P2RX7介導(dǎo)的細(xì)胞焦亡和凋亡參與TSA緩解的煙草煙霧暴露所致的小鼠子宮損傷過(guò)程
2020-06-11 07:49:20叢艷飛丁晶晶王莉莉
中國(guó)藥理學(xué)通報(bào) 2020年6期
叢艷飛,丁晶晶,李 芳,王莉莉
(中國(guó)醫(yī)科大學(xué)附屬盛京醫(yī)院實(shí)驗(yàn)研究中心, 遼寧省環(huán)境與代謝疾病動(dòng)物模型研究與應(yīng)用重點(diǎn)實(shí)驗(yàn)室, 遼寧 本溪 117000)
近年來(lái),主動(dòng)吸煙和被動(dòng)吸煙的人數(shù)在全世界范圍內(nèi)增加,女性暴露于二手煙的現(xiàn)象非常普遍,育齡期婦女吸煙率呈明顯上升趨勢(shì)[1]。在吸煙時(shí),煙草煙霧(cigarette smoke,CS)中的有害物質(zhì)會(huì)通過(guò)呼吸道的粘膜進(jìn)入人體的各個(gè)器官和組織造成全身性的傷害,例如CS中的有毒物質(zhì)通過(guò)降低胚胎植入率,減少卵巢中類(lèi)固醇激素的生成和卵泡數(shù)量等,影響女性生殖的各個(gè)階段[2]。CS混合物中的有毒物質(zhì)也可導(dǎo)致子宮發(fā)育受損,子宮是哺乳動(dòng)物重要的生殖器官,主要由3種類(lèi)型的細(xì)胞(上皮細(xì)胞,基質(zhì)細(xì)胞和子宮肌層細(xì)胞)組成。研究發(fā)現(xiàn),口服尼古丁會(huì)導(dǎo)致大鼠子宮腔上皮和腺上皮形態(tài)學(xué)改變[3],CS暴露可導(dǎo)致子宮基質(zhì)金屬肽酶 9(matrix metalloproteinase 9,MMP-9),4 型 CXC 趨化因子受體(recombinant chemokine C-X-C-motif receptor 4,CXCR4)和雌激素受體(estrogen receptor,ER)的表達(dá)增加[4]。曲古抑菌素 A(trichostatin A,TSA)能夠改變蛋白質(zhì)乙酰化水平,對(duì)多種疾病都有治療作用,例如TSA能夠緩解腎上腺嗜鉻細(xì)胞瘤造成的缺血性損傷[5]。在我們先前的研究中,CS暴露通過(guò)激活組蛋白脫乙酰基酶1/2(histone deacetylase,HDAC)導(dǎo)致子宮組織形態(tài)發(fā)生變化,TSA 通過(guò)抑制 HDAC 1/2 和 mTOR 的激活,重新激活自噬,從而引起子宮組織形態(tài)改變[6]。由于CS含有多種復(fù)雜的有害成分,CS暴露對(duì)子宮損傷的機(jī)制尚未完全清楚。
嘌呤受體(purinergic receptor P2X,ligand-gated ion channel,7,P2RX7)是由細(xì)胞外三磷酸腺苷(eATP)激活的離子通道,在與天然配體胞外 ATP(eATP) 結(jié)合后介導(dǎo)細(xì)胞內(nèi)鈉和鈣的內(nèi)流以及鉀的外流。在子宮組織中,P2RX7 主要在子宮內(nèi)膜,子宮頸內(nèi)膜和外宮頸的上皮細(xì)胞中表達(dá)[7]。在大鼠子宮肌層細(xì)胞中 ATP 誘導(dǎo)的電流通過(guò)P2RX7 受體穿過(guò)細(xì)胞膜導(dǎo)致子宮收縮[8]。P2RX7 受體可以引起 Ca2+的強(qiáng)烈增加和胱冬肽酶(cysteinyl aspartate-specific proteases,caspase)的裂解來(lái)啟動(dòng)細(xì)胞凋亡,也可以誘導(dǎo) NLRP3 炎性體和 caspase-1 的激活導(dǎo)致促炎因子的分泌,最終發(fā)生細(xì)胞焦亡。研究發(fā)現(xiàn),P2X7-NLRP3/ASC-caspase1/11-IL-1β/IL-18 軸在CS暴露導(dǎo)致的小鼠過(guò)敏性氣道炎癥中發(fā)揮重要作用[9]。此外,P2RX7 受體還可通過(guò) caspase-9 介導(dǎo)的線(xiàn)粒體途徑誘導(dǎo)人宮頸上皮細(xì)胞凋亡[10]。綜上所述,P2RX7 受體在子宮組織中發(fā)揮重要的作用,而P2RX7 受體介導(dǎo)的細(xì)胞焦亡和凋亡是否參與了CS暴露導(dǎo)致的子宮損傷?P2RX7 受體是否參與TSA對(duì)CS暴露導(dǎo)致的子宮組織損傷的緩解作用?目前尚需要進(jìn)一步探索。
1 材料與方法
1.1 材料
1.1.1實(shí)驗(yàn)動(dòng)物 SPF 級(jí) 60 d C57 BL/6 小鼠 36 只,♀,體質(zhì)量(20±0.5)g,購(gòu)自遼寧長(zhǎng)生生物技術(shù)有限公司[SCXK(遼)2015-0001]。實(shí)驗(yàn)在中國(guó)醫(yī)科大學(xué)附屬盛京醫(yī)院動(dòng)物中心 SPF 級(jí)動(dòng)物飼養(yǎng)室[SYXK(遼)2017-0004]進(jìn)行,實(shí)驗(yàn)動(dòng)物飼養(yǎng)在室溫(22 ±3)℃,相對(duì)濕度 (50±20)%,暗循環(huán),噪聲< 60 dB,氨≤1.5×10-6的SPF級(jí)房間中,自由喂食實(shí)驗(yàn)動(dòng)物飼料和水。針對(duì)實(shí)驗(yàn)動(dòng)物的所有操作流程關(guān)注動(dòng)物福利,并通過(guò)中國(guó)醫(yī)科大學(xué)附屬盛京醫(yī)院醫(yī)學(xué)倫理委員會(huì)批準(zhǔn)(2013PS06K;2019PS263K)。實(shí)驗(yàn)前詞養(yǎng) 1 周,觀察無(wú)明顯全身疾病及其他異常后開(kāi)始實(shí)驗(yàn)。CS暴露期間每2 d測(cè)量1次小鼠體重 。
1.1.2試劑 雄獅牌香煙(每支含0.7 mg 尼古丁和 8 mg 焦油);TSA(Sigma;批號(hào):T1952,規(guī)格:200 μL);一抗來(lái)源:GAPDH(康成生物;批號(hào):KC-5G4,規(guī)格:100 μL);P2RX7(proteintech;批號(hào):11144-1-AP,規(guī)格:150 μL);NLRP3(abcam;批號(hào):ab214185,規(guī)格:100 μL);ASC(萬(wàn)類(lèi)生物;批號(hào):WL02462,規(guī)格:50 μL);cleaved caspase-1(萬(wàn)類(lèi)生物;批號(hào):WL02996a,規(guī)格:50 μL);IL-1β(萬(wàn)類(lèi)生物;批號(hào):WLH3903,規(guī)格:50 μL);cleaved caspase-3(萬(wàn)類(lèi)生物;批號(hào):WL02117,規(guī)格:50 μL);cleaved caspase-9(CST;批號(hào):9509S,規(guī)格:100 μL);羊抗兔二抗(Abbkine;批號(hào):A23620,規(guī)格:50 μL);羊抗鼠二抗(Abbkine;批號(hào):A25012,規(guī)格:50 μL)
1.2 研究方法
1.2.1煙草煙霧暴露小鼠模型 將 36 只體質(zhì)量年齡大小相近的 C57 BL/6 小鼠按隨機(jī)分組的方法分為對(duì)照組、CS暴露組和CS+TSA 組,每組 12 只,對(duì)每組內(nèi)的小鼠進(jìn)行編號(hào),列表記錄它們的初始體質(zhì)量以及精神活動(dòng)狀態(tài)。自制實(shí)驗(yàn)性CS暴露裝置,CS暴露方法參照本課題組前期實(shí)驗(yàn)[6]。CS暴露組和CS+TSA 組小鼠CS暴露時(shí)間為 30 d,每天2次CS暴露,每次CS暴露給2支煙持續(xù)暴露 2 h,CS暴露時(shí)間間隔為 5 h。每次進(jìn)行CS暴露時(shí)取1支煙點(diǎn)燃,使煙霧進(jìn)入CS暴露組小鼠所處的箱子內(nèi),CS暴露組箱子放在無(wú)風(fēng)環(huán)境中,保持一定的空氣流通。對(duì)照組小鼠吸入空氣。藥物 TSA(將TSA溶于DMSO,再用生理鹽水將其配制成濃度為1.98×10-5mol·L-1的溶液),按 0.6 μg·g-1體質(zhì)量的劑量,每2 d 1次,腹腔內(nèi)注射給CS+TSA 組小鼠。CS暴露 30 d 后,對(duì)處于發(fā)情期的雌性小鼠實(shí)施安樂(lè)死,并從每組中至少收集了 5 個(gè)子宮用于進(jìn)一步研究。
1.2.2HE染色 CS暴露 30 d 后,取CS暴露組和對(duì)照組小鼠的子宮組織,用 4% 多聚甲醛固定 24 h,再經(jīng)梯度酒精的洗滌、脫水、二甲苯透明,浸蠟,進(jìn)行組織包埋,以4 μm 厚度連續(xù)切片,HE染色,中性樹(shù)脂封片后置于光學(xué)顯微鏡下觀察并拍照。
1.2.3Western blot 分析 提取對(duì)照組、CS暴露組和CS+TSA 組小鼠子宮組織總蛋白,然后將提取的各組子宮組織的總蛋白進(jìn)行 SDS-PAGE 電泳,用P2RX7(1 ∶ 2 000稀釋)、ASC(1 ∶ 2 000稀釋)、IL-1β(1 ∶ 2 000稀釋)、GAPDH(1 ∶ 5 000 稀釋)、cleaved caspase1(1 ∶ 2 000稀釋)、NLRP3(1 ∶ 500稀釋)、cleaved caspase3(1 ∶ 2 000稀釋)和 cleaved caspase-9(1 ∶ 2 000稀釋)抗體孵育。實(shí)驗(yàn)結(jié)束后用 Image-J 軟件對(duì)光密度值進(jìn)行定量檢測(cè),對(duì)不同分子量蛋白的表達(dá)情況進(jìn)行分析。
1.2.4Real time-PCR 使用 TRIzol 試劑從子宮中分離總 RNA(n=3~4/組),并用 oligo(dT)引物和 SuperScript Ⅱ 反轉(zhuǎn)錄 RNA(1 mg)。在具有 SYBR Green 的 Roche 480 PCR系統(tǒng)上進(jìn)行實(shí)時(shí)定量 PCR。將 GAPDH 作為內(nèi)源性對(duì)照,并使用 2-ΔΔct方法計(jì)算每個(gè)樣品的基因表達(dá)相對(duì)水平。基因的引物序列。

Tab 1 Primers for genes used in q-PCR
1.3 統(tǒng)計(jì)學(xué)分析方法應(yīng)用 GraphPad Prism 7.00 統(tǒng)計(jì)軟件分析數(shù)據(jù),2 組比較采用獨(dú)立樣本t檢驗(yàn),3 組比較采用單因素方差分析。
2 結(jié)果
2.1 CS暴露導(dǎo)致子宮組織結(jié)構(gòu)改變CS暴露導(dǎo)致子宮組織形態(tài)學(xué)改變。Fig 1C結(jié)果顯示,CS暴露后的小鼠比暴露于室內(nèi)空氣的小鼠的子宮肌層(中環(huán)肌和內(nèi)縱肌)和子宮內(nèi)膜更薄。Fig 1D結(jié)果顯示,CS暴露后小鼠子宮腺體組織和間質(zhì)細(xì)胞明顯減少。我們先前的研究結(jié)果發(fā)現(xiàn),TSA 有效抑制了CS暴露引起的小鼠子宮肌層變薄以及腺體組織和間質(zhì)細(xì)胞的減少[6]。
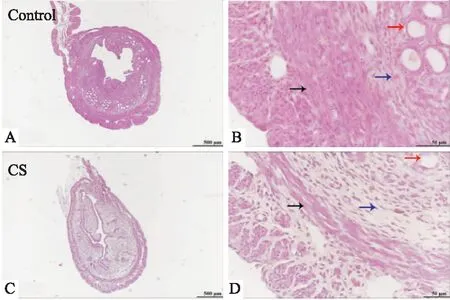
Fig 1 Histomorphologic alterations in uterine caused by CS exposure
A: The morphology of uterine tissues of mice in the optical microscope (HE,×40) Control group; B: The enlarged optical microscope image of the myometrial region in Fig A (HE,×400); C: The morphology of uterine tissues of mice in the optical microscope (HE,×40) CS group; D: The enlarged optical microscope image of the myometrial region in Fig C (HE,×400). Blue arrows point to interstitial cells, black arrows to myometrium and red arrows to glandular tissues.
2.2 TSA 抑制了CS暴露誘導(dǎo)的小鼠子宮組織細(xì)胞焦亡的發(fā)生我們前期的研究表明,CS暴露能夠引起小鼠子宮組織形態(tài)學(xué)改變,TSA 能夠改善CS暴露引起的小鼠子宮組織形態(tài)學(xué)改變[6]。因此,我們進(jìn)一步探索了TSA是否能夠通過(guò)抑制P2RX7 介導(dǎo)的細(xì)胞焦亡緩解CS暴露引起的子宮組織形態(tài)學(xué)改變。首先,我們采用 Western blot 技術(shù)檢測(cè)了各組(對(duì)照組、CS暴露組和CS+TSA 組)小鼠子宮組織中P2RX7 和細(xì)胞焦亡相關(guān)蛋白的表達(dá)水平,細(xì)胞焦亡相關(guān)蛋白包括核苷酸結(jié)合寡聚化結(jié)構(gòu)域樣受體3(NACHT-LRRPYD-containing proteins 3 inflammasome,NLRP3),白細(xì)胞介素 1β (interleukin 1β,IL-1β),凋亡相關(guān)斑點(diǎn)樣蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)和cleaved caspase 1。結(jié)果表明,CS暴露后小鼠子宮組織P2RX7 和細(xì)胞焦亡相關(guān)蛋白表達(dá)水平明顯升高,TSA有效抑制了CS暴露誘導(dǎo)的小鼠子宮組織P2RX7和細(xì)胞焦亡相關(guān)蛋白的表達(dá)。此外,我們通過(guò) Real time-PCR 檢測(cè)了小鼠子宮組織中Hdac1,IL-1β,P2rx7 和Asc的mRNA轉(zhuǎn)錄水平。結(jié)果表明,CS暴露后小鼠子宮組織中Hdac1,P2rx7和Asc的mRNA轉(zhuǎn)錄活性升高,并且TSA明顯抑制了CS暴露后小鼠子宮組織中Hdac1,Il-1β,P2rx7和Asc的mRNA 轉(zhuǎn)錄活性。因此,我們的結(jié)果表明,TSA能夠通過(guò)抑制P2RX7 介導(dǎo)的細(xì)胞焦亡改善CS暴露引起的子宮組織形態(tài)改變。
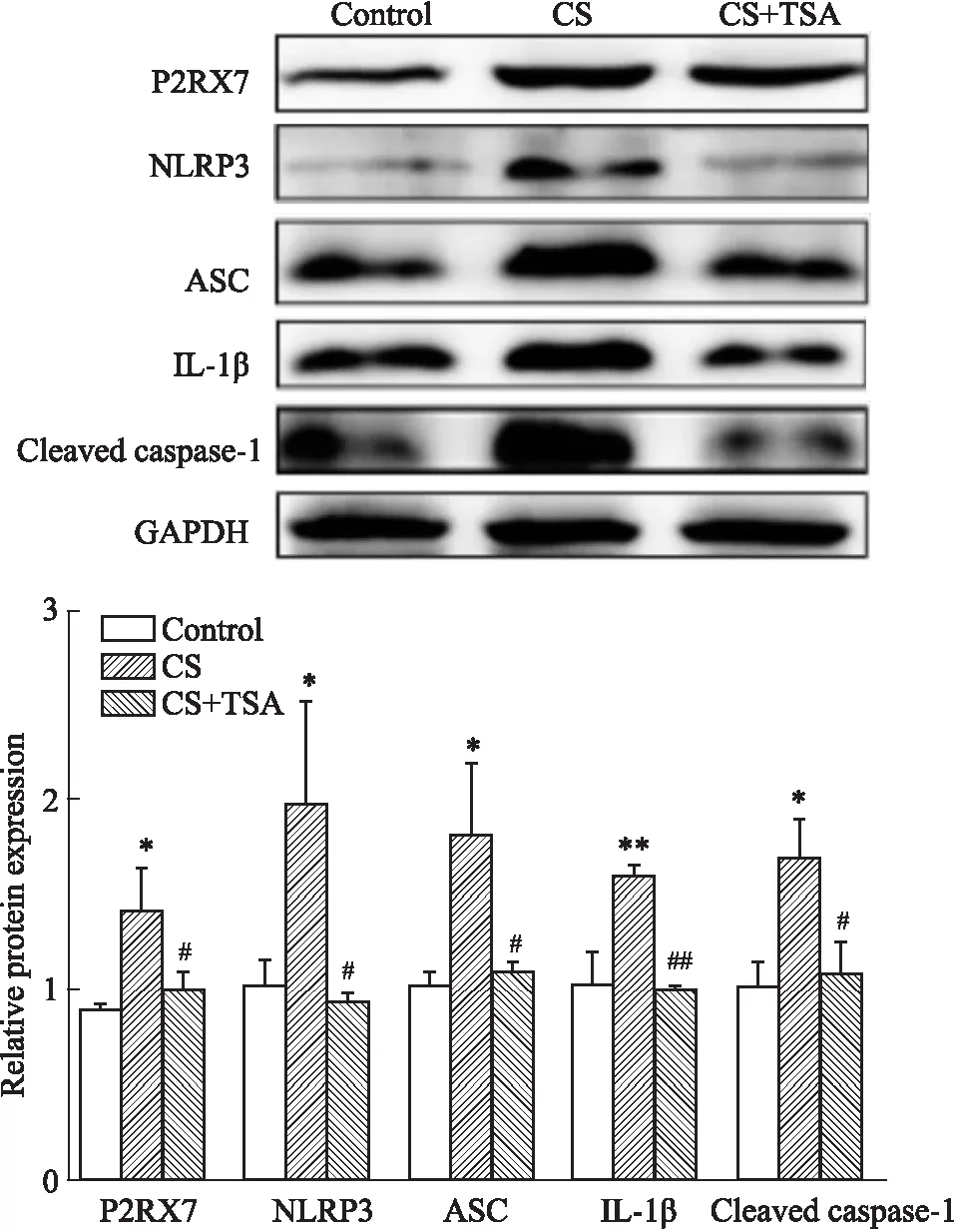
Fig 2 Activation of markers of pyroptosis activation induced by CS exposure inhibited by TSA )
Blots of expression of P2RX7 and pyroptosis related proteins in three groups.*P<0.05,**P<0.01vscontrol;#P<0.05,##P<0.01vsCS
2.3 TSA 抑制了CS暴露誘導(dǎo)的小鼠子宮組織細(xì)胞凋亡的發(fā)生為了進(jìn)一步探索TSA能否通過(guò)抑制P2RX7 介導(dǎo)的細(xì)胞凋亡,緩解CS暴露導(dǎo)致的小鼠子宮損傷,我們使用 Western blot 技術(shù)檢測(cè)了各組(對(duì)照組、CS暴露組和CS+TSA 組)小鼠子宮組織中細(xì)胞凋亡相關(guān)蛋白(cleaved caspase-3 和cleaved caspase-9)的表達(dá)水平。結(jié)果表明,CS暴露后小鼠子宮組織細(xì)胞凋亡相關(guān)蛋白的表達(dá)水平增加,TSA 抑制了CS暴露誘導(dǎo)的小鼠子宮組織細(xì)胞凋亡相關(guān)蛋白的表達(dá)。因此,我們的研究結(jié)果證實(shí)TSA也能夠通過(guò)抑制P2RX7 介導(dǎo)的細(xì)胞凋亡改善CS暴露引起的子宮組織形態(tài)改變。
Electrophoresis results of PCR products ofGapdh,Hdac1,P2rx7,AscandIl-1β. The mRNA levels ofHdac1,P2rx7 andAscincreased andIl-1βdecreased in the uterus after CS exposure, and TSA significantly inhibited the transcriptional activity ofHdac1,P2rx7 andAsc. Data are shown as means±SEM.**P<0.01vscontrol;#P<0.05vsCS
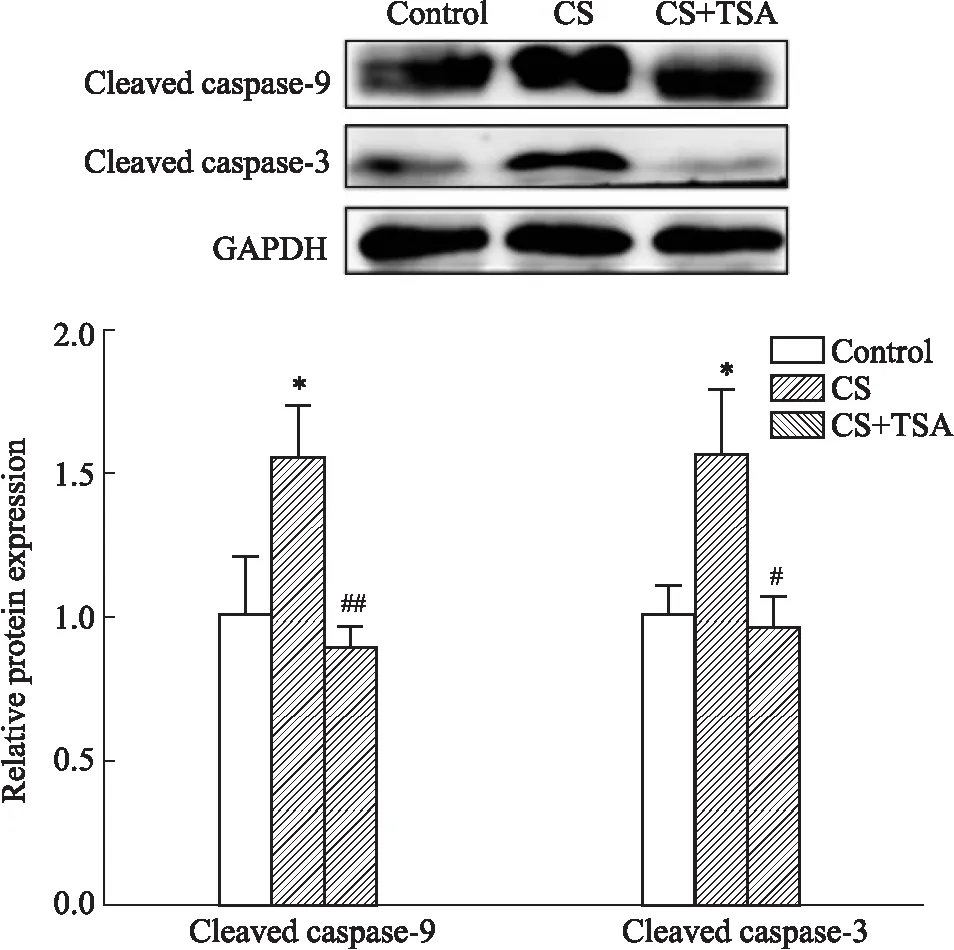
Fig 4 Activation of markers of apoptosis activation induced by CS exposure inhibited by TSA n=3 )
Blots of expression of apoptosis related proteins in three groups.*P<0.05vscontrol;#P<0.05;##P<0.01vsCS
3 討論
CS混合物中含有碳?xì)漕?lèi)化合物,醇類(lèi),酚類(lèi),醛類(lèi),酮類(lèi),生物堿類(lèi),酸類(lèi)和重金屬等多種有害的化學(xué)物質(zhì),這些有害物質(zhì)能夠?qū)е屡陨彻δ苁軗p。有研究報(bào)道,CS暴露可通過(guò) AMP 激活的蛋白激酶(adenosine 5’-monophosphate-activated protein kinase,AMPK)途徑誘導(dǎo)自噬級(jí)聯(lián)反應(yīng),以及抑制抗自噬相關(guān)蛋白 AKT 和 mTOR 的表達(dá),導(dǎo)致卵巢卵泡丟失[11],同時(shí),臨床研究表明,CS暴露后可能通過(guò)多種途徑導(dǎo)致卵巢雌激素合成減少[12],因此,CS暴露能夠損傷女性卵巢功能。雖然關(guān)于CS暴露導(dǎo)致卵巢損傷使女性生育能力受損的報(bào)道很多,但是關(guān)于CS暴露導(dǎo)致子宮損傷的作用和機(jī)制的研究較少。子宮在女性生育過(guò)程中發(fā)揮重要的作用,如果子宮組織發(fā)生結(jié)構(gòu)改變,可影響受精卵的著床,胚胎發(fā)育和激素分泌等多個(gè)過(guò)程,進(jìn)而使女性生育能力受損。僅有的文獻(xiàn)證實(shí),口服尼古丁可以導(dǎo)致大鼠子宮組織重量減少、使上皮組織、子宮肌層和子宮內(nèi)膜變薄[4]。本實(shí)驗(yàn)使用CS暴露 30 d 的方法建立了小鼠子宮損傷模型,研究發(fā)現(xiàn),CS暴露導(dǎo)致 C57 BL/6 小鼠子宮組織結(jié)構(gòu)形態(tài)學(xué)改變,包括子宮肌層和子宮內(nèi)膜變薄,子宮腺體組織和間質(zhì)細(xì)胞也明顯減少。本研究CS暴露 30 d 后 C57 BL/6 小鼠子宮組織發(fā)生的結(jié)構(gòu)變化和文獻(xiàn)報(bào)道的CS暴露導(dǎo)致的大鼠子宮組織結(jié)構(gòu)改變相似。TSA 是一種泛去乙酰化酶抑制劑,可以通過(guò)改變蛋白質(zhì)的乙酰化水平調(diào)節(jié)蛋白質(zhì)的功能。關(guān)于TSA的治療作用已經(jīng)有多篇文獻(xiàn)進(jìn)行了報(bào)道,例如TSA可以改善大鼠腎上腺嗜鉻細(xì)胞瘤導(dǎo)致的 PC12 細(xì)胞缺血性損傷[5]。我們的前期研究結(jié)果發(fā)現(xiàn)CS暴露能夠通過(guò)激活 HDAC1/2 導(dǎo)致子宮肌層和子宮內(nèi)膜變薄,而TSA可以抑制 HDAC1/2 和 mTOR 的活化重新激活自噬,從而緩解CS暴露引起的子宮組織形態(tài)改變[6]。因此,我們的CS暴露方法可以成功制作出子宮損傷的小鼠動(dòng)物模型,為進(jìn)一步揭示CS暴露導(dǎo)致子宮損傷的機(jī)制提供了很好的前期基礎(chǔ)。由于CS含有多種復(fù)雜的有害成分,所以,關(guān)于CS暴露導(dǎo)致的子宮損傷的機(jī)制還需要我們進(jìn)一步探索,為臨床治療吸煙引起的女性不孕提供新的靶標(biāo)。
在本研究中我們發(fā)現(xiàn)CS暴露導(dǎo)致小鼠子宮組織中P2RX7、細(xì)胞焦亡相關(guān)蛋白(ASC、NLRP3、IL-1β和cleaved caspase-1)和細(xì)胞凋亡相關(guān)蛋白(cleaved caspase-3和cleaved caspase-9)的表達(dá)水平升高。因此,CS暴露可能通過(guò)P2RX7 介導(dǎo)的細(xì)胞焦亡和凋亡導(dǎo)致子宮組織形態(tài)學(xué)改變。細(xì)胞焦亡是一種主要由炎性小體介導(dǎo)的程序性細(xì)胞死亡方式,通過(guò)切割 GSDMD 蛋白在細(xì)胞膜上打孔,改變細(xì)胞滲透壓使細(xì)胞膨脹,釋放內(nèi)容物和相關(guān)炎癥因子導(dǎo)致機(jī)體炎癥反應(yīng)的發(fā)生,在機(jī)體免疫調(diào)節(jié)中發(fā)揮重要作用。細(xì)胞凋亡也是一種程序性細(xì)胞死亡方式,它的發(fā)生主要依賴(lài)于半胱天冬酶 caspase-3、6、8 等,伴隨著凋亡小體的形成,由細(xì)胞膜對(duì)細(xì)胞內(nèi)容物進(jìn)行包裹,再由巨噬細(xì)胞對(duì)其進(jìn)行非炎癥性吞噬,進(jìn)而維持機(jī)體內(nèi)環(huán)境的穩(wěn)定[13]。研究發(fā)現(xiàn),細(xì)胞焦亡和凋亡在子宮的某些病理?yè)p傷變化中發(fā)揮著重要作用。例如金黃色葡萄球菌通過(guò)誘導(dǎo)子宮組織發(fā)生細(xì)胞焦亡和凋亡引起子宮內(nèi)膜炎[14],順鉑在雌性大鼠卵巢和子宮組織中能夠參與氧化應(yīng)激、炎癥反應(yīng)和細(xì)胞凋亡的發(fā)生等[15]。以上發(fā)現(xiàn)均表明細(xì)胞焦亡和凋亡在子宮組織的發(fā)育過(guò)程中發(fā)揮了重要作用,因此,抑制炎癥反應(yīng)和細(xì)胞凋亡都有可能治療子宮的病理?yè)p傷變化。P2RX7受體是同源三聚體配體門(mén)控陽(yáng)離子通道,其特征在于有2個(gè)跨膜結(jié)構(gòu)域,這種受體已在全身多種細(xì)胞中發(fā)現(xiàn),但是在免疫細(xì)胞中表達(dá)最多。ATP是P2RX7受體的天然配體,胞外 ATP 激活受體會(huì)引起 Ca2+、Na+和K+在質(zhì)膜上的流動(dòng)。P2RX7受體可以引起Ca2+的強(qiáng)烈增加和 caspase 的裂解來(lái)啟動(dòng)細(xì)胞凋亡,還可通過(guò)激活 caspase-1 和 NLRP3 來(lái)啟動(dòng)細(xì)胞焦亡。P2RX7 參與了很多疾病的發(fā)展過(guò)程,研究發(fā)現(xiàn)P2RX7 抑制劑具有治療多種疾病的潛力,包括神經(jīng)退行性疾病、精神疾病、癲癇和肌肉骨骼疾病等[16]。關(guān)于CS暴露后P2RX7介導(dǎo)的細(xì)胞焦亡導(dǎo)致慢性阻塞性肺病肺組織損傷的報(bào)道很多[9]。但是P2RX7 受體在子宮中相關(guān)功能的研究很少,僅有的文獻(xiàn)證實(shí),P2RX7受體能夠介導(dǎo)人宮頸上皮細(xì)胞凋亡[10]。迄今為止,CS暴露后P2RX7 受體在子宮組織中的作用尚未有人進(jìn)行研究。關(guān)于P2RX7 介導(dǎo)的細(xì)胞焦亡和凋亡在CS暴露所致的子宮損傷中是否發(fā)揮作用尚不清楚。本研究證實(shí)了P2RX7 介導(dǎo)的細(xì)胞焦亡和凋亡參與了CS暴露導(dǎo)致的雌性小鼠子宮損傷。然而,P2RX7 受體是否參與TSA對(duì)CS暴露導(dǎo)致的子宮組織損傷的緩解作用還未有人進(jìn)行研究。本研究發(fā)現(xiàn),TSA抑制了CS暴露導(dǎo)致的P2RX7、細(xì)胞焦亡相關(guān)蛋白(ASC、NLRP3、IL-1β和cleaved caspase-1)和細(xì)胞凋亡相關(guān)蛋白(cleaved caspase-3和cleaved caspase-9)的表達(dá)水平升高,并且TSA抑制了CS暴露導(dǎo)致的子宮組織Hdac1、P2rx7、AscmRNA 轉(zhuǎn)錄水平升高。因此,本研究證實(shí)P2RX7 介導(dǎo)的細(xì)胞焦亡和凋亡參與了TSA緩解的CS暴露所致的小鼠子宮損傷過(guò)程。CS暴露后子宮組織Il-1β的mRNA 轉(zhuǎn)錄水平與蛋白表達(dá)水平不一致,我們推測(cè)可能存在其它復(fù)雜的轉(zhuǎn)錄調(diào)控機(jī)制影響Il-1β的mRNA轉(zhuǎn)錄水平,未來(lái)我們會(huì)進(jìn)一步進(jìn)行探索。
綜上所述,P2RX7 介導(dǎo)的細(xì)胞焦亡和凋亡參與TSA緩解的煙草煙霧暴露所致的小鼠子宮損傷過(guò)程。本研究為T(mén)SA靶向治療煙草煙霧暴露導(dǎo)致的女性生育能力受損提供了依據(jù),為闡明煙草煙霧暴露導(dǎo)致女性不孕的機(jī)制奠定了基礎(chǔ),對(duì)未來(lái)治療吸煙引起的女性不孕提供一定的指導(dǎo)意義。
(致謝:感謝中國(guó)醫(yī)科大學(xué)附屬盛京醫(yī)院實(shí)驗(yàn)研究中心提供實(shí)驗(yàn)平臺(tái))。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
汽車(chē)工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
