SUMO特異性蛋白酶3通過調控巨噬細胞極化促進磷酸鈣誘導的小鼠腹主動脈瘤形成*
2020-06-09 02:41:38張永興倪煥爾李偉峰
中國病理生理雜志 2020年5期
關鍵詞:小鼠
陳 陽, 張永興, 倪煥爾, 李偉峰, 汪 芳
(上海交通大學附屬第一人民醫院心內科,上海200080)
腹主動脈瘤(abdominal aortic aneurysm,AAA)是腹主動脈的永久性局部擴張至正常腹主動脈直徑的150%[1]。臨床AAA多發生在腎下動脈處[2];破裂性AAA及其導致的相關病理生理損害的死亡率超過80%[3]。目前除了手術修復,尚無藥物可以治療。因此,探究AAA的病理生理學調控機制,對于臨床腹主動脈瘤的藥物防治靶點研發具有重要意義。
在AAA中,病理改變可見血管平滑肌細胞的凋亡和缺失、炎癥細胞的浸潤以及基質金屬蛋白酶(matrix metalloproteinases,MMPs)表達上調等引發彈性蛋白斷裂和細胞外基質(extracellular matrix,ECM)的降解[4]。單核-巨噬細胞是參與AAA免疫應答重要的炎癥細胞,在特定的免疫微環境下,骨髓單核細胞(M0)可以分化為促炎的巨噬細胞(M1型)或抗炎的巨噬細胞(M2型)[5]。而MMPs和許多促炎癥反應因子,如腫瘤壞死因子α(tumor necrosis factorα,TNFα)等,由M1型巨噬細胞分泌產生,而M2型巨噬細胞則分泌抗炎細胞因子,如白細胞介素10(interleukin-10,IL-10)等。巨噬細胞M1/M2的失衡被認為是AAA發生發展的重要環節[6],同時也有研究報道巨噬細胞極化可以調控新生血管形成[7]。巨噬細胞在AAA免疫應答中發揮至關重要的作用[8],因此調控巨噬細胞極化的信號通路對于研究AAA發病機制具有重要意義。
SUMO(small ubiquitin-related modifier)是一系列重要的類泛素蛋白,能對底物進行翻譯后SUMO修飾(SUMOylation)[9]。SUMO 修飾是一個可逆過程,SUMO特異性蛋白酶家族(sentrin/SUMO-specific proteases,SENPs)能夠去除底物蛋白的SUMO修飾,或影響底物與其它蛋白質的相互作用,從而影響底物的生物學效應。有研究表明,SENP3敲除減少了脂多糖(lipolysaccharide,LPS)誘導的巨噬細胞Toll樣受體(Toll-like receptor,TLR4)炎癥信號的激活和促炎因子的產生,提高小鼠存活率[10]。目前已有證據顯示TLR4炎癥信號的激活能促進巨噬細胞的極化,加重炎癥損傷[11-12]。此外,TLR4激活促進AAA的調控關系也已被證明[13]。但SENP3在AAA病程中對巨噬細胞極化的影響尚不清楚。
本課題組構建SENP3單核細胞特異性敲除的小鼠,并通過磷酸鈣(calciumphosphate,CaPO4)誘導的AAA動物模型,探討SENP3通過影響巨噬細胞的功能在AAA發生發展中的作用,以期進一步闡釋調控AAA發生和進展的潛在機制。
材料和方法
1 動物
C57BL/6野生型(wild type,WT)小鼠購于上海斯萊克動物公司。C57BL/6背景的Senp3flox/flox(以下簡稱WT)小鼠由南京大學模式動物研究中心產生,與C57BL/6背景的Lyz2-Cre小鼠雜交產生Senp3flox/flox;Lyz2-Cre(SENP3單核細胞特異性敲除,即條件性敲除,conditioned knockout,cKO)小鼠。嚴格按照中華人民共和國科學技術部發布的《實驗室動物的護理和使用指南》中的規定進行動物實驗,該方案已獲得上海交通大學醫學院附屬機構動物護理和使用委員會的批準[使用許可證號為SYXK(滬)2018-0027],所有手術均在戊巴比妥鈉麻醉下進行,并盡一切努力使動物痛苦最小化。本研究選用SPF級cKO小鼠以及同窩出生的WT小鼠,雄性,8~12周齡,體重20~22 g。小鼠飼養于上海交通大學醫學院動物實驗中心,飼以國家標準嚙齒類動物飼料,溫度為(23±2)℃,濕度為50%±10%。
2 主要試劑
小鼠SENP3、β-actin、IL-1β和IL-6抗體購自Cell Signaling Technology;小鼠 CD206、CD86和 TNFα 抗體購自Abcam;小鼠MMP-9抗體購自Affinity;熒光素標記的II抗(Alexa Fluor?488標記的山羊抗鼠IgG、Alexa Fluor?555標記的山羊抗鼠IgG、Alexa Fluor?488標記的山羊抗兔IgG和Alexa Fluor?555標記的山羊抗兔IgG)購自Invitrogen;生物素化羊抗小鼠IgG(1∶500)和生物素化羊抗兔IgG(1∶500)購自Jackson;免疫組化顯色試劑盒購自Vector Laboratories;胎牛血清(fetal bovine serum,FBS)購自Gibco;鼠源巨噬細胞集落刺激因子(macrophage colony-stimulating factor,M-CSF)購自PeproTech;多聚甲醛購自Sigma;Western blot化學發光顯色劑購自中國上海圣爾生物科技有限公司;二水氯化鈣(calcium chloride dihydrate,CaCl2·2H2O)、磷 酸 鹽 緩 沖 液(phosphatebuffered saline,PBS)和Tween-20購自中國生工生物工程(上海)股份有限公司;TBS緩沖液購自上海索萊寶生物科技有限公司;二氫乙啶(dihydroethidium,DHE)染色液購自中國上海碧云天生物技術有限公司;戊巴比妥購自Sigma。
3 主要方法
3.1 改良磷酸鈣誘導的小鼠腎下AAA模型 參照現有研究,相對于傳統CaCl2模型,改良方法依次使用CaCl2和PBS誘導產生動脈瘤所需時間較短[14]。我們選用同窩出生、C57BL/6背景的8~12周齡雄性小鼠,按基因型分成2組:WT組(n=20)和cKO組(n=18)。小鼠在戊巴比妥(100 mg/kg)麻醉下,無菌環境里先用0.5 mol/L CaCl2浸泡的棉球包裹游離的腹主動脈孵育10 min,隨后改用PBS浸泡的棉球孵育5 min。消毒紗布小心吸干殘余液體,用無菌生理鹽水沖洗腹腔,減少其余組織損傷,結扎出血點,關腹部縫合。造模時間為14 d。手術操作交由未知小鼠基因型與具體分組的人來完成。
3.2 實驗小鼠標本的收集、處理和保存 在術后第2周,戊巴比妥麻醉小鼠后剖腹,體式顯微鏡下小心暴露腹主動脈,剝離腹主動脈動脈周圍筋膜和脂肪組織,游離升主動脈至腹主動脈髂總動脈分叉處,游標卡尺測此處動脈直徑。部分標本放入凍存管中存放于液氮罐中,用于提取組織RNA進行RT-qPCR實驗,以及提取組織蛋白進行Western blot實驗。另一部分標本在操作過程中小心保持形態完整,在4%多聚甲醛溶液中固定48 h,再分成兩部分:(1)其中一些標本經過常規脫水、透明、浸蠟、包埋制成蠟塊,將蠟塊制成4~5μm厚的連續切片,供HE染色、Verhoeff-van Gieson(VVG)特殊染色及免疫組織化學染色;(2)另一些標本用最優切片溫度(optimum cutting temperature,OCT)包埋劑包埋,制成動脈橫截面7 μm厚的連續冰凍切片,用于免疫熒光和活性氧簇(reactive oxygen species,ROS)的DHE染色。
3.3 免疫組織化學染色法 石蠟切片置于60℃烘箱1 h,置于二甲苯I和二甲苯II中各15 min,再依次浸泡100%乙醇、100%乙醇、95%乙醇、95%乙醇、75%乙醇和雙蒸水各5 min完成脫蠟。脫蠟后,石蠟切片置于抗原修復液(檸檬酸三鈉緩沖液,pH 6.0)中,95℃水浴修復6 min。PBS漂洗后封閉,經I抗(兔抗小鼠SENP3多克隆抗體)4℃孵育過夜,生物素標記的II抗(生物素化的羊抗兔IgG)室溫孵育2 h后在顯色液 A+B(1∶1)下孵育 1 h,DAB 顯色 5~10 min。適時終止后,經梯度濃度乙醇脫水和二甲苯處理,用中性樹膠封片,在顯微鏡下觀察和拍照。
3.4 小鼠骨髓原代單核細胞(bone marrow-derived monocytes,BMDMs)的提取和誘導 小鼠麻醉后,脫頸處死,無菌條件下取小鼠下肢骨,DMEM培養基沖出骨髓,過40μm濾網,室溫下500×g離心5 min,去除上清液后用含10%FBS和30μg/L M-CSF的DMEM培養基重懸,接種到10 cm培養皿,7 d后細胞分化為M0,之后分別用來進行M1極化誘導[100 μg/L LPS+50μg/L干擾素γ(interferonγ,IFNγ)處理24 h]和 M2 極化誘導(20 μg/L IL-4 處理 24 h)[6,15]。BMDMs置于37°C、5%CO2培養箱,隔天換液。
3.5 組織和細胞免疫熒光法 (1)組織免疫熒光:脫蠟步驟同3.4中所述,其余步驟與細胞免疫熒光步驟相似。(2)細胞免疫熒光:細胞用PBS清洗之后,迅速加含4%多聚甲醛的固定液,于4°C固定過夜;0.2%Triton X-100處理10 min,并用封閉液封閉1 h;CD86抗體(1.0 mg/L)4°C孵育過夜,次日用PBS洗3次后,熒光素標記的山羊抗兔IgG(1∶500)在室溫下進行染色標記,然后觀察。細胞圖像通過DM2500型熒光顯微鏡(Leica)拍照。
3.6 ROS的DHE染色法 DHE在ROS作用下脫氫后,可與RNA或DNA結合產生紅色熒光。冰凍切片用PBS清洗3次,每次5 min后,按1∶100工作濃度用PBS稀釋配制DHE染色液,每個樣品滴加50μL工作液,室溫下避光孵育60 min。然后同樣用PBS清洗3次,每次5 min。熒光顯微鏡觀察(最大激發波長為300 nm,最大發射波長為610 nm)。
3.7 組織RNA抽提和RT-qPCR檢測 TRIzol一步法提取小鼠腹主動脈瘤體組織中的RNA后,利用NanoDrop 2000分光光度計檢測樣品RNA濃度,并注意A260/A280是否在1.8~2.0范圍,以判斷RNA提取過程中是否有污染。之后按照TaKaRa的PrimeScript?RT reagent Kit使用說明書,進行逆轉錄操作。qPCR按照TaKaRa SYBR?Premix Ex Taq?(Perfect Real Time)Kit說明書,在冰上將擴增體系中的各組分充分混勻后,使用LightCycler?480型實時熒光定量PCR儀檢測。采用兩步法熒光定量PCR擴增程序進行擴增。反應結束后檢查實時熒光定量PCR的擴增曲線是否正常,融解曲線是否為單峰。計算公式:ΔCt=Ct目的基因-Ct內參照(GAPDH);ΔΔCt=ΔCt待測樣品-ΔCt對照樣品;相對表達量=2-ΔΔCt。引物由中國生工生物有限公司負責合成,主要引物序列見表1。

表1 RT-qPCR引物序列Table 1.Sequences of the primers for RT-qPCR
3.8 蛋白提取和Western blot RIPA緩沖液提取來自主動脈或細胞的蛋白。將主動脈組織速凍在液氮中,在RIPA緩沖液中通過組織研磨破碎儀粉碎,冰上靜置40~60 min,12 000×g、4℃離心15 min。然后將上清液轉移到新EP管用于后續實驗。BCA protein assay kit測定其蛋白濃度,再根據樣品蛋白濃度,按蛋白總量30~50μg范圍內計算出上樣體積。Western blot檢測:組織蛋白樣品經SDS-PAGE后轉至聚偏二氟乙烯膜,5%脫脂奶粉37℃封閉1 h,TBST緩沖液洗3次,每次5 min,然后加入I抗(SENP3、TNFα、IL-1β和MMP9抗體均為1∶1 000稀釋,β-actin抗體為1∶5 000稀釋)于4℃孵育過夜;次日,加入II抗(1∶10 000)于37℃孵育1 h;用增強化學發光(enhanced chemiluminescence,ECL)試劑顯影,顯影儀采用ImageQuant LAS4000(GEHealthcare)。
4 統計學處理
以均數±標準差(mean±SD)表示符合正態分布的計量資料,采用GraphPad Prism 8.0進行統計分析和作圖。兩組間符合正態分布的計量資料比較采用雙側t檢驗,兩組間等級資料比較采用Mann-WhitneyU檢驗。兩個率或兩個構成比的比較采用Fisher確切概率法,生存分析比較采用Mantel-Cox檢驗。以P<0.05為差異有統計學意義。
結 果
1 SENP3調控巨噬細胞極化
RT-qPCR結果顯示,SENP3 mRNA在M1型巨噬細胞誘導過程中表達升高(P<0.01),但是M2型巨噬細胞中表達顯著降低(P<0.01),見圖1A、B。細胞免疫熒光圖片中可見M1分化時細胞核內SENP3蛋白表達升高,見圖1C。此外,在巨噬細胞受刺激由M1向M2型轉換時,SENP3的mRNA表達降低(P<0.01),而在M2向M1型轉換時,SENP3的mRNA表達升高(P<0.01),見圖1D、E。進一步,細胞免疫熒光結果顯示,cKO小鼠和其同窩出生的WT小鼠的原代BMDMs在誘導M1分化過程中,cKO組M1型巨噬細胞的標志物CD86表達低于WT組,與之對應的是,誘導M2分化時cKO組M2型巨噬細胞的標志物CD206表達則高于WT組,見圖1F。

Figure 1.SENP3 expression was up-regulated during M1 polarization and M1/M2 transformation.A and B:the BMDMs from wildtype(WT)mice were isolated,cultured and treated with M1 stimulation(100 μg/L LPS+50 μg/L IFNγ)(A)or M2 stimulation(20μg/L IL-4)(B)for 24 h,and the mRNA expression of SENP3 was detected by RT-qPCR;C:the BMDMs from WTmice were treated with M1 stimulation(100μg/L LPS+50μg/L IFNγ)or M2 stimulation(20μg/L IL-4)for 24 h,and immunofluorescence was conducted to analyze SENP3 protein expression(red:SENP3;blue:DAPI);D:the BMDMs from WT mice were cultured in DMEM containing 100μg/L LPS+50μg/L IFNγfor 24 h,and then in DMEM containing 100μg/L LPS+50μg/L IFNγor 20μg/L IL-4 for another 24 h,and the mRNA expression of SENP3 was detected by RT-qPCR;E:the BMDMs from WT mice were cultured in DMEM containing 20μg/L IL-4 for 24 h,and then in DMEM containing 20μg/L IL-4 or 100μg/L LPS+50μg/L IFNγfor another 24 h,and the mRNA expression of SENP3 was detected by RT-qPCR;F:BMDMs isolated from WT and cKO mice were cultured and treated with M1 stimulation(100μg/L LPS+50μg/L IFNγ)or M2 stimulation(20μg/L IL-4)for 24 h,and the M1 marker or M2 marker was measured by immunofluorescence(red:CD206;green:CD86;blue:DAPI).Mean±SD.n=6.**P<0.01 vs PBSgroup;##P<0.01 vs LPS+IFNγ/LPS+IFNγ group;△△P<0.01 vs IL-4/IL-4 group.圖1 巨噬細胞M1極化和M1/M 2轉化過程中SENP3表達上調
2 SENP3在改良磷酸鈣誘導的模型小鼠AAA組織中高表達
RT-qPCR和Western blot結果顯示,SENP3在小鼠AAA組織中表達顯著升高(P<0.05),見圖2A、B。石蠟切片免疫組織化學染色顯示,AAA病灶組織內膜、中膜層有缺失;SENP3陽性染色主要分布在中膜層細胞核及核周的部分胞質內,與對照組相比,陽性染色面積顯著增大(P<0.05),見圖2C。
3 單核細胞SENP3特異性敲除抑制磷酸鈣誘導的小鼠A A A形成

Figure 2.SENP3 was up-regulated in the aorta tissue of mice with CaPO4-induced abdominal aortic aneurysm(AAA).The AAA model was establshed in 8~12-week-old C57BL/6Jmice by administration with CaCl2 and PBSfor 14 d.A:SENP3 mRNA expression was measured by RT-PCR;B:SENP3 protein expression was measured by Western blot;C:representative immunohistochemical staining images for SENP3 in paraffin-embedded sections from aorta tissues were shown,and the SENP3 positive area was determined.Mean±SD.n=6.*P<0.05,**P<0.01 vs Con group.圖2 SENP3在CaPO 4誘導的小鼠腹主動脈瘤中高表達
RT-qPCR和Western blot檢測結果顯示,cKO小鼠BMDMs中,SENP3表達顯著降低(P<0.01),見圖3A。cKO雄鼠和同窩出生的WT雄鼠AAA造模結果顯示,cKO小鼠腹主動脈成瘤較小或無法形成AAA,見圖3B,相應地,cKO組AAA造模小鼠生存率顯著高于WT組(P<0.05),見圖3C。改良磷酸鈣法誘導的AAA中cKO小鼠AAA發生率顯著低于WT組(P<0.05),見圖3D;WT小鼠中,磷酸鈣誘導的AAA主動脈瘤體外部直徑和動脈重/總體重比值均高于cKO組(P<0.05),見圖3E、F。HE染色可見正常動脈具備清晰、完整的彈力板層將內、中、外膜分開,動脈無異常擴張,內皮細胞和平滑肌細胞排列整齊;而造模后,WT組發生AAA的動脈擴張和增厚程度比cKO組嚴重,見圖3G。此外,VVG染色可見cKO組僅有少量或無內彈性膜崩解及彈性纖維斷裂(黑色著色為彈性纖維),而WT組的彈性纖維斷裂更為嚴重,纖維斷裂等級數據的差異有統計學顯著性(P<0.05),見圖3G、H。
4 SENP3敲除抑制AAA的炎癥和氧化應激
通過對AAA瘤體組織進行RT-qPCR和Western blot檢測顯示,在磷酸鈣誘導下,WT組的主動脈組織炎癥因子IL-1β、IL-6和TNFα的表達顯著高于cKO組,見圖4A~C。DHE染色檢測動脈組織中的ROS水平,結果顯示:cKO組的主動脈組織中氧化應激水平低于WT組,見圖4D。
5 SENP3敲除抑制MMP-9表達上調所導致的ECM重構
RT-qPCR、Western blot和免疫組織化學染色結果顯示,在cKO組瘤體內MMP-9的mRNA和蛋白表達比WT組顯著下調,見圖5A~C。MMP-9陽性著色在細胞質中,瘤體的病灶中伴隨著大量浸潤的炎癥細胞和破損的血管平滑肌細胞,而cKO組的中膜層相對完整,在MMP-9高表達的相應部位,對應WT組瘤體動脈中膜的彈力板層破壞更為嚴重,見圖5C。
6 SENP3介導MKK7 de-SUMO修飾調控JNK通路和MMP-9表達
Co-IP實驗結果顯示,巨噬細胞MKK7的SUMO2/3修飾在誘導M1分化的過程中減少(圖6A),而MKK7與SENP3的相互作用增強(圖6B);在轉染Flag-MKK7野生型(Flag-MKK7 WT)和SUMO修飾位點K18突變體(Flag-MKK7 K18Rmutant)后誘導BMDMs進行M1極化,結果顯示,后者p-JNK和MMP-9水平均高于前者,見圖6C、D。
討 論
1 SENP3介導的巨噬細胞M 1極化促進AAA發生和進展
哺乳動物體內目前共發現有7種SENPs,其中SENP3對氧化應激最為敏感[16-17],主要介導底物蛋白的去除SUMO2/3修飾(de-SUMO2/3修飾),在腫瘤增殖、轉移和炎癥細胞的激活中發揮重要作用。同時,在免疫應答條件下巨噬細胞極化成M1型,線粒體中大量生成ROS,在氧化應激環境下分泌釋放大量的炎癥因子,放大炎癥損傷[5]。本研究表明SENP3在巨噬細胞M1/M2極化時存在表達差異,并且敲除SENP3能改變M1/M2巨噬細胞的誘導分布,這提示SENP3也參與調控巨噬細胞M1和M2表型轉換。人類外周血單核細胞衍生的巨噬細胞蛋白質組學和轉錄組學分析顯示,AAA與非AAA外周動脈疾病的患者之間,巨噬細胞的蛋白和基因表達譜存在顯著差異,其中差異表達的基因和蛋白質富集在與AAA相關的ECM重塑和炎癥等方面,表明單核-巨噬細胞譜系在AAA的ECM重塑或炎癥過程具有重要作用[18]。在體外,cKO小鼠的M0受刺激后發生M1極化的比例降低,而在磷酸鈣誘導的腹主動脈瘤小鼠模型中,SENP3敲除能夠有效降低AAA的氧化應激損傷、炎癥反應和ECM重塑,從而導致AAA發病減少和小鼠存活率的升高。以上結果表明,SENP3介導的巨噬細胞M1極化可促進AAA發生和進展。

Figure3.Monocyte-specific SENP3 knockout protectsed mice from CaPO4-induced AAA.A:SENP3 expression in abdominal aorta tissue was measured by RT-qPCR and Western blot;B:representative images of mouse abdominal aorta tissue in 8~12-week-old cKO male mice(n=6)and WT littermates(n=6)administered with CaCl2 and PBSfor 14 d(the arrowhead indicates typical aneurysm);C:Kaplan-Meier survival curves(WT:n=20;cKO:n=15);D:incidences of AAA in animal model;E:quantification analysis for maximal external diameters of abdominal aortas in animal model;F:total wet weight of abdominal aorta per body weight in animal model;G:HE and VVG staining for paraffin-embedded section of AAA tissue in animal model;H:grade of elastic degradation.Unpaired two-tailed t-test was used for statistical analysis in A,Eand F,and Mann-Whitney U test was used in H.Two-sided Fisher's exact test was used to compare variables in D.Statistical significance on Kaplan-Meier curves(C)was assessed with Mantel-Cox test.Mean±SD.*P<0.05,**P<0.01 vs WTgroup.圖3 單核細胞SENP3特異性敲除減少CaPO4誘導的小鼠腹主動脈瘤形成
2 SENP3促進巨噬細胞炎癥反應并調控ECM的機制

Figure 4.Monocyte-specific SENP3 knockout inhibited the flammation and oxidative stress in AAA.A,Band C:the mRNA and protein levels of inflammatory cytokines(IL-1β,IL-6 and TNFα)in the aorta tissues of WT and cKO mice were detected by RT-qPCR and Western blot;D:in situ dihydroethidium(DHE)staining for ROSproduction in the frozen sections of the aorta tissues from WT and cKOmice.Unpaired two-tailed t-test was used for statistical analysis in A,B and C.Mean±SD.n=5.**P<0.01 vs WTgroup.圖4 單核細胞SENP3特異性敲除抑制AAA的炎癥和氧化應激反應

Figure 5.Monocyte-specific SENP3 knockout inhibited MMP-9-induced ECM remodeling.A:the mRNA level of MMP-9 was measured by RT-qPCR;B:the protein level of MMP-9 was measured by Western blot;C:immunohistochemical staining for MMP-9 in paraffin-embedded sections fromaorta tissues.Mean±SD.n=5.*P<0.05,**P<0.01 vs WTgroup.圖5 單核細胞SENP3特異性敲除抑制MMP-9介導的細胞外基質重構
SENP3及其介導的de-SUMO修飾在單核-巨噬細胞分化和炎癥反應中發揮重要作用。SUMO蛋白酶SENP3在小鼠炎癥性休克模型中可以激活TLR4下游MAPK/AP-1通路,正向調節巨噬細胞炎癥反應,而SENP3敲除則顯著抑制了這一過程,提高了小鼠炎癥損傷后的存活率[10]。我們的研究結果表明相,磷酸鈣誘導的AAA病灶中有大量炎癥細胞浸潤,SENP3介導巨噬細胞M1極化發揮促炎作用從而加重AAA。研究表明,鈣離子可誘導主動脈中層平滑肌細胞凋亡和彈性蛋白降解,刺激單核-巨噬細胞遷移趨化,并促進炎癥因子表達分泌[19-20]。M1型巨噬細胞的MMPs(MMP-9最具代表性)表達上調,能夠降解和破壞中層彈性纖維板以及Ⅰ型和Ⅲ型膠原,反映病情進展。病灶處炎癥損傷-修復過程ECM的主動病理性重構依賴于MMPs的分泌和活化,大量降解動脈中層的主要應力結構——彈力蛋白。同時,炎癥細胞對組織的浸潤和相關炎癥因子表達上調會促進巨噬細胞的MMPs進一步表達,加重彈力蛋白的降解,從而引發破壞-重塑惡性循環,促進AAA的形成和動脈瘤體破裂。我們構建的小鼠模型表明,單核細胞SENP3敲除顯著減輕了這種炎癥級聯放大效應,延緩AAA的疾病進展。
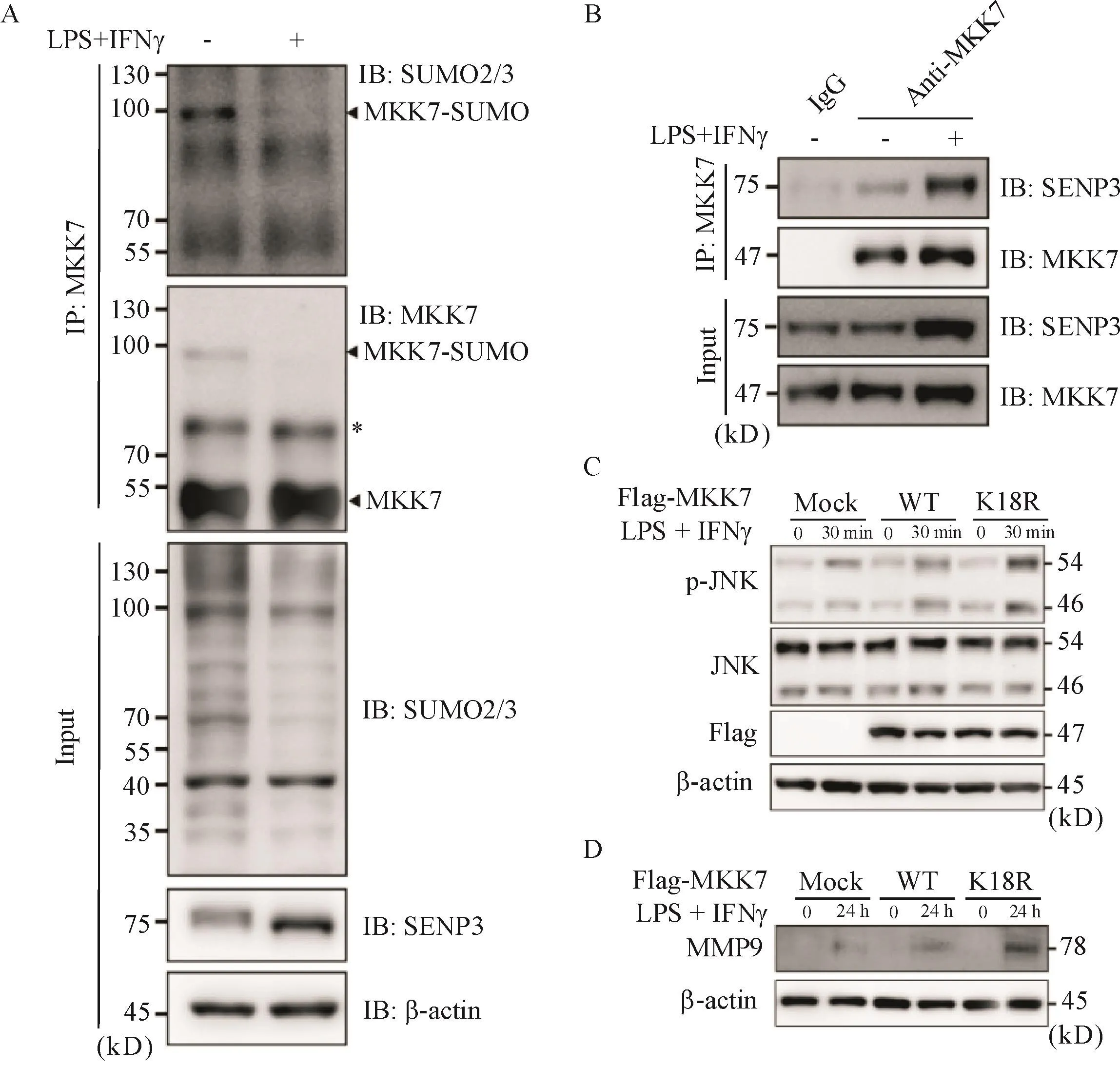
Figure 6.SENP3 mediated MKK7 de-SUMO modification to regulate JNK pathway and MMP-9 expression.A:BMDMs treated with or without LPS(100 μg/L)and IFNγ(50μg/L)for 48 h were collected,and endogenous MKK7 conjugated with SUMO2/3 was detected by co-immunoprecipitation(Co-IP);B:BMDMs were treated with LPS(100 μg/L)and IFNγ(50 μg/L)for 30 min,and the interaction of endogenous SENP3 and MKK7 was determined by Co-IP;C and D:BMDMs were transfected with Flag-tagged MKK7 WT or SUMOless mutant K18R and incubated with or without LPS(100 μg/L)and IFNγ(50μg/L)for the indicated time,and then p-JNK,JNK and MMP-9 were determined by immunoblotting(IB).*:non-specific bands.圖6 SENP3通過介導MKK 7的de-SUMO修飾調控JNK通路和MMP-9表達
這些研究也揭示了不同炎癥環境下的單核-巨噬細胞系中SUMO修飾底物和多種調控機制,為我們深入了解單核-巨噬細胞譜系中轉錄因子的翻譯后調控機制提供了充分的證據。目前尚無研究表明MMP-9能被SUMO化修飾,SUMO修飾功能位點預測(http://www.jassa.fr/)也同樣表明,MMP-9沒有可能的SUMO修飾位點。因此,巨噬細胞MMP-9合成增多可能是SENP3調控的效應分子和終末環節。SENP3作為一種de-SUMO蛋白酶,主要功能是對底物蛋白進行翻譯后de-SUMO修飾,降低底物蛋白的SUMO2/3水平,從而影響底物蛋白的功能及活性。我們課題組前期研究表明,SENP3通過介導MKK7 de-SUMO修飾調控JNK/c-Jun通路,從而參與巨噬細胞炎癥反應[10]。我們現有的研究證據也表明SENP3作用于AAA疾病的機制在于de-SUMO調控MKK7及MAPK/JNK通路,進而促進MMP-9合成和分泌,最終導致ECM重構的病理學改變。未來我們將進一步通過Co-IP實驗鑒定SENP3及其介導的de-SUMO2/3在氧化應激以及CaPO4誘導的AAA中的其他作用底物。此外,其他類型細胞如平滑肌細胞的凋亡和增殖異常,以及內皮細胞參與的新生血管形成[7]等,在AAA形成中也發揮了重要作用,我們將在接下來的工作中繼續開展相關研究。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
