為HSZ002、GDX003-HJT和TGT002-HZ。經(jīng)鏡檢后其形態(tài)(圖3e~h、圖4e~h和圖5e~h)與上述供試菌株形態(tài)一致,且HSZ002和GDX003-HJT在系統(tǒng)進化樹上與供試菌株聚為一支(圖6)。
3 討論
本研究結(jié)合形態(tài)學(xué)和分子生物學(xué)手段,最終將青海省櫻桃葉斑病的病原菌鑒定為鏈格孢、細極鏈格孢和刺盤孢屬,分離頻率分別為84.95%、5.02%和10.03%,且鏈格孢為引起青海省櫻桃葉斑病的主要病原菌。這是國內(nèi)首次發(fā)現(xiàn)在自然條件下鏈格孢屬真菌和刺盤孢屬真菌能夠引起櫻桃葉斑病。
世界上已報道的鏈格孢屬真菌有500多種,且95%寄生在不同植物上,具有寄主范圍廣,傳播途徑多,侵染能力強和危害程度大等特點[29]。該類菌種引發(fā)的病害在經(jīng)濟作物、糧食作物和果蔬生產(chǎn)中均有發(fā)生[30],嚴重阻礙了我國農(nóng)業(yè)生產(chǎn)的發(fā)展。
與其他真菌相比,鏈格孢屬真菌具有明顯的特征,易于區(qū)分,但種級間的特征存在很大變異,特別是屬內(nèi)的小孢子種極易受培養(yǎng)條件的影響而產(chǎn)生趨同現(xiàn)象,使鏈格孢屬真菌難以鑒定到種[31]。傳統(tǒng)的鏈格孢屬真菌分類方法以形態(tài)學(xué)為主,系統(tǒng)發(fā)育為輔,這種方法在一定程度上滿足了一些菌種的分類要求,但易受人為因素和環(huán)境條件的干擾,使得菌種之間的進化距離和親緣關(guān)系不能被準確反映,菌種的鑒定結(jié)果也存在一定的偏差[32]。近年來,隨著分子生物學(xué)技術(shù),尤其是PCR技術(shù)的廣泛應(yīng)用,使得人們從分子水平上認識到菌種的遺傳和進化情況,更能準確地進行菌種鑒定,基于此技術(shù),一些研究者結(jié)合形態(tài)特征對鏈格孢屬種級分類鑒定及其系統(tǒng)發(fā)育做出了廣泛的研究:宋順華等[33]、Hong等[34]和趙金梅等[35]利用rDNA-ITS序列對病原菌進行了分類鑒定,趙艷琴等[36]利用rDNA-ITS和EF-1α基因序列鑒定了病原菌的種類;趙圓等[37]和岳海梅等[38]利用endo PG序列,曲文文等[31]利用ITS序列并結(jié)合隨機片段OPA2-1和OPA1-3基因序列對鏈格孢屬種間的進化情況及其親緣關(guān)系進行了系統(tǒng)發(fā)育分析。
本研究利用rDNA-ITS、EF-1α和Alt a 1三個基因序列對致病菌株的系統(tǒng)發(fā)育進行了研究,這些序列在一定程度上反映了菌種的進化程度和親緣關(guān)系的遠近,但rDNA-ITS和EF-1α基因序列卻很難區(qū)分鏈格孢屬真菌中形態(tài)相近的小孢子種,由此推測rDNA-ITS和EF-1α基因在鏈格孢屬真菌的遺傳進化中存在一定的變異性,這與張榮等[39]和岳海梅等[40]的研究結(jié)果相似。特異性基因位點Alt a 1基因的保守性較其他已知基因位點高,且在鏈格孢屬不同種間遺傳信息差異大,可以將病原菌準確鑒定到種,Hong等[28]也對此進行過詳細的研究并得到相同的結(jié)果。本研究還單獨利用rDNA-ITS序列對刺盤孢屬進行系統(tǒng)發(fā)育分析,但該序列不能將病原菌準確鑒定到種。正如符丹丹[41]在對中國蘋果炭疽病病原菌遺傳多樣性研究中所發(fā)現(xiàn)的那樣,ITS序列在對刺盤孢屬真菌中部分種進行鑒定時很難將其準確區(qū)分。因此,對刺盤孢屬進行準確鑒定還需探索更多保守性較強的特異性基因位點。
本研究鑒定出的3種病原菌均為國內(nèi)首次發(fā)現(xiàn)其在自然條件下能夠侵染櫻桃葉片,引發(fā)葉部病害。本研究結(jié)果可為這3種病原菌的進一步研究和青海省櫻桃葉斑病的有效防控提供依據(jù)。
參考文獻
[1] 孫楊,韓偉,孫玉剛,等.山東地區(qū)櫻桃褐斑病病原鑒定及rDNA-ITS序列分析[J].中國農(nóng)業(yè)科技導(dǎo)報,2017,19(7):44-49.
[2] 孫楊,付全娟,孫玉剛,等.櫻桃褐斑病病原菌生物學(xué)特性及品種抗性評價[J].植物保護,2017,43(4):110-114.
[3] 黃貞光,劉聰利,李明,等.近20年國內(nèi)外甜櫻桃產(chǎn)業(yè)發(fā)展動態(tài)及對未來的預(yù)測[J].果樹學(xué)報,2014,31(S1):1-6.
[4] GUO Yinglan, LIU Xijin. Studies on the genus Pseudocercospora in China I [J]. Mycosystema, 1989, 8(2): 225-240.
[5] 徐麗,王甲威,陳新,等.甜櫻桃流膠病原菌的分子鑒定和致病性檢測[J].植物病理學(xué)報,2015,45(4):350-355.
[6] 彭子嘉,隋韻靜,沈闊程,等.四川瀘定櫻桃根癌病風(fēng)險分析[J].中國南方果樹,2018,47(5):94-96.
[7] 王燕,王春偉,王琳,等.櫻桃灰霉病菌LFD-RPA快速檢測方法的建立[J].植物病理學(xué)報,2019,20(7):1-13.
[8] 劉志恒,白海濤,楊紅,等.大櫻桃褐腐病菌生物學(xué)特性研究[J].果樹學(xué)報,2012,29(3):423-427.
[9] THOMIDIS T, TSIPOURIDIS C. First report of Alternaria leaf spot on cherry trees in Greece [J]. Plant Disease, 2006, 90(5): 680.
[10] WHARTON P S, IEZZONI A, JONES A L. Screening cherry germ plasm for resistance to leaf spot [J]. Plant Disease, 2003, 87(5): 471-477.
[11] JOSHUA J, MMBAGA M T. Perpetuation of cherry leaf spot disease in ornamental cherry [J]. Journal of Phytopathology, 2015, 163(3): 194-201.
[12] GREEN H, BENGTSSON M, DUVAL X, et al. Influence of urea on the cherry leaf spot pathogen, Blumeriella jaapii, and on microorganisms in decomposing cherry leaves [J]. Soil Biology & Biochemistry, 2006, 38(9): 2731-2742.
[13] DAZ R, ZAS R, FERNNDEZ-LPEZ J. Genetic variation of Prunus avium in susceptibility to cherry leaf spot (Blumeriella jaapii) in spatially heterogeneous infected seed orchards[J]. Annals of Forest Science, 2007, 64(1): 21-30.
[14] TEZCAN H. An outbreak of cherry leaf spot (Blumeriella jaapii[Rehm] Arx.) on sweet cherry in Canakkale province, Turkey [J]. Acta Horticulturae, 2005, 795(2): 965-968.
[15] BLUMER S. Beitrge zur Kenntnis von“Cylindrosporium padi”[J]. Phytopathologische Zeitschrift,1958, 33(3): 263-290.
[16] ANNESI T, MOTTA E, FORTI E. First report of Blumeriella jaapii teleomorph on wild cherry in Italy [J]. Plant Disease, 1997, 81(10): 1214.
[17] SILVA G S, CUTRIM F A, FERREIRA F A.Leaf spot and fruit rot of barbados cherry caused by Calonectria ilicicola [J]. Fitopatologia Brasileira, 2001, 26(1): 101.
[18] CHOI I Y, BRAUN U, PARK J H, et al. First report of leaf spot caused by Pseudocercospora pruni-persicicola on sweet cherry in Korea [J]. Plant Disease, 2014, 98(5): 693.
[19] KHAN N A, BHAT M A, AHMAD K, et al. First record of cherry leaf spot caused by Cylindrosporium padi (P. Karst.) from India [J]. Applied Biological Research, 2014, 16(1): 122-123.
[20] KUMLA J, SUWANNARACH N, LUMYONG S. First report of Phoma leaf spot disease on cherry palm caused by Phoma herbarum, in Thailand [J]. Canadian Journal of Plant Pathology, 2016, 38(1): 103-106.
[21] 趙遠征,劉志恒,李俞濤,等.大櫻桃黑斑病病原鑒定及其致病性研究[J].園藝學(xué)報,2013,40(8):1560-1566.
[22] 方中達.植病研究方法[M].第3版.北京:中國農(nóng)業(yè)出版社,1998.
[23] 張?zhí)煊?中國真菌志:鏈格孢屬[M].北京:科學(xué)出版社,2003.
[24] SUTTON B C. The Coelomycetes [M]. England: Commonwealth Mycological Institute, 1971.
[25] MURRAY M, THOMPSON W. Rapid isolation of high molecular weight plant DNA [J]. Nucleic Acids Research, 1980, 8(19): 4321-4326.
[26] GLASS N L, DONALDSON G C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes [J]. Application Environment Microbiology, 1995, 61(4): 1323-1330.
[27] CARBONE I, KOHN L M. A method for designing primer sets for speciation studies in filamentous ascomycetes [J]. Mycologia, 1999, 91(3): 553.
[28] HONG S G, CRAMER R A, LAWRENCE C B, et al. Alt a 1 allergen homologs from Alternaria and related taxa: analysis of phylogenetic content and secondary structure [J]. Fungal Genetics and Biology, 2005, 42(2): 119-129.
[29] 曾向萍,符美英,何舒,等.大麻葉斑病病原菌鑒定[J].分子植物育種,2018,16(21):7094-7098.
[30] 陳冠州,沈會芳,林壁潤,等.蜘蛛蘭紅斑病的病原鑒定[J].廣東農(nóng)業(yè)科學(xué),2015,42(9):73-76.
[31] 曲文文,劉霞,楊克強,等.山東省危害核桃的鏈格孢屬真菌鑒定及其系統(tǒng)發(fā)育[J].植物保護學(xué)報,2012,39(2):121-128.
[32] 劉春來,文景芝,楊明秀,等.rDNA-ITS在植物病原真菌分子檢測中的應(yīng)用[J].東北農(nóng)業(yè)大學(xué)學(xué)報,2007,38(1):101-106.
[33] 宋順華,馬樂樂,寧利園,等.北京西瓜嫁接苗褐斑病病原鑒定[J].植物保護學(xué)報,2017,44(5):865-866.
[34] HONG S G, MACCARONI M, FIGULI P J, et al. Polyphasic classification of Alternaria isolated from hazelnut and walnut fruit in Europe[J]. Mycological Research, 2006, 110(11): 1290-1300.
[35] 趙金梅,高貴田,谷留杰,等.中華獼猴桃褐斑病病原鑒定及抑菌藥劑篩選[J].中國農(nóng)業(yè)科學(xué),2013,46(23):4916-4925.
[36] 趙艷琴,于華榮,石凱,等.高粱鏈格孢葉斑病病原鑒定[J].植物病理學(xué)報,2017,47(2):282-285.
[37] 趙圓,王玲杰,王雪峰,等.雜柑褐斑病的病原鑒定[J].果樹學(xué)報,2014,31(2):292-295.
[38] 岳海梅,張榮,孫廣宇.小孢子鏈格孢endoPG基因核苷酸序列分析及系統(tǒng)發(fā)育研究[J].菌物學(xué)報,2007,26(2):202-210.
[39] 張榮,岳海梅,孫廣宇.小孢子鏈格孢OPA2-1核苷酸序列分析及系統(tǒng)發(fā)育研究[J].菌物學(xué)報,2009,28(4):553-557.
[40] 岳海梅,莊華,孫廣宇,等.銀杏內(nèi)生小孢子鏈格孢的分離與鑒定[J].西北農(nóng)林科技大學(xué)學(xué)報(自然科學(xué)版),2010,38(7):191-196.
[41] 符丹丹.中國蘋果炭疽病病原菌的遺傳多樣性[D].楊凌:西北農(nóng)林科技大學(xué),2014.
(責(zé)任編輯: 田 喆)
